/ — головной мозг, 2—окологлоточный коннектив, 3—ганглий брюшной нервной лестницы, 4 — нервы сегмента, 5 — нефридий, 6 — рот, 7 — целом, 8 — кишка, 9 — диссепимент, 10 — мезентерий, 11 — пищевод, 12 — ротовая полость, 13 — глотка, /4 — мускулы — ретракторы глотки, 15 — кольцевая и продольная мускулатура, 16 —обонятельный орган, 17 — глаз, 18 — яичник, 19 и 20 — спинной и брюшной кровеносные сосуды, 21 — сплетение сосудов на кишечнике, 22 — кольцевой сосуд, 23 — сосуд глотки, 24 — пальп
255
сегментами стенки соседних целомических мешков образуют двухслойную поперечную перегородку — септу, или диссепимент, пересекающий полость тела. Септами целом делится на известное число поперечных участков, обычно соответствующих числу сегментов тела. Впрочем, в некоторых отделах тела септы могут иногда редуцироваться. Целом заполнен водянистой жидкостью, в которой плавают амебоидные клетки.
Функции целома многообразны, прежде всего он имеет опорное значение. Целомическая жидкость, по составу близкая к воде, представляет практически несжимаемое тело; при сокращении кожно-мускульного мешка стенки тела давят на целомическую жидкость, и все тело приобретает значительную упругость (ригидность). При вытягивании тела, при его сокращении, рытье норок и других типах движения значительное количество целомической жидкости перегоняется из одних отделов тела в другие (это возможно при отсутствии диссепиментов в некоторых сегментах или при наличии в них отверстий). Другие функции целома — распределительная, выделительная и половая. В целом поступают питательные вещества из кишечника, которые затем распределяются в организме. В целом поступают и в нем накапливаются жидкие и твердые экскреты — продукты обмена веществ. Наконец, в целоме происходит созревание половых клеток — спермиев и яиц.
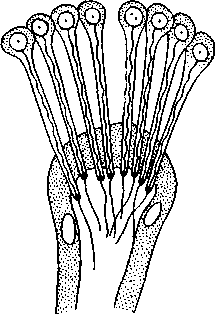
Пищеварительная система начинается ртом, который лежит на брюшной стороне перистомиума. Кишечник состоит из эктодермальной передней, энтодермальной средней и эктодермальной задней кишок. Последняя заканчивается на анальной лопасти порошицей (см. рис. 212). Передняя кишка нередко состоит из нескольких отделов. Передний из них — небольшая ротовая полость, за которой следует очень мускулистая глотка. У многих хищных видов бродячих полихет на внутренней стенке глотки кутикула местами сильно утолщается и образует острые хитиноидные зубцы, или челюстные пластинки; зубцы при выворачивании глотки служат для схватывания добычи. У нехищных сидячих форм глотка развита слабо. Средняя кишка имеет обыкновенно вид прямой трубки, задняя кишка короткая. Бродячие полихеты — преимущественно хищники и питаются различными мелкими животными; сидячие питаются главным образом взвешенными в воде органическими частицами и мелкими организмами, причем у многих для собирания пищевых частиц и для подгона их ко рту служат сильно развитые перистые головные щупики (пальпы). Они же выполняют роль жабр.
Органы дыхания многощетинковых довольно разнообразны. Наиболее просто организованные представители дышат всей поверхностью тела подобно низшим червям. У большинства функцию дыхания принимают на себя известные участки параподий. Часто спинной усик превращается в жабру (см. рис. 215, В).Внутрь ее заходят кровеносные сосуды, а через стенки растворенный в воде кислород поступает в кровь. Жабры имеют листовидную, перистую или кустистую форму и развиваются большей частью не на всем протяжении тела, а только на известном его участке.
Кровеносная система (см. рис. 217, В),как правило, состоит из двух главных продольных сосудов — спинного и брюшного. Один проходит вдоль всего тела над кишкой, другой — под нею, и оба залегают между двумя слоями спинного и брюшного мезентериев. Полость кровеносных сосудов представляет остатки первичной полости тела. Оба сосуда сообщаются многочисленными мелкими сосудами и лакунами, проходящими под перитонеальным эпителием кишки и кольцевыми сосудами, огибающими в стенке тела целом. Кольцевые сосуды расположены метамерно, иногда
256
по нескольку в каждом сегменте. К ним относятся сосуды, идущие к жабрам и возвращающие кровь от жабр и от нефридиев, где кровь освобождается от продуктов распада. Главные сосуды и их ветви образуют в конце концов сеть тончайших капилляров. Последняя так густа, что, например, у пескожила (Arenicola) в ней насчитывается до 6000 ячеек на 1 мм2. Кровеносная система замкнута, т. е. капилляры, несущие кровь от главных сосудов к тканям тела, переходят непосредственно в систему капилляров, возвращающих кровь к главным сосудам.
Движение крови по телу обусловливается тем, что стенки спинного сосуда, а иногда и некоторых других сокращаются и ритмически пульсируют. Общее направление тока крови в спинном сосуде идет сзади наперед, в брюшном — спереди назад. По околокишечному сплетению сосудов кровь перегоняется из спинного сосуда в брюшной, по кольцевым сосудам движется из спинного сосуда в брюшной в передней части тела и в обратном направлении в задних сегментах.
Кровь нередко окрашена в красный цвет, например у пескожила (Arenicola),от присутствия в ней железосодержащего вещества, близкого к гемоглобину позвоночных. Разница лишь в том, что это вещество находится не в особых кровяных клетках, а растворено в жидкости крови. У некоторых сидячих полихет кровь зеленой окраски, зависящей от присутствия в ней хлорокруорипа, по свойствам близкого к гемоглобину.
Среди многощетинковых есть небольшое количество представителей, например семейство Glyceridae, у которых кровеносная система подверглась редукции. В таких случаях функцию крови принимает на себя жидкость цслома; иногда она розового оттенка от присутствия вещества, близкого к гемоглобину. Нет кровеносной системы и у всех олигомер-ных форм (Dinophilus, Myzostomum и др.).
Выделительная система полихет состоит из пефридиев. Обычно каждый сегмент туловища обладает одной парой выделительных каналов, поэтому нефридии также называют сстментарными органами (см. рис. 216; 217, Л). Внутренний конец каждого нефридия находится в целомиче-ской полости сегмента около его задней стенки, капал нефридия пронизывает диссепимент, проходит в полость следующего сегмента и затем открывается на боковой стороне тела наружу. Итак, каждый канал находится в двух сегментах, начинается в одном, по главной массой залегает в следующем.
Дальнейшие исследования показали, что нефридии многих полихет, несмотря на одинаковое положение, имеют различную природу. У ряда низших форм сегментарные органы представлены протонефридиями. В таких случаях па внутреннем конце выделительного канала сидит пучок тонких отростков, булавовидно вздутых на концах; концевой отдел канала как бы усажен пучком булавок, это особые жгутиковые клетки — соленоциты. Головка соленоцита представляет собой тело клетки с ядром; от клетки отходит длинный полый стебелек, или трубочка, открывающаяся в просвет главного канала прото-нефридия. От тела клетки в каналец торчит бьющий жгутик (рис. 218), иногда вместо одного жгутика имеется пучок жгутов. Как видно, соленоциты могут быть легко приравнены к звезд- п Л10 -,,
J л. рис- 218. Конец одной из
чатым клеткам нротонефридиев, а жгутики — к ветвей „ротонефридия
257
мерцательному пламени. Phyllodoce (по Гудричу)
9-763
Изучение соленоцитов с помощью электронного микроскопа показало, что они устроены совершенно сходно с терминальными клетками протонефридиев плоских червей. В обоих случаях стенка трубочки близ тела клетки имеет несколько десятков тончайших параллельных друг другу продольных щелевидных отверстий.
Функция соленоцита та же, что и у звездчатой клетки протонефридиев плоских червей, т. е. осморегуляция. Выделение продуктов обмена веществ происходит через стенки главного канала протонефридия.
У многих полихет вместо концевого пучка соленоцитов, который в данном случае атрофировался, появляется небольшое, усаженное ресничками отверстие, открывающееся в целом, нефридий превращается в метанефридий. Гомология метанефридиев протонефридиям подтверждается также развитием тех и других из эктодермы.
Однако дело осложняется тем, что у многих Polychaeta нефридии соединяются с половыми протоками. Исходной формой половых протоков у многощетинковых были, по-видимому, половые воронки, или цело-модукты, — короткие каналы мезодермальной природы, открывающиеся одним концом наружу, другим (воронкой) в целом. У некоторых полихет произошло прирастание половых воронок к главному каналу имевшегося в каждом сегменте протонефридия; получились смешанные нефридии, или нефромиксии (рис. 219), причем канал и соленоциты их происходят от протонефридия, воронка же — от полового протока (рис. 219,В).
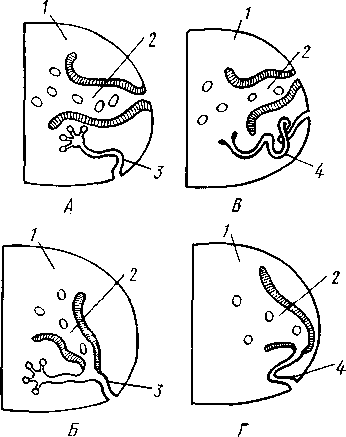
Рис. 219. Нефромиксии полихет. А — Vanadis formosa, протонефри-дий и лежащая рядом независимая от него половая воронка — це-ломодукт; Б — Podarke latifrons — половая воронка, сросшаяся с каналом метанефридия; В — Alciope contrainiu — протонефридий с приросшей к нему половой воронкой (по Гудричу):
I — половая воронка, 2—нефридиаль-ный канал, 3 — наружное отверстие
Рис. 220. Схема взаимоотношений между нсфридиями и целомодуктами у полихет. А — гипотетическая ступень с самостоятельной половой воронкой и протонефридием; Б — нефромиксии Phyllodocidae: В — Capitel-lidae, половая воронка и метанефри-днй; Г—нефромиксии большинства полихет (по Бриану):
/ — целом, 2 — половая воронка. 3 — протонефридий, 4 — метанефридий
258
В тех случаях, когда протонефридии уже превратились в метанефри-дии, дело может обстоять двояко: иногда половые протоки и метанеф-ридии остаются обособленными; у части же полихет происходит слияние половой воронки с внутренним (отверстым) концом метанефридия. В результате получаются тоже нефромиксии, но несколько иного свойства, возникающие из слияния половых воронок не с протонефридиями, а с метанефридиями (рис. 219, Б; 220, Г).Нередко, особенно у сидячих Polychaeta, число выделительных каналов сокращается; они сохраняются всего в нескольких сегментах. Обычно, у сидячих полихет происходит физиологическая дифференциация нефромиксисв на чисто выделительные и служащие для выведения половых продуктов.
Кроме нефридисв есть другие образования, играющие выделительную роль. В некоторых местах, главным образом на стенках кровеносных сосудов, перитонеальный эпителий состоит из крупных клеток, содержащих множество желтых зерен, — это накопляющиеся в клетках нерастворимые продукты обмена веществ (гуанин или соли мочевой кислоты). Наполнившись экскретами, эти, как их называют, хлорагогенные клетки отмирают, а содержимое их поступает в целом и оттуда через нефридии — наружу.
Нервная система обнаруживает ряд ступеней усложнения. Типичная центральная нервная система (см. рис. 217, А) состоит из парных мозговых ганглиев, отходящих от них и огибающих глотку двух окологлоточных коннективов и парного брюшного нервного ствола. Коннективами называются нервные стволы, соединяющие разноименные ганглии (надглоточные и подглоточные или ганглии двух соседних сегментов). Поперечные нервные стволы, связывающие ганглии одного сегмента, называются комиссурами.
У примитивных форм обе половины брюшного ствола еще широко расставлены и имеют равномерную обкладку из нервных клеток. Далее, у части Polychaeta правый и левый брюшные стволы начинают сближаться к срединной линии; в каждом сегменте на них появляется по скоплению ганглиозных клеток, и оба ганглия сообщаются поперечной перемычкой (комиссурой) —получается брюшная нервная система, метко названная нервной лестницей (см. рис. 212). У большинства полихет стволы сближаются еще больше, оба ганглия каждого сегмента сливаются, сами же стволы между ганглиями либо еще остаются разделенными, либо тоже сливаются, и тогда первичную парность брюшного ствола можно определить только на поперечных разрезах. Нервная система такого устройства называется брюшной нервной цепочкой.
Другое постепенное усложнение нервной системы заключается в ее погружении из кожного эпителия в полость тела. Во время эмбрионального развития нервная система закладывается в виде утолщения эктодермы. У некоторых Polychaeta брюшная нервная система и во взрослом состоянии остается в кожном эпителии, не меняя своего положения. У многих полихет нервные стволы уходят под кожу и даже под кожно-мускульный мешок, располагаясь в полости тела. В некоторых случаях, когда ряд сегментов сливается друг с другом (например, в области глотки), может происходить и соответственная концентрация нервных узлов.
От центральной нервной системы отходят многочисленные нервы. Головной мозг посылает от себя нервы к антеннам, пальпам и глазам, а каждый ганглий брюшной нервной цепочки иннервирует различные органы соответствующего сегмента.
Органы чувств лучше всего развиты у бродячих полихет. Кроме эпителиальных чувстительных клеток, рассеянных в коже, есть специальные
9* 259
органы осязания и химического чувства. Таковыми служат антенны, пальпы, ресничные ямки, располагающиеся на простомиуме, и чувствительные усики параподий.
Органы чувства равновесия редки, однако у некоторых полихет, особенно у сидячих, в передних сегментах тела имеется от 1 до 5 и более пар статоцистов.
Органы зрения имеются почти у всех многощетинковых. Чаще всего глаза расположены на спинной стороне простомиума в числе 2 или 4 и
8
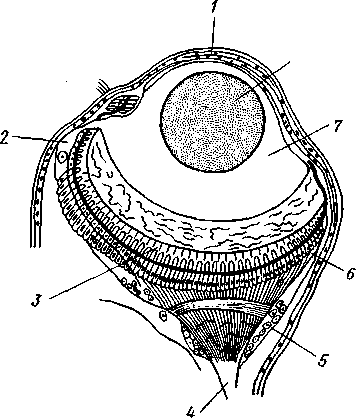
Рис. 221. Глаз полихеты Alciope (из Ливанова):
/ — роговица. 2— клетка, выделяющая стекловидное тело, 3 — светочувствительные палочки, 4 — зрительный нерв, 5 — зрительный ганглий, 6 сетчатка, 7 — стекловидное тело, S — хрусталик
иннервируются от средней части головного мозга. Эти надмозговые глаза в наиболее простом случае представляют бокаловидное впячи-вание эктодермы с суженным отверстием. Выстилающий такую глазную ямку эпителий играет роль сетчатки, а по краям впячивания переходит в обыкновенный кожный эпителий. Сетчатка слагается из клеток двух сортов. Одни из них светочувствительны, и они-то, собственно, и являются настоящими рети-нальными клетками. Каждая такая клетка на конце, обращенном в полость впячивания, т. е. к источнику света, несет светочувствительную палочку, а на другом конце продолжается в нервное волокно. Нередко, особенно у планктонных форм, например у Alciope (рис. 221), глаз сильно усложняется: обособляется от кожи, превращаясь в замкнутый глазной пузырь, а внутри него дифференцируется, кроме двояковыпуклого хрусталика, еще прозрачное стекловидное тело. Все описанные глаза необращенного, или неинвертированного, типа. Но помимо надмозговых глаз или (при их редукции) вместо них у полихет нередко развиваются глаза на других частях тела, притом обладающие иным строением. Так, у многих сидячих форм, живущих в трубках, развиваются многочисленные глаза на пальпах, превращенных в жабры. У некоторых мелких форм (АтрЫсога и др.), ползающих задним концом вперед, развиваются глазки возле порошицы. Эти «вторичные» глаза различного строения.
Половая система устроена крайне просто. Многощетинковые кольчецы раздельнополы, наружных отличий между полами нет. Половые железы формируются во всех (кроме передних и самых задних) или лишь в некоторых плодущих сегментах под слоем перитонеального эпителия. Чаще всего у основания параподий или по соседству с нефридия-ми половые клетки усиленно делятся и образуют на стенке целома местное набухание или половую железу (см. рис. 216; 217, Б). Развивающиеся гонады сначала прикрыты тонким слоем перитонеального эпителия, который впоследствии лопается, и образующиеся половые клетки попадают в целом, где они свободно плавают в полостной жидкости и достигают полной зрелости. У некоторых Polychaeta никаких половых протоков нет, так что живчики или яйца выводятся наружу просто через раз-
260
рыв стенки тела. У очень немногих полихет (сем. Capitellidae) имеются самостоятельные половые воронки с короткими выводными' каналами, открывающимися наружу (см. рис. 220, В).Наконец, в большинстве случаев эти воронки, как сказано, вступают в сообщение с нефридиями, и тогда нефридии несут двоякую функцию — выделение экскретов и выведение наружу половых продуктов. Оплодотворение обыкновенно наружное.
Рис. 222. Тихоокеанский палоло Eunice viridis (из Фаусека). Передний, более толстый отдел тела, — атокный, задний, более тонкий, — эпитокный
Нередко у полихет в тесной связи с половым размножением находится бесполое, приуроченное к периоду созревания половых продуктов. Переходом к этому размножению можно считать так называемые эпи-токные формы многощетинковых. Обычно наступление половой зрелости не отражается па организации червя. Иногда, однако, плодущие сегменты претерпевают сильную модификацию и в виде так называемой эпигокной части резко отличаются от остального атокного участка животного. Чаще всего эпитокной становится задняя половина червя. Изменения эпитокного участка сказываются в более сильном развитии параподий и щетинок, в сильной рудиментации кишечника, изменении окраски и т. д. Эпитокные половозрелые стадии некоторых представителей семейств Nereidae, Eunicidae и других поднимаются со дна и при помощи мощных веслообразных пара-подий свободно плавают, приступая к половому размножению. Таков знаменитый па-лоло Тихого океана, который периодически в несметном количестве поднимается со дна и роится в поверхностных слоях воды, представляя в это время лакомую пищу для туземцев (Eunice viridis; рис. 222). У других видов задняя половина тела отрывается от передней, регенерирует на месте разрыва новую голову и, обладая более мощными параподиями, уплывает. Бесполая половина остается на прежнем месте и регенерирует задние сегменты.
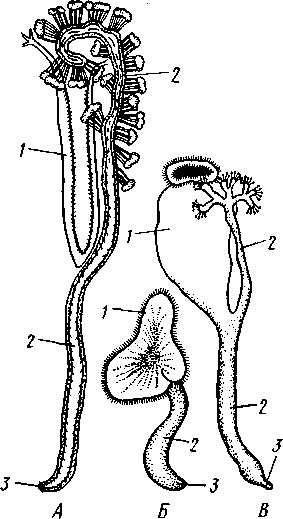
У Autolytus (рис. 223, A), Myrianida и некоторых др. эпитокная половина может еще до отделения от атокной образовать голову, а бесполая—-начать регенерацию задних сегментов: животное имеет вид двух особей разного строения, соединенных зоной регенерации. Иногда, наконец, еще до отделения первого сформировавшегося полового индивида впереди него за счет зоны регенерации образуется вторая, третья и т. д. до 30 половых особей, расположенных в один ряд (рис. 223, А);получается временная цепочка особей (Autolytus и некоторые другие). Затем половые особи отделяются и уплывают (рис. 223, Б, В).
|
 Скачать 14.77 Mb.
Скачать 14.77 Mb.