Тыныс алу жүйелері
Тыныс алу қызметі.
Тыныс алу жүйесінің құрылысы мен қызметі адамның жасына, жынысына байланысты ерекшеліктері болады.
Тыныс алу жүйесінің мүшелері қызметтеріне қарай екіге бөлінеді:
ауаны сыртқы ортадан ішке және іштен сыртқа қарай қозғайтын тыныс алу жолдары - мұрын қуысы, көмей, кеңірдек және бронхылар.
түскен газдарды алмастыру процессін жүргізетін мүше - өкпе;
Бұл мүшелер орналасулары бойынша жоғарғы және төменгі тыныс алу жолдарына болып бөлінеді:
жоғарғы тыныс алу жолдары - мұрын қуысы, жұтқыншақ, ауыз қуысы;
төменгі тыныс алу жолдары - көмей, кеңірдек, бронхы.
Мұрын қуысы қызметі
Ауа мұрын арқылы екі бөлікті-қалқалы мұрын қуысына түседі. Әрбір бөлікті-қалқада мұрын қуысының ішкі бетін ұлғайтатын үш мұрын қалқаны орналасқан. Ол эпителийлі қабықпен және көп қан тамырларымен жабылған. Тамырлармен аққан қан түскен ауаны дене температурасына дейін жылытады, ал сілемейлі қабық ауаны ылғалдайды және шаң-тозаңдар мен микроорганизмдерді тұтып қалады. Тұтылған микроорганизмдерді лейкоциттер қорытады (фагоцитоз), қорытудан артылған шаң-тозаңдар эпителий түктері (кірпікшелер) арқылы сыртқа шығарылады. Мұрын қуысында жылытылған, ылғалданған және тазартылған ауа жұтқыншақ арқылы көмейге түседі.
Көмей қызметі
Көмей - іші қуыс түтік, қабырғалары сіңір, буын және бұлшық еттермен қосылған бірнеше шеміршек тұрады. Көмейге ас бөлшектері немесе басқа заттар түскенде, сондай-ақ қабыну процестерінде адам қатты жөтелгенде дем шығарылады. Бұл көмейдің тазартылуына әсер етіп, тыныстың төменгі бөлімдеріне зиянды заттардың енуіне кедергі келтіреді. Көмей ауаны жұтқыншақ арқылы кеңірдекке өткізеді.
Кеңірдек және бронхылар қызметі
Кеңірдек - өңештің алдында орналасқан ұзындығы 9-13 см, диаметрі 15 мм. түтік. Ол кеңірдек қабырғасының қабысып қалуына кедергі келтіретін шеміршекті жартылай сақиналардан тұрады. Сақиналар мойынның қандай қозғалысы болмасын ауаны ұстап қалмайды Өңешке кеңірдектің артқы жұмсақ қабырғасы жанасып, астың өңешпен еркін жылжуына мүмкіндік береді. V кеуде омыртқасы түсында кеңірдек екі бронхыға бөлінеді. Бронхылар өкпенің оң жақ және сол жақ бөліктеріне кіріп, тармақталып, бронхы ағашын түзеді. Ұштарындағы жіңішке тармақтары - бронхиолалар өкпе көпіршіктері - альвеолалармен аяқталады.
Өкпенің қызметі
Кіші қан айналымы арқылы өкпеге веналық (көк) қан түседі, бұл жерде оттегімен қанығып, көмір қышқыл газынан ажыратылып, алқызыл түсті артериялық қанға айналады. Көмір қышқыл газы өкпе көпіршіктеріне (альвеолаға) өтіп, дем шығару кезінде ағзадан шығады. Артериялық қан ары қарай үлкен қан айналымы арқылы дене мүшелеріне өтіп, олардың жасушаларын ауамен қамтиды, оны тұтыну барысында жасушалардан көмір қышқыл газы бөлініп шығып, қан қайтадан веналық (көк) болады.
Тыныс алу бөліміндегі газ алмасу механизмі.
Газ алмасу, биологияда – ағза мен оны қоршаған орта арасындағы газ алмасу процесі. Адам мен жануарлар ағзасы тыныс алу кезінде сыртқы ортадан оттекті сіңіріп, өзінен көмір қышқыл газын және зат алмасу нәтижесінде пайда болатын әр түрлі газ қоспаларын сыртқа шығарады. Газ алмасусыз тірі ағзада зат және энергия алмасуы дұрыс жүрмейді. Зат алмасу процесінің нәтижесінде ағза сіңірген қоректік заттардың (белок, көмірсу, май) құрамындағы химиялық энергия бөлініп, ағзаның тіршілік әрекетіне қажетті энергияға (мысалы, 1 л оттек 4,7 – 5,0 ккал жылу береді) айналады. Организм пайдаланатын оттек пен одан бөлініп шығатын көмір қышқыл газдың мөлшері адамның жасына, тіршілік әрекетіне, атқаратын қызметіне, сыртқы ортаның температурасына және тағамның құрамына байланысты өзгеріп тұрады. Адам жұмыс істегенде, ауа райы салқындағанда, калориялы тамақ ішкенде ағзадағы газ алмасу күшейе түседі. Демалыс кезінде, ұйықтағанда газ алмасу төмендейді. Газ алмасу арқылы ағзадағы энергия мөлшерін анықтауға болады. Газ алмасу процесі кезінде ағза оттекті көп мөлшерде пайдаланса, барлық дене мүшелерінің қызметі артып, тыныс алу және қан айналысы жақсарады, осыған байланысты қанның физикалық-химиялық құрамы да өзгереді. Газ алмасу бір клеткалы қарапайымдыларда, олардың бүтіндей денесі арқылы, көп клеткалы жануарларда арнаулы тыныс мүшелері (кеңірдек, желбезек, өкпе) арқылы жүреді. Өсімдіктердегі газ алмасу жануарлар мен адам ағзасындағыдан өзгеше болады. Тірі азғалар сияқты өсімдіктер де барлық мүшелері арқылы тыныс алады, тыныс алу процесі өсімдіктің өсіп келе жатқан жас мүшелері мен тіндерінде, әсіресе, жапырақтарында күшті, ал сабағы мен тамырында баяу жүреді. Өсімдіктер тыныс алған кезде, керісінше, ауадан көмір қышқыл газ сіңіріліп, оттек бөлінеді.
Өкпедегі газ алмасу – тыныс алғанда атмосфералық ауаның құрамында 79% – азот, 21% – оттек, 0,03% – көмірқышқыл газы және т. б. болады. Тыныс шығарғанда
оттектің мөлшері азайып (16%), көмірқышқыл газы (4%) көбейеді (69, асурет). Тыныс алу мен тыныс шығару кезінде азоттың мөлшері
өзгермейді. Тыныс алу актысы кезінде өкпе көпіршіктерінен оттек
қанға, көмірқышқыл газы өкпеге өтеді.
Оттекті тасушы – гемоглобин. Өкпеде қан көмірқышқыл газынан
тазарып, оттекке қанығады. Оттекке қаныққан қан үлкен қанайналым
шеңберімен ағзаның барлық ұлпаларына таралады.
Ұлпалардағы газ алмасу. Ұлпа жасушаларындағы оттектен
артерия қанындағы оттектің мөлшері көп болады. Оттек
капиллярлардың қабырғаларынан жасушаларға өтеді де, олардың
тіршілік әрекеттеріне жұмсалады. Артерия қанымен жасушалардың газ
алмасуы диффузия жолымен жүреді. Түзілген көмірқышқыл газы ұлпа
жасушаларынан қанға өтіп, артерия қаны вена қанына айналады.
Ұлпаларда қан оттекті беріп, көмірқышқыл газына қанығады.
Өкпедегі және ұлпалардағы газ алмасу үш кезеңнен тұрады.
Біріншісі – сыртқы (өкпелік), екіншісі – газдардың қан арқылы
тасымалдануы, үшіншісі – ішкі (ұлпалық) газ алмасу деп аталады.
Альвеолалардағы сұйықтықтың тасымалдануын реттеу
Сурфактант функциялары
Сурфактант - су-ауа шекарасында орналасқан липопротеидті беттік белсенді зат, альвеолаларды ішкі жағынан қаптайды және беттік керілуді азайту арқылы дем шығару кезінде альвеолалардың құлауын болдырмайды. Сурфактантты өкпе жасушаларының ерекше түрі – II типті альвеолоциттер бөледі. Туа біткен беттік-активті заттардың жетіспеушілігі жаңа туған нәрестелерде өкпенің ашылмауымен көрінеді - респираторлық дистресс синдромы (РДС).
Беттік белсенді заттың негізгі компоненттері фосфолипидтер, ақуыздар және ішінара полисахаридтер.
Сурфактант функциялары
• Өкпенің серпімділігінің жоғарылауы;
• Ателектаздың алдын алу (дем шығару кезінде өкпенің құлауы);
• Иммундық қорғанысқа қатысу.
Альвеолалар сәнде судағы ауа көпіршіктері ретінде қарастырылады. Су-ауа шекарасындағы беттік керілу күштері альвеолаларды қысуға бейім. Сондықтан олардың құлауын болдырмау үшін Р альвеолаларында белгілі бір ауа қысымы қажет, оны Лаплас теңдеуі бойынша есептеуге болады, ол Р-ның беттік керілуге (γ) және альвеолалардың радиусына (r) тәуелділігін көрсетеді:
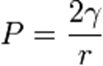
Тыныс алу жолдарының жасушалық құрамы.
Мұрын қуысының тыныс алу бөлігі бір қабатты, көп қатарлы кірпікшелі
эпителимен тысталған, құрамында кірпікшелі, бокал тəрізді, жиекті немесе
микробүрлері бар қыстырма немесе базальді жасушалардан тұрады.
Аталған жасушалардың көмегі арқылы мұрын қуысына ауадан келіп түскен
бөтен- текті заттар мен микроорганизмдер сыртқа шығады не болмаса
жойылады. Бокал тəрізді жасушалары- муцинді бөледі. Муцин бөтен
заттарды өзіне жабыстырады да, сыртқа шығуына септігін тигізеді. Мұрын
қуысынан бөлінетін секреттің құрамында антидене-иммуноглобулин «А»
эпителидің астындағы меншікті пластинкасының құрамындағы
плазмоциттерден бөлінеді. Бұл плазмоциттер лизоцимді де синтездейді.
Жиекті жасушалар-хеморецепторлар болып саналады. Базальді немесе
қыстырма жасушалары регенерациялық- камбиалді қызмет атқарады.
Эпителидің меншікті пластинкасының құрамындағы борпылдақ дəнекер
тінінде белокты-шырышты, аралас бездермен бірге лимфоидты
фолликулдар да болады.
Кеңірдек эпителиінің құрамында бірнеше жасушалар қатары болады. Оларға: кірпікшелі, бокал тəрізді жасушасы, қыстырма немесе базальді жəне эндокриноциттер жатады. Бокал тəрізді жасуша мен кірпікшелі жасушалардың құрылысы мен қызметі мұрын қуысындағы жасушаларға ұқсас. Шырышты-кірпікшелі жасушалары мукоцилиярлы конвейер түзеді. Қалыпты жағдайда бокал тəрізді мен кірпікшелі жасушалардың саны бір-біріне тең. Ал, патологиялық кезде мысалы: шылым тартатын адамдардың кеңірдегінің қабыну процестерінде бокал тəрізді жасушаларының саны көбейеді де, секрет көп бөлініп, қатайып, бактериоцидтік қызметі нашарлайды да «темекішіліктің трахит» науқасы байқалады. Бұл науқас бронхтарда да болуы мүмкін («бронхит»). Кеңірдек эпителиіндегі эндокриноциттердің пішіні-призма тəрізді. Ерекшеліктері: базальді бөлігінде гормоналді түйіршіктері болады. Мұндай гормоналді түйіршіктерімен:серотонин, кальцитонин, бомбезин, нейромедиаторлар, норадреналин мен пептидтер бөлінеді. Бұл гормондардың қызметі - ауа жолындағы бездердің секрециясын жəне ет тіндерінің жиырылғыштық қасиетін реттеу. Базалді жасушалары- камбиалді қызмет атқарады. Соңғы мəліметтер бойынша кеңірдек пен ауа жолдарының эпителиінде Лангерганс жасушаларының бар екені анықталған.
Өкпенің жасушалары: Секреторлы жасушалары (Клара жасушасы) микробүрлері жоқ,
цитоплазмасында көптеген түйіршіктері мен белок синтезіне қатысатын
органеллалары болады. Бұлардың құрамында ферменттері де болады жəне
сурфактант бұзатын секрет бөледі.
Кірпікшесіз жасушалардың цитоплазмасында гликоген мен секреторлы
түйіршіктері бар екені анықталған, бірақ қызметі əлі белгісіз.
Жиекті жасушалардың микробүрлері мен нейросекреторлы түйіршіктері
болады. Бұлар қанға кальцитонин, бомбезин, серотонин гормондарын бөліп,
хеморецепциялық қызмет атқарады.
Тыныс алу жолдарын бактерияға қарсы қорғаудың спецификалық емес факторлары.
Өздеріңіз білетіндей, тыныс алу жолдарында жергілікті иммунитетті қоса алғанда, инфекцияға қарсы қорғаныс механизмдері бар. Соңғы жылдары тыныс алу жолдарының иммунологиялық белсенділігі туралы көбірек айтылады, бұл жергілікті иммунитеттің күйімен байланысты. Жергілікті иммунитет – келесі антигендік тітіркенуге бірінші болып әрекет ететін тыныс алу жолдары мен өңеш жолдарының қиылысында орналасқан жұтқыншақтың шырышты қабығының және лимфоэпителиальды мүшелерінің бейспецификалық қорғаныс факторларын қамтитын шырышты қабықтың тосқауылдық қызметі. иммундық қорғаныс механизмдерін белсендіру.
Бастапқыда жергілікті иммунитет деп шырышты жасушалардың тосқауылдық қызметтерін, нейтрофилдер мен макрофагтардың фагоцитарлық белсенділігін, Т-клетка иммунитетін, антиденелерді, сыртқы секрецияның микробқа қарсы ақуыздарын, фермент ингибиторларын қамтитын жасушалық және секреторлық спецификалық емес және спецификалық реакциялар кешені түсіндірілді. Жергілікті иммунитет секреторлық иммунитетпен анықталмайды, бірақ оның орталық буыны ретінде секреторлық компонентті қамтамасыз ететін безді эпителийдің қатысуымен шырышты қабаттардың лимфоидты ұлпасының В-жасушалық реакциясы қарастырылады. Кейінірек жергілікті иммунитет түсінігі айтарлықтай кеңейді және қазір макрофагтармен, нейтрофилдермен, эозинофильді гранулоциттермен, мастикалық жасушалармен және дәнекер тінінің және эпителийдің басқа жасушаларымен ынтымақтаса отырып, шырышты қабаттарды толтыратын лимфоидты қатардағы барлық жасушалардың жауаптарының жиынтығын қамтиды. . Шырышты қабықтардың тосқауыл құрайтын және макроорганизмді әртүрлі патогенді және шартты-патогенді микрофлоралардың патогендік әсерінен қорғайтын ерекше қорғаныс механизмдері иммунокомпетентті және көмекші жасушаларды, әртүрлі кластағы иммуноглобулиндерді, соның ішінде секреторлық, секреторлық, микрофлораны қоса алғанда, шырышты қабықтың иммундық жүйесі деп аталады. цитокиндер.
Жоғарғы тыныс алу жолдары - мұрын, мұрын қуысы, жұтқыншақ - тыныс алатын ауаны онда ілінген бөлшектердің едәуір бөлігінен тазартатын өзін-өзі тазартатын аэрозольдік сүзгі.
Балалардағы тар алдыңғы мұрын саңылаулары арқылы ауа мұрын қалқасынан және конкасынан түзілген кең кеңістікке енеді. Мұнда ауа ағыны ламинарлы сипатын жоғалтады. Шырышты қабаттың бетіндегі бөлшектердің тұнбасына әкелетін құйынды турбулентті қозғалыстар пайда болады. Ауырлық күші үлкенірек бөлшектерді тұндыру үшін де маңызды. Нәтижесінде тыныс алу жолдары 50 мкм өлшемдегі барлық бөлшектерді сақтайды. Тыныс алу жүйесіндегі әртүрлі мөлшердегі бөлшектер былай бөлінеді: 50 мкм-ден асатын бөлшектер мұрында, мұрын қуысында және мұрын-жұтқыншақта ұсталады, диаметрі 30-50 мкм бөлшектер трахеяға, 10-30 мкм бронхқа өтеді. , 3-10 мкм - бронхиолаларға , 1-3 мкм - альвеолаларға.
Хлор арнасындағы муковисцидозға арналған трансмембраналық өткізгіштікті реттегіш
Муковисцидоздың трансмембраналық реттегіші (CFTR - Cystic Fibroz Transmembrane keçiricilik реттегіші) - жасуша мембранасы арқылы хлор иондарын тасымалдауға қатысатын ақуыз, сондай-ақ осы ақуызды кодтайтын геннің атауы. CFTR гені 7-хромосоманың ұзын қолында орналасқан. CFTR генінің мутациялары муковисцидоз ауруының басталуына әкеледі, сонымен қатар ер бедеулігінің себебі болуы мүмкін. Ең жиі кездесетін ΔF508 мутациясы (барлық анықталған гендік мутациялардың 50%-дан астамы), онда фенилаланин-508 қалдығы полипептидтік тізбектен жойылады, бұл плазмалық мембранадағы ақуыздың қатпарлануының бұзылуына әкеледі.
CFTR ақуызы негізінен тыныс алу жолдарының, сілекейдің, тер бездерінің, ұйқы безінің және ішектің эпителий жасушаларында локализацияланған. Жасуша ішінде ол апикальды мембрананың бетінде, сонымен қатар эндоплазмалық тордың мембраналарында және пиноцитарлы көпіршіктердің бөлігі ретінде орналасады. CFTR - хлор иондарын белсенді тасымалдауға арналған мембраналық арна.
Респираторлық вирустардың өмірлік циклі.
Жедел респираторлық инфекциялар (ЖРВИ) – көптеген қоздырғыштар тудыратын, ауа тамшылары арқылы берілетін және адамның тыныс алу жүйесінің жедел зақымдалуымен сипатталатын аурулар тобы. ЖРВИ-ға тұмау, парагрипп, аденовирустық, респираторлық синцитиальды, риновирустық инфекциялар және т.б.
Тұмау – тұмау вирусы тудыратын, ең көп асқынатын жіті респираторлық инфекциялардың бірі.
Тұмаудың қоздырғыштары тұмау вирусының үш түрі (А типі, В типі және С типі). Тұмау вирусының барлық үш түрі парамиксовирустар тобына жатады, алайда олар құрылымы жағынан айтарлықтай ерекшеленеді, сондықтан вирустың бір түріне қарсы иммунитет басқа түрдегі вирусқа қарсы «жұмыс істемейді». Сонымен қатар, бір типті (әсіресе А типті) вирустар құрылымын тез өзгертуге және өзгертуге қабілетті. Осы процестің нәтижесінде жыл сайын біздің иммундық жүйемізге белгісіз тұмау вирустарының жаңа түрлері шығарылады. Адамдардың барлық жас санаттары тұмауға бейім.
Күзгі-көктемгі және қысқы айларда максималды таралу гипотермиямен байланысты, бұл осы аурулардың дамуына мүмкіндігінше ықпал етеді. Тәуекел тобына балалар, қарт адамдар, жүкті әйелдер, созылмалы жүрек және өкпе аурулары бар адамдар жатады.
Инфекцияның көзі – клиникалық айқын немесе жойылған түрімен ауыратын адам. Инфекция ауа тамшылары арқылы және ластанған қолдар немесе респираторлық вирустарды тасымалдайтын заттар арқылы өтеді.
ЖРВИ-ның негізгі белгілері - мұрынның ағуы, жөтел, түшкіру, бас ауруы, тамақ ауруы, шаршау, дене қызуының көтерілуі.
Тұмау клиникасы. Тұмаудың инкубациялық кезеңі өте қысқа – бірнеше сағаттан 2-3 күнге дейін.
Тұмау вирусының барлық түрлері тыныс алу жолдарының шырышты қабығына жоғары жақындыққа ие, онда вирус бірінші кезекте көбейеді. Тұмау белгілері тез өсуде: тамақ ауруы, түшкіру, дене температурасы қатты көтеріледі (40 ° C дейін), әлсіздік, қатты бұлшықет және бас аурулары пайда болады. Келесі күндері тұрақты температураның фонында төс сүйегінің артындағы ауырсынумен бірге жүретін құрғақ, шулы жөтел пайда болады. Құрғақ жөтел бірте-бірте ылғалды, кеудеге айналады. Аурудың асқынбаған ағымымен тұмау белгілерінің регрессиясы 5-6-шы күні байқалады.
Тыныс алу жүйесі тарапынан бронхит пен пневмонияның дамуы мүмкін (өкпе тіндерінде вирустың көбеюінен туындаған ауыр геморрагиялық пневмония сирек дамиды). Тұмаумен ауыратын балаларда вирустық энцефалит немесе менингит дамуы мүмкін.
Тыныс алу ацидозы
Өкпенің вентиляциясының бұзылуы
өкпенің зақымдануы немесе ауруы (пневмония, фиброз, өкпе ісінуі, бөгде зат),
тыныс алу бұлшықеттерінің зақымдануы немесе ауруы (калий жетіспеушілігі, операциядан кейінгі ауырсыну, жарақат, майлы шөгінділердің жиналуы),
тыныс алу орталығының тежелуі (апиындар, барбитураттар), дұрыс емес желдету режимі,
бронх демікпесі, эмфизема, бронхит.
Өкпенің жеткіліксіз желдетуімен pCO2 мәні 140-150 мм Hg жетуі мүмкін.
Компенсация
Бүйрек
ацидо- және аммониогенезді күшейту, НСО3– иондарының қорын құру және артық Н+ иондарын жою;
HCO3– максималды мүмкін реабсорбциясы.
Тыныс алу жүйесі
Тыныс алу жүйесі бұзылыстарды өтей алмайды, өйткені оның проблемалары ацидоздың себебі болып табылады.
Сүйек
Сүйек тінінде H + иондарының орнына кальций иондары қанға бөлінеді.
Буферлік жүйелер
Бикарбонатты және гемоглобинді буферлеу жүйелері тиімсіз, өйткені өкпенің дисфункциясы бар
Тыныс алу алколозы
Себептер
1. Эндогендік синтез және қанға НСО3– иондарының секрециясының жоғарылауы:
тынымсыз құсу, асқазанның фистуласы, ішек өтімсіздігі бар асқазанның шырышты қабаты жасушалары,
гиповолемиямен, ілмектік диуретиктерді (фуросемид, тиазидтер) қолдану арқылы альдостерон әсерінің күшеюі нәтижесінде бүйрек эпителийінің жасушалары.
2. НСО3– жоғары концентрациясының ятрогендік түзілуі
сілтілі ерітінділерді көктамыр ішіне енгізумен, постгиперкапникалық күйде.
Компенсация
Тыныс алу жүйесі
Тыныс алу жүйесі тыныс алу жиілігі мен тереңдігінің төмендеуіне жауап береді. Гиповентиляция pCO2 концентрациясын арттырады. Осыған байланысты карбонатты буфердің сыйымдылығы артады және НСО3– / Н2СО3 қатынасы 20:1-ге дейін қалыпқа келеді.
Бүйрек
НСО3– реабсорбциялау қабілеті шекті мәнге ие болғандықтан, алкалоз кезінде және біріншілік несептегі артық НСО3– сүзілу кезінде НСО3– соңғы несепте жиналады, бұл оның рН-ын 8-ге дейін арттыруы мүмкін.
Газдардың қан арқылы тасымалдануы.
Қандағы газдар физикалық еріген және химиялық байланысқан екі негізгі күйде болады. О және СО-ның физикалық еруі: Генри заңына бағынады, оған сәйкес сұйықтықта еріген газ мөлшері оның парциалды қысымына пропорционал. Сондықтан қандағы физикалық еріген O мазмұны өте аз СО ерігіштігімен байланысты. шамамен 20 есе жоғары. Формада еріген O мөлшерінің ерігіштігі өте шамалы.Олардың мәндері химиялық байланысқан мөлшерден әлдеқайда артта. Қалыпты физиологиялық жағдайда (Р.0,% 3D 100 мм с.б.) 0,31 мл O 100 мл қанда ериді.эритроцит ішінде.
Гемоглобин қызыл пигменті, адам мен жануарлардың қаны Бір эритроцитте шамамен 340 000 000 гемоглобин молекуласы болады деп есептелінеді, олардың әрқайсысы шамамен 10 ° атомнан тұратын 8 адам қанында орта есеппен 14,5% гемагпобин бар eo жалпы мөлшері -750 г Гемоглобин күрделі. гемапротеиндер тобына жататын белок, оның құрамындағы протеиндік компонент глобин белокты емес протездік топпен ұсынылған Гемоглобин молекуласының протездік тобы асыл тастар деп аталатын 4 бірдей қосылыспен ұсынылған (54-56-сурет) темір атомы тақырыптың ортасында орналасқан және қанға тән қызыл түс береді, оның тотығу дәрежесі оттегінің қосылуына немесе бөлінуіне қарамастан тозбайды
12.Қан арқылы оттегінің тасымалдануы
Гемоглобиннің әрбір граммы 1,39 мл оттегін байланыстыруға қабілетті, сондықтан гемоглобин мөлшері 150 г/л болса, әрбір 100 мл қанда 20,8 мл оттегі болады. Толық қаныққан кезде гемоглобинмен байланысуы мүмкін оттегінің мөлшерін көрсететін мөлшер гемоглобиннің оттегі сыйымдылығы деп аталады.Егер сіз оттегінің әртүрлі концентрациясы бар қан үлгілерін алсаңыз, онда газ бен қан арасындағы тепе-теңдікке жеткенде, сіз қандағы PO, газ және O мазмұнын өлшей алады және O: қанмен байланыстыру қисығын ала алады.
Қанның тыныс алу функциясының тағы бір көрсеткіші - қандағы оттегінің мөлшері, ол оттегінің шынайы мөлшерін көрсетеді. гемоглобинмен байланысты және плазмада физикалық еріген. 100мл артериялық қанда қалыпты жағдайда 19-20мл оттегі болады, веноздық қанның бірдей көлемінде 13-15мл оттегі болады, ал артерио-веноздық айырмашылық 5-6мл гемоглобинмен байланысты оттегі мөлшерінің қатынасы. соңғысының оттегі сыйымдылығына гемоглобиннің оттегімен қанығу дәрежесінің көрсеткіші болып табылады: сау адамдарда артерия қанының гемоглобинінің оттегімен қанығуы 86% құрайды. Өкпеде оксиэмовлобиннің түзілуі және оның ұлпаларда қалпына келуі қандағы оттегінің ішінара кернеуіне байланысты.Ол жоғарылағанда гемогпобиннің қанығуының оттегімен қанығуы жоғарылайды, азайған сайын төмендейді.Бұл қатынас сызықты емес табиғат және S-тәрізді пішіні бар оксигемоглобиннің диссоциация қисығымен өрнектеледі.
Белгілі бір жағдайларда оксигемоглобиннің диссоциация қисығы рН өзгеруінің әсерінен S-тәрізді пішінін сақтай отырып, бір бағытқа немесе басқаға ауысуы мүмкін. CO2 кернеуі. дене температурасы, гемоглобиннің оттегімен байланысу қабілеті жұмыс істейтін бұлшықеттерде интенсивті метаболизм нәтижесінде СО және сүт қышқылының түзілуі артады, ал жылу өндірісі де жоғарылайды Осы факторлардың барлығы гемоглобиннің оттегіге жақындығын төмендетеді. Оксигемоглобин және тіндердің оттегін тұтыну мүмкіндігі артады. Температураның төмендеуі, СО кернеуінің төмендеуі және диссоциациясы солға ығысады, гемоглобиннің оттегіге жақындығы артады, нәтижесінде тіндерге оттегінің жеткізілуі төмендейді.
13.Көмірқышқыл газының қанмен тасымалдануы
Метаболизм өнімі бола отырып, СО2 организмде дисперсті және байланысқан күйде болады. CO ерігіштік коэффициенті 0,231 ммоль-Л кПа «(0,0308 ммоль-Л» мм сын. бағ.), бұл оттегінен 20 есе дерлік жоғары. Алайда еріген күйінде ол химиялық байланысқан күйде, негізінен бикарбонаттар түрінде, сондай-ақ ақуыздармен (карбамин немесе карбоксиді қосылыстар деп аталатын) бірге тасымалданады. Артериялық қанда СО кернеуі. 5,3 кПа (40 мм рт.ст.), оның аралық сұйықтықтағы кернеуі 8,0-10,7 кЛа (60-80 мм рт.ст.). Осы градиенттердің арқасында ұлпаларда түзілген СО2 интерстициальды сұйықтықтан қан плазмасына, одан эритроциттерге өтеді.СО сумен реакцияға түсіп, көміртек оксиді, СО, + Н.0 "" НСО, Бұл реакция ұлпа капиллярларында да қайтымды, негізінен НСО түзілуіне қарай жүреді, Плазма атада реакция баяу жүреді, бірақ эритроциттерде фермент әсерінен көмір қышқылының түзілуі СО реакциясын жылдамдатады. ылғалдану 15000-20000 есе. Көмір қышқылы HCO иондарына диссоциацияланады.НСО иондарының мөлшері жоғарылағанда олар эритроциттен плазмаға диффузияланады, ал n иондары эритроцитте қалады, өйткені. эритроциттердің мембранасы катиондарды салыстырмалы түрде өткізбейді. NSO иондарының шығымы. плазмадағы хлор иондарының плазмадан түсуімен теңестіріледі. Бұл кезде плазмада натрий иондары бөлінеді, олар эритроциттен келетін NSOZ иондарымен байланысып, NaHCO3- гемоглобин және плазма ақуыздарын түзеді, әлсіз қышқылдардың қасиетін көрсетеді, эритроциттерде калиймен, плазмада натриймен сопи түзеді. . Көмір қышқылының қышқылдық қасиеті күштірек, сондықтан белоктар әрекеттескенде Н ионы белок анионымен байланысады, ал НСОЗ ионы сәйкес катионмен бикарбонат түзеді (NaHCO3 плазмасында, эритроцитте KHCO3)
14.Қан мен ұлпалар арасындағы газ алмасу.
Ұлпалардағы газ алмасу (газдардың ұлпадағы диффузиясы) өкпедегідей заңдылыққа бағынады. Ол Фик заңымен толық сипатталған.
Оттегінің, көмірқышқыл газының және азоттың ұлпаларда диффузиясы мынада жүреді: ағып жатқан қан арасындағы осы газдардың концентрация градиентінің әрекеті капиллярлар және жасушааралық сұйықтық. Концентрация градиенттерін бағалау қандағы және интерстицийдегі әрбір газдың кернеулерінің шамасын, сондай-ақ жолын білу керек.
Олардың диффузиясы кезінде газдар жеңеді.
Тіндік капиллярлардың қанындағы 02 кернеуі 100 мм сын.бағ. дейін. Өнер. (13.3кПа), жасушааралық ортада – 20-дан 45 мм рт.ст. Өнер. (2,7-6,0 кПа). Кернеу айырмашылығы капилляр қабырғасындағы оттегі 55-80 мм сын.бағ. аралығында болады. Өнер. (7,3-10,6 кПа). Көмірқышқыл газының кернеуі: қанда - шамамен 40 мм рт.ст. Өнер. (5,3 КПа), интерстицийде - бастап
60-80 мм сын.бағ Өнер. (8,0-10,7 кПа). Қан тамырларыныңқабырғасындағы СО2 кернеуінің айырмашылығы капилляр 20-40 мм сын.бағ. жетеді. Өнер. (2,7-5,3 кПа). СО2 таралатындықтан оттегіге қарағанда шамамен 20 есе жылдамырақ, СО2 шығару әлдеқайда оңай оттегімен қамтамасыз ету.
Тіндерде газ алмасуға тыныс алу органдарының кернеу градиенттері ғана әсер етпейді қан мен интерстициальды сұйықтық арасындағы газдар, сонымен қатар алмасу аймағы беті, диффузиялық қашықтықтың мәні және олардың диффузиялық коэффициенттері газдарды тасымалдау жүзеге асырылатын орталар. Газдардың диффузиялық жолы қысқа, капиллярлық желінің ауданы үлкенірек. 1 мм2 үшін жалпы капиллярлық қабаттың беті, мысалы, қаңқа бұлшықетінде 60 м жетеді 2 және ішінде миокард - 100 м диффузия аймағы қызыл қан жасушаларының санын да анықтайды, таралуына байланысты уақыт бірлігінде капиллярлар арқылы ағу
микротамырлардағы қан ағымы. 02-нің қаннан тінге шығуына конвекция әсер етеді плазма мен интерстициальды сұйықтық, сонымен қатар эритроциттер мен жасушалардағы цитоплазма маталар. 02 тінде диффузияны ұлпа процесінде жасушалар тұтынады тыныс алу, сондықтан оның қан арасындағы кернеу айырмашылығы, интерстициалды сұйықтық пен жасушалар үнемі болады, бұл олардың диффузиясын қамтамасыз етеді
. Тіндердің оттегін тұтынуының жоғарылауымен, оның қандағы кернеуі
төмендейді, бұл оксигемоглобиннің диссоциациясын жеңілдетеді.
15.Тыныс алу жүйесінің биохимиялық негіздері.
Кез келген заманауи адам, тіпті биология мен медицинадан өте алыс болса да, өкпенің адам мен жануарлар ағзасындағы рөлі ауадағы оттегін сіңірумен және денеден көмірқышқыл газын шығарумен байланысты екенін еш ойланбастан айтады. Оттегінің органикалық заттардың молекулаларынан энергия алу үшін тірі табиғат таңдаған тотықтырғыш ретіндегі рөлін ашатынына қарамастан, тіндердің оттегін пайдалану процестері туралы ештеңе білетін адамдар айтарлықтай аз.
Аэробты организмдер үшін энергияның бірден-бір көзі тағамнан көмірсулардың, майлардың және ақуыздардың тотығуы болып табылады. Бұл процесс оттегі болмаған кезде де болуы мүмкін, яғни. анаэробты жағдайда. Дегенмен, анаэробты ыдырау энергетикалық жағынан тиімсіз, өйткені оның өнімдері әлі толық тотыға қоймаған және олардың молекулаларында энергияның көп мөлшері сақталады. Атап айтқанда, глюкозаның анаэробты ыдырауы кезінде, нәтижесінде сүт қышқылы түзіледі, бөлінетін энергия мөлшері 2 моль АТФ сақтауға мүмкіндік береді, ал глюкозаның СО2 және Н2О толық тотыққан өнімдеріне дейін аэробты ыдырауы 36 моль синтездеуге мүмкіндік береді. бір моль глюкозаға АТФ.
Жансыз табиғаттағы жану реакциясынан айырмашылығы, мысалы, автомобиль қозғалтқышында бензиннің құрамына кіретін көмірсутектердің жануы, тірі организмдердегі заттардың аэробты ыдырауы кезінде оттегі тотығатын органикалық молекулалардың құрамына кірмейді. Кребс цикліндегі ацетил КоА-ның ыдырау реакцияларының ешқайсысы оттегінің жетіспеушілігі жағдайында цикл жұмысының баяулауына қарамастан, оттегінің сіңірілуімен байланысты емес. Заттардың аэробты ыдырауы кезінде, егер ол тотығатын молекулалармен тікелей әрекеттеспесе, оттегі не үшін қажет? Тірі жасушада оттегі қанттардың, май қышқылдарының және амин қышқылдарының метаболиттерінен дегидрогеназа коферменттері (NAD+, FAD) қабылдаған сутегі атомдарын тотықтыру үшін қолданылады. Сутектің молекулярлық оттегімен тотығуы ішкі митохондриялық мембрананың тыныс алу тізбегінде жүреді және жасушадағы оттегін пайдаланудың оксидазалық жолын білдіреді. Тыныс алу тізбегіндегі сутегінің тотығуы кезінде бөлінетін энергия тотығу фосфорлану кезінде АТФ-ның энергияға бай фосфаттық байланыстары түрінде сақталатындықтан және жасуша өмірлік белсенділіктің әртүрлі көріністеріне жұмсалатындықтан, оксидаза жолы энергетикалық маңызы бар. Осылайша, тірі организмдердің оттегін пайдалануының көп сатылы процесінде өкпе бастапқы кезеңде атмосфералық ауадан оттегін сіңіреді, ал соңғы кезеңде органикалық заттардың тотығу ыдырауының өнімі болып табылатын көмірқышқыл газын организмнен шығарады.
16.Өкпедегі зат алмасудың ерекшеліктері.
Ұзақ уақыт бойы өкпедегі метаболизмнің сипаттамалары туралы ақпарат өте шектеулі болды. Өкпенің зат алмасудағы рөлі туралы ойларымыздың дамуының артта қалуының себептері олардың құрылымының ерекшеліктерімен байланысты. Өкпе кеуде қуысының көп бөлігін алады, сонымен бірге өкпенің массасы бүкіл ағзаның массасының тек 1% құрайды. Өкпе – айналымдағы қанның бүкіл көлемі бір айналымда өтетін жалғыз орган. Өкпе массасының шамамен 30% олардың құрамындағы қанға түседі. Демек, өкпенің массасы олар арқылы өтетін қан көлеміне қатысты шамалы. Сондықтан өкпеге келетін қандағы және өкпеден ағып жатқан қандағы метаболиттердің концентрацияларының айырмашылығын зерттеу (артериовенозды айырмашылық) осы органдағы биохимиялық процестерді түсіну үшін аз нәтиже береді, өйткені айырмашылықтар өте мардымсыз. Ерекшелік - бұл қанның газдық құрамының айырмашылығы, бірақ ол өкпедегі зат алмасуды сипаттамайды, бірақ олардың газ алмасуындағы рөлі.
Өкпенің метаболизмін зерттеу in vitro эксперименттік үлгілерін жасауды талап етті. Мұндай модельдің мысалы ретінде оқшауланған перфузиялық өкпе моделін келтіруге болады.
Жасанды ортамен мүшенің перфузиясы қанды толық жууға қол жеткізуге мүмкіндік береді. Демек, өкпеге кіріп-шығатын сұйықтықтағы заттардың мөлшерінің айырмашылығы толығымен осы заттардың өкпе жасушаларындағы биохимиялық түрленулеріне байланысты. Өкпе тініндегі метаболикалық процестерді зерттеуге арналған басқа модельдер - өкпе тінінің бөлімдері, субклеткалық органеллалардың фракциялары және өкпе жасушаларының дақылдары. Оқшауланған II типті альвеолоциттердің культурасын қолдану бұл жасушалардың беттік-активті заттар өндірісіндегі рөлін егжей-тегжейлі сипаттауға мүмкіндік берді.
17.Өкпеде отын ретінде қолданылатын субстраттар
Физиологиялық жағдайларда глюкоза көптеген органдар үшін жасушалық отынның негізгі түрі болып табылады. Отын ретінде тек глюкозаны пайдаланатын ұлпалар глюкозаның міндетті тұтынушылары деп аталады. Мұндай ұлпаларға мысал ретінде жүйке ұлпасы, ішек эпителийі, бүйрек миы, эритроциттер жатады. Глюкозамен қатар тіндердің көпшілігі энергия көздері ретінде басқа субстраттар – май қышқылдары, кетон денелері, амин қышқылдары пайдалана алады. Бұл субстраттарды қолдану жасушаларда глюкоза жеткіліксіз болған жағдайда - ораза кезінде, қарқынды физикалық жұмыс кезінде, суық климатта, кейбір гормоналды бұзылулармен (қант диабеті) артады. Мұндай ұлпалар глюкозаның факультативті тұтынушылары деп аталады.
Глюкозаның міндетті емес тұтынушыларына өкпе де жатады, олар үшін глюкоза қалыпты жағдайда өкпенің негізгі энергетикалық субстраты болып табылады. Өкпенің перфузиясы бар эксперименттерде глюкозаны пайдалану жылдамдығы аэробиоз жағдайында тіннің 40 мкмоль сағ-1 г-1 құрғақ салмағын құрайды. Тиімсіз анаэробты метаболизмге көшумен глюкозаның өкпенің сіңуі 2 еседен астам артады.
Денедегі барлық мүшелердің ішінде өкпе ең көп оттегімен қаныққан орган болып табылады. Осыған байланысты өкпе тінінде анаэробты гликолиздің жоғары қарқындылығы біршама күтпеген сияқты.
Метаболизденетін глюкозаның жартысына жуығы лактатқа айналады, бұл гликолиздің маңызды рөлін және Кребс циклінің шектеулі рөлін көрсетеді.Өкпедегі глюкозаның тек 22%-ы ғана СО2 және H2O-ға дейін аэробтық ыдырауға ұшырайды. Оның ішінде көмірқышқыл газының 1/4 бөлігі пентоздық цикл реакцияларында цитоплазмада, ал 3/4 бөлігі үшкарбон қышқылы цикліндегі изоцитрат пен альфа-кетоглутараттың тотығу декарбоксилдену реакцияларында митохондрияларда түзіледі. Өкпедегі гликолиздің жоғары қарқындылығының ықтимал түсіндірмесі өкпе тінінің жасушалық құрамы бойынша өте гетерогенді және көптеген жасушаларда Кребс циклінің реакциялары жүретін митохондриялар саны аз болуы мүмкін.
Глюкоза көміртегі атомдарының едәуір бөлігі белоктар мен нуклеин қышқылдарының құрамына кіреді (18%).Глюкозаның аз бөлігі май қышқылдарына (4%), басқа липидтерге (3%) және полисахаридтерге (гликоген) (5%) айналады. .
Глюкозамен қатар өкпе май қышқылдарын тотықтыруға қабілетті.
18.Өкпенің оттегін тұтынуы
Бір немесе басқа органның оттегін тұтыну жылдамдығын біле отырып, осы органдағы метаболизмнің қарқындылығын және жалпы оның жұмысының қарқындылығын бірден бағалауға болады. Метаболикалық белсенді органдар оттегінің айтарлықтай мөлшерін тұтынады, ал белсенді емес органдар, керісінше, оттегін айтарлықтай аз тұтынады. Оттегінің көп мөлшері өкпе арқылы қанға түседі. Дегенмен, ұзақ уақыт бойы бұл түсініксіз болды - өкпенің өзі қанша оттегіні пайдаланады? Бұл сұраққа жауап өкпе тінінің перфузиялық бөлімдерімен жүргізілген тәжірибелердің нәтижелерімен берілді. Олардың оттегін қабылдау жылдамдығы құрғақ ұлпа салмағының 30-150 мкл O2 мин-1 г-1 құрайды, бұл жүрек, бүйрек, ми, қалқанша безі сияқты метаболикалық белсенді органдарға қарағанда айтарлықтай төмен. Сонымен қатар, бұл мәндер тыныштықтағы қаңқа бұлшықеттеріне және метаболикалық белсенді емес органдарға қарағанда жоғары. Осылайша, өкпенің оттегін тұтыну жылдамдығын орташа деп санауға болады. Бұл өкпе дене салмағының 1% құрайды және өкпе ағзаға түсетін оттегінің 1% пайдаланатынына сәйкес келеді. Өкпе тінінің гетерогенді және O2 тұтынуы бойынша айтарлықтай ерекшеленуі мүмкін жасушалардың әртүрлі түрлерімен ұсынылғанын есте ұстаған жөн. Атап айтқанда, оттегінің жоғары шығыны II типті альвеолоциттерге тән.
19.Өкпенің энергиямен қамтамасыз етілуіне аэробты метаболизмнің үлесі. Өкпедегі энергияға тәуелді процестер.
Өкпедегі глюкозаның көп бөлігі лактат түзу үшін анаэробты ыдырауға ұшырайтынын көрсетеді. Бұдан өкпені энергиямен қамтамасыз етуге аэробты метаболизмнің үлесі аса маңызды болмауы керек деп болжауға болады. Алайда, аэробты метаболизм ингибиторларының және митохондриялардағы тыныс алу мен фосфорланудың айырғыштарының ATP мазмұнына және перфузиялық өкпелердегі ATP / ADP қатынасына әсері бар эксперименттердің нәтижелері бұл тұжырымды растамайды және керісінше, маңызды екенін біржақты көрсетеді. Өкпенің өмірлік белсенділігін энергиямен қамтамасыз етудегі аэробты метаболизмнің рөлі (3-сурет). ...
Перфузиялық оқшауланған өкпелердегі АТФ мөлшері (құрғақ тіндердің салмағы 10 мкмоль/г) аэробты метаболизмге байланысты жұмыс істейтін көптеген тіндердегіге ұқсас және ATP/ADF қатынасы 8,5 құрайды. Тыныс алу ингибиторы СО болған кезде өкпедегі АТФ құрамы мен ATP/ADF қатынасы 2-3 есе төмендейді. Динитрофенолмен тыныс алудың және фосфорланудың ажыратқышының әсері де СО әсерінен аз дәрежеде болса да, өкпе тінінде АТФ және АТФ/АДФ индексінің азаюына әкеледі.
Өкпедегі энергияға тәуелді процестер
Өкпеде бұлшықеттің жиырылуы, ішектің немесе бүйрек түтіктерінің эпителийіндегі заттардың сіңірілуі және жүйке импульсінің өткізілуі сияқты ұшпа процестер болмайды. Өкпенің негізгі қызметі - газды тасымалдау энергиямен қамтамасыз етуді қажет етпейді. «Тыныс алудың биофизикалық негіздері» тарауында альвеолярлы ауа мен қан арасындағы газдардың тасымалдануы қарапайым диффузиямен басқарылатын процесс екендігі айтылды. Соған қарамастан, өкпеде өтетін бірқатар процестер, мысалы, бронхтардағы кірпікті тасымалдау, альвеолярлы макрофагтардың фагоцитарлық белсенділігі, бронхтың тегіс бұлшықет элементтерінің жиырылуы, негізгі беттік белсенді заттың - дипальмитойлфосфатидилхолиннің синтезі және оның альвеолярлы қуысына секрециясы. қарқынды энергиямен қамтамасыз етуді талап етеді және гипоксия жағдайында баяулайды
20.Сурфактант жүйесі. Сурфактант функциялары. Сурфактанттың негізгі компоненттері.
Беттік-активті зат – су-ауа шекарасында орналасқан липопротеидті беттік белсенді зат, альвеолаларды ішкі жағынан қаптайды және беттік керілуді азайту арқылы дем шығару кезінде альвеолалардың құлауын болдырмайды. Сурфактантты өкпе жасушаларының ерекше түрі – II типті альвеолоциттер бөледі. Туа біткен беттік-активті заттардың жетіспеушілігі жаңа туған нәрестелерде өкпенің ашылмауымен көрінеді - респираторлық дистресс синдромы (РДС).
Беттік белсенді заттың негізгі компоненттері фосфолипидтер, ақуыздар және ішінара полисахаридтер. 1-кестеде беттік белсенді заттың химиялық құрамы туралы ақпарат берілген.
Липидтер
Липидтер БАЗ массасының 90%-дан астамын құрайды және бұл мөлшердің жартысы қаныққан глицерофосфолипидке – дипальмитойлфосфатидилхолинге келеді. Беттік белсенді заттың екінші компоненті басқа глицерофосфолипид – фосфатидилглицерин, оның құрамында негізінен қанықпаған май қышқылдарының қалдықтары бар. Беттік активті фосфолипидтер альвеолалардың ішкі қабырғасында беттік моноқабатты құрайды (4-сурет). Фосфолипидтердің полярлы бастарының сулы фазаға, ал гидрофобты май қышқылының құйрықтары ауаға бағытталуына байланысты су-ауа шекарасында беттік керілудің төмендеуі байқалады. Аз мөлшерде беттік белсенді заттың құрамында бейтарап липидтер мен холестерин бар. Барлық беттік-белсенді липидтер II типті альвеолоциттер арқылы синтезделеді, оларда пластинкалы денелер деп аталатын секреторлық органоидтарда жиналады.
Ақуыздар
Белоктар беттік белсенді зат массасының 10% құрайды. Осы мөлшердің ішінде белоктардың жартысы қан плазмасының ақуыздары, ал екінші жартысы беттік белсенді заттың меншікті белоктары – А (СП-А), В (СП-В), С (СП-С) және Д (СП) аполипопротеидтері. -D). Ағылшын тіліндегі SP аббревиатурасы беттік-белсенді зат ақуызы сөз тіркесімінен шыққан, яғни. беттік белсенді зат ақуызы.
• SP-A және SP-D бактериялар бетіндегі көмірсутекті домендерді тану және байланыстыру қабілетіне байланысты туа біткен иммундық жауаптарға қатысады. Байланысу макрофагтар арқылы микроорганизмдердің фагоцитозымен бірге жүреді. SP-A теріс кері байланыс механизмі арқылы БАЗ синтезін реттеуге де қатысады.
• SP-B және SP-C – альвеолалар бетінің беттік белсенді затпен дұрыс толтырылуына ықпал ететін және өкпе қызметінің биофизикалық механизмдерін қамтамасыз ететін гидрофобты мембраналық ақуыздар. Бұл ақуыздардың туа біткен жетіспеушілігі өкпе қызметінің ауыр бұзылыстарымен бірге жүреді: SP-B тапшылығы өкпе жеткіліксіздігінің белгілерімен бірге жүреді, ал SP-C тапшылығы интерстициальды пневмонияның дамуына ықпал етеді.
Беттік белсенді зат аполипопротеидтер II типті альвеолоциттермен түзіледі. Посттрансляциялық модификациялар нәтижесінде бұл белоктар үш өлшемді құрылымға ие болады (5-сурет) және оларды пластинкалы денелер түрінде жасушаға буып-түйеді - диаметрі 1 мкм липопротеидтік сипаттағы концентрлік сақиналар.
Беттік активті заттардың функциялары
• Өкпенің серпімділігінің жоғарылауы;
• Ателектаздың алдын алу (дем шығару кезінде өкпенің құлауы);
• Иммундық қорғанысқа қатысу.
Альвеолалар сәнде судағы ауа көпіршіктері ретінде қарастырылады. Су-ауа шекарасындағы беттік керілу күштері альвеолаларды қысуға бейім. Сондықтан олардың құлауын болдырмау үшін Р альвеолаларында белгілі бір ауа қысымы қажет, оны Лаплас теңдеуі бойынша есептеуге болады, ол Р-ның беттік керілуге (γ) және альвеолалардың радиусына (r) тәуелділігін көрсетеді:
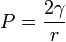
Сурфактанттың негізгі компоненттері:
Фосфолипидтер: 85%
• дипальмитойлфосфатидилхолин
• қанықпаған фосфатидилхолин
• Фосфатидилглицерин
• Фосфатидилинозит
• Фосфатидилетаноламин
• Сфигномиелин
• Басқа
Бейтарап липидтер: 5%
Холестерин, бос май қышқылдары
Ақуыз: 10%
• Беттік-белсенді зат ақуызы А
• Беттік белсенді зат протеині В
• Беттік белсенді зат протеині C
• Беттік белсенді зат ақуыз D
• Басқа
21.Өкпенің эластикалық қасиеттері.
Өкпе тінінің икемділігін өкпенің кеуде қуысының барлық көлемін алу қабілеті ретінде қарастыруға болады. Өкпенің серпімділігінде негізгі рөл БАЗ-ға тиесілі. Дем шығарудың соңғы фазасында беттік-белсенді зат қабықшасының қысылуы альвеолалар бетіндегі беттік керілудің нөлдік мәндерге дейін төмендеуімен бірге жүреді. Нәтижесінде ингаляция кезінде өкпенің ашылуы әлдеқайда жеңіл. Беттік-активті заттың беттік керілуін азайту да альвеолаларда сұйықтықтың жиналуын болдырмайды.
22.Ксенобиотикалық метаболизм.
Ингаляциялық ауамен организмге көптеген улы қосылыстар енеді. Бұл заттар тірі заттарға жат және ксенобиотиктер деп аталады. Ксенобиотиктер адамның өндірістік әрекеті нәтижесінде түзіліп, қоршаған ортаны ластайды. Көптеген ксенобиотиктер химиялық ароматты көмірсутектер немесе олардың туындылары болып табылады. Өкпе арқылы адам ағзасына түсетін спецификалық ксенобиотиктер темекі түтінінің құрамдас бөліктері болып табылады, мысалы, бензпирен, автомобильдің пайдаланылған газдары, қазандық құбырының түтіні және т.б. Физика-химиялық қасиеттері бойынша мұндай ксенобиотиктер гидрофобты қосылыстарға жатады. Ішке түскен кезде олар жасушаның гидрофобты фазасында жинақталады, яғни. биологиялық мембраналарда және организмге жағымсыз әсер етеді – канцерогендік, мутагендік, аллергендік, тератогендік және т.б.Суда нашар ерігіштікке байланысты ксенобиотиктер биологиялық сұйықтықтардың ағынымен организмнен шығарыла алмайды, т.б. жинақтаушы әсер етеді. Ксенобиотикті организмнен шығару үшін оның молекуласының полярлығын арттыру қажет. Бұған ксенобиотиктерді тотықтыру және тотығу нәтижесінде түзілген полярлы оттегі бар топтарға гидрофильді адъювантты молекулаларды қосу арқылы қол жеткізуге болады. Бауырдың детоксикация қызметі осы екі тотығу және конъюгация реакцияларының қосындысына негізделген.
Ксенобиотиктердің тотығуы (детоксикацияның бірінші фазасы) эндоплазмалық тор мембраналарының монооксигеназа жүйесі арқылы жүзеге асады. Монооксигеназа жүйесінің негізгі компоненті гемопротеин – Р-450 цитохромы болып табылады. Бұл ақуыз молекулалық оттегін белсендіреді және оттегі молекуласының атомдарының бірін ксенобиотикалық молекулаға енгізеді. Мұндай реакциялар гидроксилдену реакциялары деп аталады, өйткені олардың нәтижесінде гидрофобты молекулада полярлы гидроксил топтары пайда болады
Детоксикацияның екінші фазасында трансфераза ферменттері тотыққан ксенобиотиктің гидрофильді адъювант молекуласымен конъюгация реакциясын жүзеге асырады, оның рөлін глюкурон және күкірт қышқылы, амин қышқылдары және кейбір басқа заттар атқарады.
Әртүрлі ксенобиотиктер үшін Р-450 цитохромының спецификалық формалары бар, ал ксенобиотик ағзаға түскенде сәйкес ген экспрессияланады және берілген ксенобиотикке тән Р-450 цитохромының формасы синтезделеді. P-450 цитохромдарының супертұқымы бірнеше ондаған гендермен кодталатын бірнеше отбасын біріктіреді.
Ең күшті монооксигеназа жүйесі бауыр жасушаларында болады. Өкпеде монооксигеназа жүйесі де бар және оның сыйымдылығы бауырдың монооксигеназа жүйесінің сыйымдылығының шамамен 25% құрайды. Демек, ағзадағы ксенобиотиктерді бейтараптандыруға өкпенің үлесі де өте маңызды
23.Биологиялық белсенді заттардың (гормондар және гормоналды емес биорегуляторлар) және дәрілік қосылыстардың алмасуы.
Өкпе екі механизм арқылы биологиялық белсенді заттар (БАС) мен дәрілік қосылыстардың алмасуына белсенді қатысады:
қанда айналатын БАС және препараттарды ұстап, оларды биологиялық белсенділіктің өзгеруімен жүретін химиялық өзгерістерге ұшырату;
бірқатар биологиялық белсенді заттарды синтездейді.
Бірқатар биологиялық белсенді заттар мен препараттар өкпе арқылы инактивацияланатыны анықталды. Кейбір биологиялық белсенді заттар, керісінше, өкпедегі метаболикалық өзгерістер нәтижесінде белсендіріледі. Өкпе липидті табиғаттың биорегуляторларын – простагландиндерді синтездейді. Ақырында, кейбір биологиялық белсенді заттарда өкпе арқылы өткеннен кейін биологиялық белсенділік өзгермейді.
Өкпенің биологиялық белсенді заттар мен дәрілік заттардың алмасуындағы рөлі.
|
 Скачать 60.74 Kb.
Скачать 60.74 Kb.