А-Догель - Зоология беспозвоночных_2. Тип губки (spongia, или porifera)
 Скачать 14.77 Mb. Скачать 14.77 Mb.
|

|
НАДРАЗДЕЛ PARAZOA ТИП ГУБКИ (SPONGIA, ИЛИ PORIFERA) Губки — многоклеточные водные, главным образом морские, неподвижно прикрепленные ко дну и подводным предметам животные. Симметрия отсутствует или имеет место неотчетливая радиальная симметрия. Органы и ткани не выражены, хотя тело построено из разнообразных клеток, выполняющих многие функции, и межклеточного вещества. Внутренние полости выстланы хоаноцитами — особыми жгутиковыми воротничковыми клетками. Нервная система отсутствует. Тело пронизано многочисленными порами и идущими от них каналами, сообщающимися с полостями, выстланными хоаноцитами. Через тело губки осуществляется непрерывный ток воды. Почти все обладают сложным минеральным (СаСО3, SiO2) или органическим скелетом. В современной фауне насчитывается около 5000 видов губок. Строение. Губки имеют форму мешка или глубокого бокала, который основанием прикреплен к субстрату, а отверстием, или устьем (osculum), обращен кверху (рис. 74). Помимо этого отверстия стенки губки пронизаны тончайшими порами, ведущими извне во внутреннюю, парагастральную полость. Тело состоит из двух слоев клеток: наружного — дермального (эктодерма) и внутреннего, выстилающего внутреннюю полость, — гастраль-ного (энтодерма). Между ними выделяется мезоглея — слой особого 101 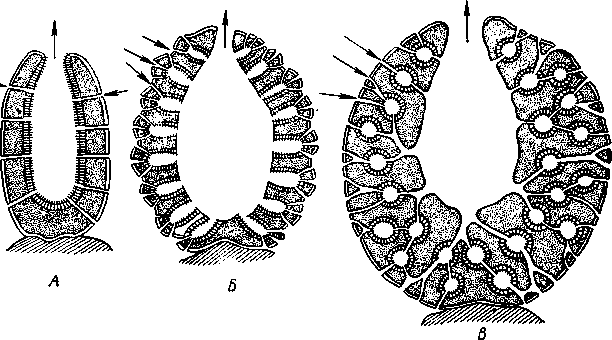 Рис. 74. Различные типы строения губок и их канальной системы (по Гессе). А — аскон; Б — сикон; В — лейкон. Стрелки показывают направление тока воды в теле губки бесструктурного вещества с отдельными разбросанными в нем клетками. У большинства губок мезоглея сильно утолщается. В мезоглее формируется также скелет. Наружный слой клеток губок в виде плоского эпителия. Мельчайшие поровые канальцы, проходящие через стенки тела губки, открываются наружу, пронизывая отдельные клетки наружно- 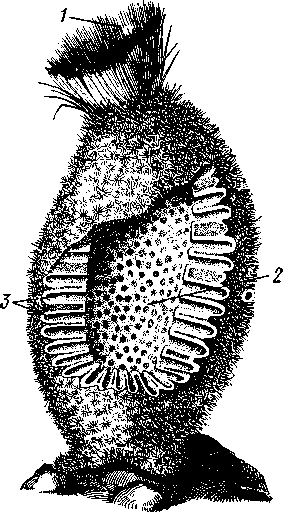 Рис. 76. Общий вид губки Sycon raphanus со вскры-^-той парагастральной полостью (по Пфуртшеллеру): ; — устье, 2 — полость тела, 3 — каналы Рис. 75. Схематический разрез через стенку тела губки типа аскона. Вверху—наружная стенка тела, внизу— парагастральная полость (по Штемпелю): / — клетки, выстилающие наружную стенку тела и стенки поровых канальцев, 2 — жгутиковые воротничковые клетки, 3 — лицевая клетка в мезоглее, 4 — склеробласт с развивающейся спикулой, 5 — пора, 6 — звездчатые клетки в мезоглее 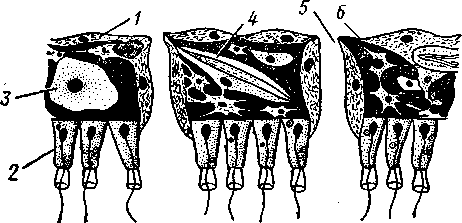 го слоя (пороциты). Гастральный слой слагается из особых воротнич-ковых клеток (хоаноцитов). Они имеют цилиндрическую форму (рис. 75), а из центра свободного, торчащего в парагастральную полость конца клетки выдается длинный жгутик, основание которого окружено цитоплазматическим воротничком. Такое строение клеток среди всех Metazoa наблюдается почти исключительно у губок, а среди Protozoa — лишь у Choanoflagellata, или воротничковых жгутиконосцев. Электронно-микроскопическое исследование хоаноцитов показало, что их тонкое строение полностью совпадает с таковым Choanoflagellata. Наиболее простую форму строения губок называют типом аскон. Однако у большинства видов эта стадия преходяща и характерна только для молодых особей. Усложнение во время индивидуального развития приводит к возникновению форм типа сикон (см. рис. 74,Б) или, если этот процесс идет еще дальше, к формам типа лейкон' (см. рис. 74,В).Эти понятия обозначают неодинаковую сложность организации губок разных групп и не соответствуют систематическим подразделениям. Усложнение заключается главным образом в том, что мезоглея сильно утолщается и вся слагающаяся из хоаноцитов энтодерма, которая у губок типа аскон выстилает парагастральную полость (см. рис. 74,Л), перемещается (как бы вдавливается) внутрь мезоглеи, образуя здесь жгутиковые карманы (у сиконов, см. рис. 74,Б) или округлые Ascon, Sycon и Leucon — роды губок, имеющие описанное строение. 102 небольшие жгутиковые камеры (у лейконов, см. рис. 74,В, рис. 77). При этом парагастральная полость изнутри у сиконов и лейконов (в отличие от асконов) оказывается выстланной плоскими клетками дермаль-ного слоя (эктодермой). Между внешней средой и парагастральной полостью связь осуществляется при помощи системы каналов, состоящей 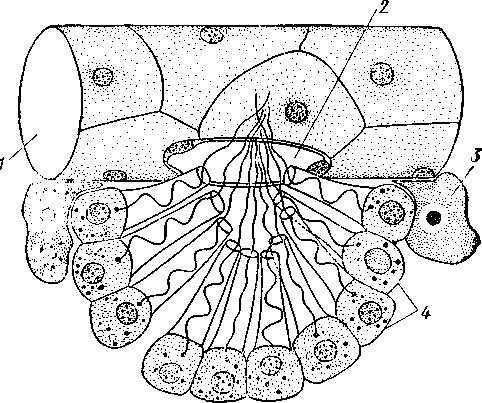 Рис. 77. Жгутиковая камера пресноводной губки Ephy-datia (по Кестнеру): / — отводящий канал, выстланный плоскими клетками, 2 — отверстие, сообщающее жгутиковую камеру с каналом, 3—архео-цит, 4 — воротничковые жгутиковые клетки (хоаноциты) из приводящих каналов, идущих от поверхности тела к жгутиковым камерам (рис. 77), и из отводящих каналов, сообщающих жгутиковые камеры с парагастральной полостью. Эти каналы представляют собой глубокие впячивания эктодермы, тогда как вся энтодерма сосредоточена в жгутиковых камерах. Число жгутиковых камер у губок велико. Например, у относительно небольшой губки Leuconia aspera (лейконоидный тип) высотой 7 см и толщиной в 1 см число жгутиковых камер превышает 2 млн. Число приводящих каналов более 80 тыс., отводящих — 5200. Клеточные элементы. В мезоглее рассеяны разнообразные клеточные элементы (см. рис. 75). Основные типы клеток следующие. Имеется значительное количество неподвижных звездчатых клеток, являющихся соединительноткаными опорными элементами (колленциты). Вторую категорию составляют склеробласты — клетки, внутри которых закладываются и развиваются отдельные скелетные элементы губок (см. ниже). В мезоглее располагается, кроме того, значительное количество подвижных клеток—амебоцитов. Среди последних можно различить клетки, внутри которых происходит переваривание пищи, воспринимаемой от хоаноцитов. Часть амебоцитов — археоцитов является недифференцированными резервными клетками, способными превращаться во все перечисленные типы клеток, а также давать начало половым клеткам. Недавними исследованиями показана широкая способность превращения одних клеточных элементов в другие, которая не наблюдается в других группах многоклеточных животных и показывает, что у губок отсутству- 103 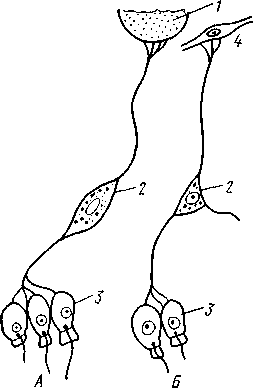 ют настоящие дифференцированные ткани. Так, хоаноциты энтодермы могут терять жгуты и уходить в мезоглею, превращаясь в амебоциты. В свою очередь, амебоциты превращаются в хоаноциты. Покровные (эк-тодермальные) клетки также могут углубляться в мезоглею, давая при этом амебоидные клетки, и т. п. Все это указывает на большую примитивность губок. Вопрос о возможности взаимопревращений одних типов клеток губок в другие изучен, однако, недостаточно. Вероятно, разные систематические группы губок в этом отношении не идентичны. Особенно широкими потенциями обладают, по-видимому, клеточные элементы известковых губок. В некоторых группах Spongia (в наибольшей степени это выражено у стеклянных губок класса Hyalo-spongia) происходит вторичное слияние почти всех клеточных элементов, что приводит к образованию синцитиев. Рис. 78. «Нервные элементы» тела губки Si/con raplia-nus (по Грассе и Тюзэ). А — «нервная клетка», осуществляющая при помощи отростков связь между стсн-koi'i канала и хоаноцитамн; Б — то же, между покрои-ной клеткой и хоаиоцнтами: 1 — прпснет к.чпала, 2—«нсри-пая клетка», ,ч— хоаноциты, 4 — покроннаи клетка Обычно принимается, что у губок совершенно отсутствует нервная система. В последнее время это утверждение поставлено под сомнение. Некоторыми зоологами в мезоглее описываются особые звездчатые клетки, соединяющиеся между собой отростками и дающие отростки к эктодерме и жгутиковым камерам. Эти клетки рассматриваются как нервные элементы, передающие раздражение (рис. 78). Однако физиологически их нервная функция никак не доказана, вероятно, эти так называемые «нервные клетки» — лишь одна из форм опорных соединительнотканых клеток (колленцитов). Физиология. Если к воде, содержащей живую губку, прибавить мелко растертую тушь, то видно, что зерна туши током воды, постоянно проходящим через канальную систему, увлекаются через поверхностные поры внутрь тела губки, проходят через каналы, попадают в парагастральную полость и через оскулум выводятся наружу. Опыт показывает путь воды и взвешенных в воде мелких частиц пищи, проходящих через тело губки. Самый ток воды через тело вызывается действием воротничковых клеток в жгутиковых камерах: жгутики клеток бьют всегда в одном направлении— к парагастралыюй полости. Количество фильтруемой через тело губки воды велико. Известковая губка Leuconia высотой 7 см за сутки пропускает через тело 22 л воды. При этом движение воды в конечных отделах канальной системы совершается со значительной силой. У Leuconia вода пз оскулум а выбрасывается на расстояние 25—50 см. Воротничковыс клетки захватывают из циркулирующей мимо воды взвешенные в пей мелкие пищевые частицы (бактерии, простейшие и т. п.) и заглатывают их. Участие хоаноцитов з процессе пищеварения может быть различно. У большинства известковых губок они не только захватывают частички пищи, но в них образуются пищеварительные вакуоли (как у простейших) и протекает внутриклеточное пищеварение. При этом лишь часть заключенной пищи передается амебоцитам мезоглеи. У других (стеклянные губки) хоапоциты только «ловят» пищу, не переваривают ее и сразу же передают амебоцитам. 104 Наконец, у некоторых видов за хоаноцитами сохраняется лишь гидрокинетическая функция (движение воды, вызываемое биением жгутиков), а пищевые частицы улавливаются непосредственно амебоцитами по ходу каналов. Утеря хоаноцитами пищеварительной функции — явление вторичное. Губки неподвижны и почти неспособны к каким бы то ни было изменениям формы тела. Только поверхностные поры могут медленно замыкаться при сокращении цитоплазмы пороцитов (с. 102). Очень медленно может сокращаться оскулярпая часть тела некоторых губок. Это происходит при сокращении особых, вытянутых в длину клеток—миоцитов. Раздражимость у губок почти ничем не обнаруживается: можно действовать на губку различными раздражителями (механическими, термическими и т. д.) —никакого внешнего эффекта не получится; это свидетельство отсутствия у губок нервной системы. Скелет. Только у немногих губок тело остается совершенно мягким, у большинства скелет твердый и служит для опоры тела и стенок канальной системы. 'Скелет состоит либо из минерального вещества: углекислой извести или кремнезема, либо из органического вещества спонгина, напоминающего своими свойствами рог, либо из сочетания кремнезема и спонгина. Помещается скелет всегда в мезоглее. Минеральный скелет состоит из микроскопических телец, игл (спп-кул), формирующихся внутри особых клеток-скслетообразоиательниц, или склсробластов (см. рис. 75). В цитоплазме склеробласта появляется маленькое зернышко, которое увеличивается, разрастается и образует правильной формы скелетную иглу. Во время роста игла окружена цитоплазмой склеробласта, которая одевает иглу тончайшим слоем. Рост происходит путем отложения па поверхности иглы новых слоев минерального вещества. Когда игла достигает предельных размеров, рост ее прекращается, склеробласт отмирает ц игла остается свободно лежать в мезоглее. Иглы обычно правильной геометрической формы и разнообразны, но могут быть сгруппированы в четыре основных типа: одноосные — в виде 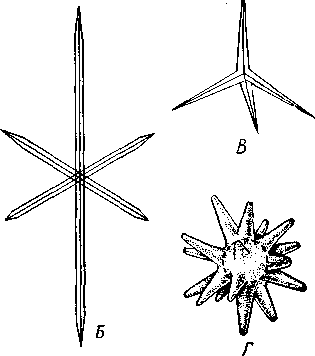  105 Рис. 79. Различные формы игл губок (по Догелю). А — одноосная игла; Б — трехосная; В — четырехосная; Г — многоосная; Д — сложная трехосная игла, или флориком стеклянных губок; Е — неправильная игла прямой или изогнутой палочки; трехосные — в виде трех взаимно пересекающихся под прямым углом лучей; четырехосные — 4 луча сходятся в центрах так, что между двумя соседними лучами образуется угол в 120°; многоосные — в виде шариков или маленьких звездочек (рис. 79).  Иглы каждого типа имеют много раз новидностей, а каждый вид губок обла- •' дает обыкновенно двумя, тремя или даже более сортами игл. В наиболее простом случае иглы лежат независимо друг от друга, у других губок иглы зацепляются концами, обра- 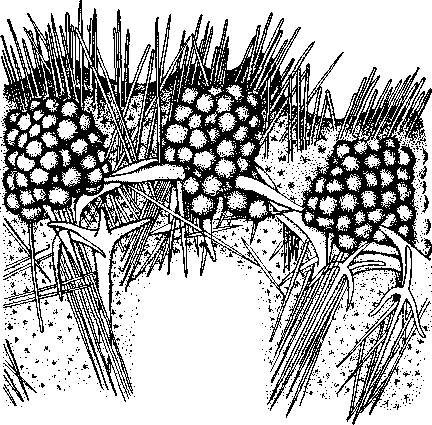 Рис. 80. Стеклянная губка Eupledel- Рис. 81. Строение скелета четырехлучевой губки la (по Шульце): (по Шульце). Разрез через поверхностный слой / _ оскулумы, 2 — базальные иглы, погру- тела, видны радиально расположенные крупные жснные в субстрат иглы и шаровидные мелкие иглы, занимающие пе риферическое положение зуя нежный решетчатый остов; иглы могут спаиваться друг с другом при помощи минерального или органического цемента, образуя сплошной скелет (рис. 80, 81). Интересно, что расположение осей в некоторых формах игл точно воспроизводит положение оптических осей в кристаллах. Так, трехосные иглы в этом отношении похожи на кристаллы правильной или кубической системы, четырехосные отвечают кристаллам гексагональной системы. Такое соответствие нередко рассматривается как выражение сходства между ростом и формированием кристаллов в неживой природе и образованием игл. Последний процесс Геккель назвал биокристаллизацией. Необходимо, однако, подчеркнуть разницу, выясняющую неправильность чисто механического толкования этих явлений. Отдельные лучи трех- и четырехлучевых игл формируются разными склеробластами и лишь позднее сливаются вместе, давая начало одной сложной игле. Между тем кристаллы образуются в маточном растворе простым наложением новых слоев минерального вещества на растущий кристаллик. Таким образом, биокристаллизация резко отличается от настоящей кри- 106 сталлизации тем регулирующим влиянием, которое на нее оказывает организм. Роговой, или спонгиновый, скелет состоит из сильно разветвляющейся внутри мезоглеи сети роговых волокон желтоватого цвета. Химический отстав спонгина близок к шелку, притом с некоторым, иногда до-  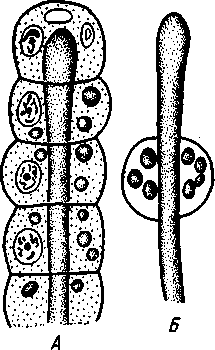 Рис. 82. Развитие спонгинового скелета. А — клетки-спонгиоблас-ты, формирующие скелетный тяж из спонгина; Б — скелетный тяж, освободившийся от клеточных элементов (по Грассс и Тюзэ) Рис. 83. Колония роговой губки Aplyslna aerophola с четырьмя оскулума- ми — / (по Пфуртшенглсру) вольно значительным (до 14%) содержанием иода. Он формируется несколько иначе, чем минеральный. Растущие волокна скелета окружены сплошным футляром из мелких клеток-скелетообразовательниц, так что образование волокон идет не внутриклеточно (как в случае игл), а межклеточно (рис. 82). Электронно-микроскопические исследования показали, что тяжи спонгина слагаются из тончайших субмикроскопических фибриллей, обладающих поперечной исчерченностью (как колла-геновые волокна в соединительной ткани высших животных). Наконец, имеются губки, совершенно лишенные скелета. Бссскелет-ные губки очень мелкие — свидетельство опорного значения скелета, без которого губки не могут разрастаться. Размножение и развитие. Губки размножаются бесполым и половым способами. Бесполое размножение носит характер почкования. На поверхности губки появляется бугор, в который продолжаются все слои тела и парагастральная полость. Этот бугор постепенно растет, на конце его прорывается новый оскулум. Полное отделение почки происходит сравнительно редко, обычно дочерние особи сохраняют связь с материнской — возникает колония (рис. 83). Границы между отдельными особями могут сглаживаться, так что вся колония сливается в общую массу. В таких колониях о числе слившихся особей можно судить по числу оскулумов. Особый способ внутреннего почкования существует у пресноводной губки бадяги. Летом бадяга размножается обыкновенным почкованием 107 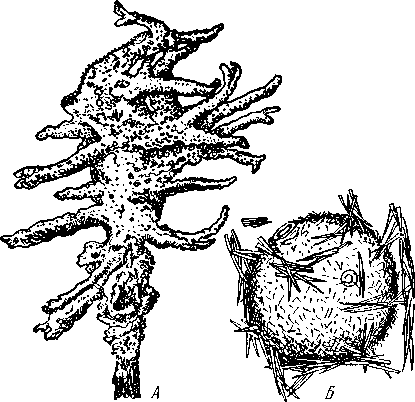 и половым путем. Но к осени в мезоглее бадяги наблюдается образование амебоидными клетками особых шаровидных скоплений — геммул (рис. 84). Геммула, или внутренняя почка, представляет многоклеточную массу, окруженную оболочкой из двух роговых слоев, между которыми имеется прослойка воздуха с мелкими кремнеземными иглами, поставленными перпендикулярно к поверхности геммулы. Зимой тело бадяги умирает и распадается, а геммулы падают на дно и, защищенные своей оболочкой, сохраняются до следующей весны. Тогда содержащаяся внутри геммулы клеточная масса выползает наружу, прикрепляется ко дну. и развивается в новую губку. Большая часть губок (в том числе все известковые губки) гермафродиты, часть видов раздельнополы. Половые клетки их происходят из амебоидных клеток (ар- Рмс. 84. Пресноводная губка бадяга Spongilla (по Речвому). /1 — общий вид губки в естественную величину; Б — отдельная геммула (увеличено) хеоцитов), ползающих в мезоглее. Они залегают в мезоглее под энтодермой жгутиковых камер. Живчики выходят в полость канальной системы, выводятся через оскулум, проникают в другие особи губок, имеющие зрелые яйца, и оплодотворяют последние. Начальные стадии развития яйца протекают внутри материнского оргапиз- 108 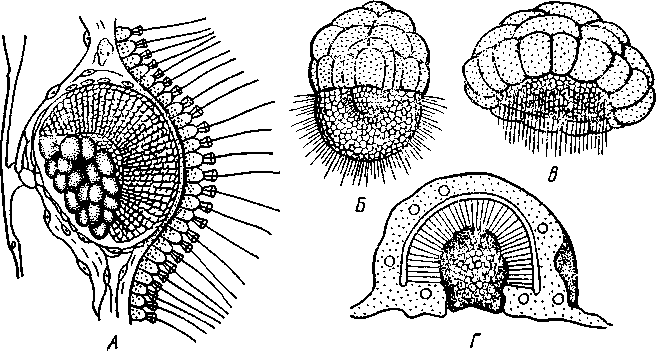 Рис. 85. Развитие известковой губки Sycon raphanus (по Шульце). А — зародыш (псевдогаструла) в теле материнской особи, крупные клетки впятились внутрь полости бластоцеля; Б — свободноплавающая амфйбластула, крупные клетки вновь выпятились; В — впячивание мелких клеток, несущих жгутики (гаструляция); Г — прикрепление и начало метаморфоза личинки ма. У части известковых губок развитие протекает следующим образом. Яйцо большей частью испытывает полное и сначала равномерное дробление, давая последовательно начало 8 бластомсрам, лежащим венчиком в одной"Чтлоскости. Далее экваториальной бороздой зародыш делится на 8 мелких верхних и 8 более крупных нижних клеток. При дальнейшем развитии мелкие бластомеры делятся быстрее крупных. Получается полый однослойный шар — бластула, у которой верхняя половина состоит из мелких цилиндрических, снабженных жгутиками клеток микромеров, а нижняя — из крупных зернистых макромеров. Вследствие различия бластомеров на полюсах бластула губок называется амфибластулой (рис. 85). Еще находясь в теле материнского организма, амфибластула претерпевает своеобразное изменение. Ее крупноклеточная половина начинает впячиваться в мелкоклеточпую, по процесс вскоре останавливается, крупные клетки выпячиваются обратно и личинка возвращается к состоянию амфибластулы. Последняя через систему каналов выходит из тела губки и через некоторое время личинка прикрепляется к субстрату тем полюсом, на котором расположены мелкие, несущие жгутики клетки. Одновременно эти клетки впячиваются внутрь бластулы и оказываются лежащими внутри зародыша, который становится на этой стадии двухслойным (рис. 85). Более крупные клетки амфибластулы образуют наружный слой. В дальнейшем внутренний слой жгутиковых клеток образует клетки жгутиковых камер губок, а наружные клетки — дермаль-ный слой, мезоглею и все ее клеточные элементы. 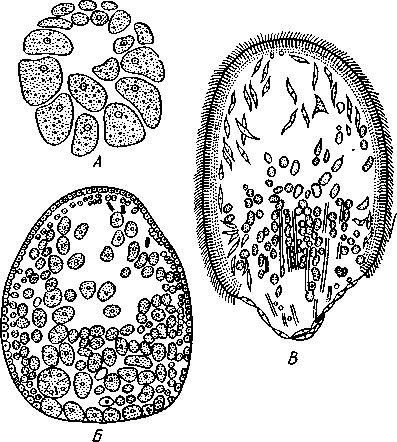 У большинства других животных, в эмбриональном развитии которых имеется бластула, слагающаяся из клеток различной величины (аналогично амфибластулс губок), более крупные клетки так называемого вегетативного полюса обычно дают начало энтодерме, мелкие же клетки анималыюго полюса — эктодерме. У губок обратные отношения. Кроме того, у губок дважды происходит впячиваиие полюсов бластулы внутрь. Существенный вопрос раз вития губок — установление мо мента гаструляции. Одни ученые не придают значения первому преходящему впячиванию амфи бластулы и называют получаю щуюся при этом стадию ложной гаструлой (псевдогаструла), а настоящей гаструляцией считают вторичное впячивание. Другие придерживаются обратной точки зрения и считают истинной га струляцией первое впячивание. Особенности эмбрионального раз вития губок дают основание уче ным считать, что у губок первич ная эктодерма (мелкие жгутико вые клетки) становится на место энтодермы, и обратно. По их мне нию, у губок произошло извраще ние зародышевых пластов. На Рис. 86. Развитие МухШа, идущее по типу этом основании зоологи дают губ- кремневых и роговых губок (по Маасу). кямнпчкянир — животные rktro- Л —дробление яйца; Б — образование ли- кам название животные, выво чинки. в —закладка элементов скелета роченные наизнанку (Enantio- (спикул) внутри паренхимулы zoa). 109 У неизвестковых губок и некоторых известковых эмбриональное раз* витие иное. У многих из них в результате дробления образуется бластула, стенки которой состоят из более или менее одинаковых клеток, снабженных жгутиками. В дальнейшем отдельные клетки стенки бластулы заползают в ее полость, которая постепенно заполняется рыхло расположенными клеточными элементами. На этой стадии личинка называется паренхимулой (рис. 86). В дальнейшем паренхимула садится на дно, ее поверхностные жгутиковые клетки погружаются внутрь и дают начало воротничковому эпителию. Клетки внутреннего пласта, напротив, выходят на поверхность и образуют покровный клеточный слой и мезоглею губки. Таким образом, извращение зародышевых пластов происходит и при этом типе развития. Вопрос о причинах, вызывающих у губок извращение зародышевых пластов, во многом еще неясен. Одна из наиболее обоснованных гипотез была высказана В. Н. Беклемишевым, который связывает этот процесс с образом жизни губок на личиночной и взрослой стадиях. Жгутиковые клетки (кинетобласт) свободноплавающих бластул губок выполняют двигательную (кинетическую) функцию. Когда личинки садятся на субстрат, то двигательная функция кинетобласта сохраняется, но она переносится внутрь тела развивающейся губки и становится мерцательно-вододвижущей, вызывающей не передвижение организма в воде, а передвижение воды в организме. По мере погружения кинетобласта внутрь другие клеточные элементы, входившие в состав тела плавающей личинки, постепенно образуют наружный слой тела губки. Таким образом извращение зародышевых пластов оказывается обусловленным изменением образа жизни животного в ходе онтогенеза. Предполагается, что эти стадии рекапитулируют соответствующие этапы филогенеза. 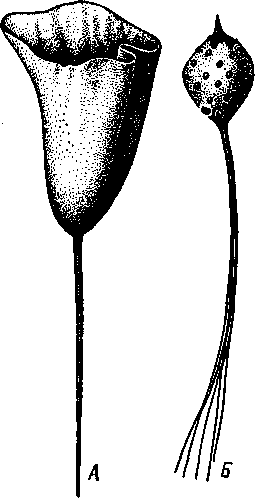 Губки в высокой степени способны к регуляции. При удалении отдельных участков тела происходит их восстановление. Если губку растереть или даже протереть через сито, то образовавшаяся кашица, состоящая из отдельных клеток и групп клеток, оказывается способной к восстановлению целого организма. При этом клетки, активно двигаясь, собираются вместе, а затем из клеточного скопления формируется маленькая губка. Процесс формирования целого организма из скопления соматических клеток называют соматическим эмбриогенезом. Рис. 87. Глубоководные губки (из Колтуна). А — dives; Б — Hyalo-пета elegans Экология и практическое значение губок. Наибольшего видового разнообразия губки достигают в тропических и субтропических зонах Мирового океана, хотя и в арктических, и субарктических водах их встречается немало. Большинство губок — обитатели небольших глубин (до 500 м). Число глубоководных губок невелико, хотя их находили на дне самых глубоких абиссальных впадин (до И км). Губки поселяются преимущественно на каменистых грунтах, что связано со способом их питания. Большое количество иловых частиц забивает канальную систему губок и делает их существование невозможным. Лишь немногие виды живут ПО на илистых грунтах. В этих случаях у них обычно имеется одна или несколько гигантских спикул, которые втыкакпся в ил и приподнимают губку над его поверхностью (например, виды родов Hyalostylus, Hyalo-пета) (рис. 87). Губки, обитающие в приливно-отливной зоне (на лито-рали), где они подвергаются действию прибоя, имеют вид наростов, подушечек, корочек и т. п. У большинства глубоководных губок скелет кремневый — прочный, но хрупкий, у мелководных губок — массивный или эластичный (роговые губки). Фильтруя через тело огромное количество воды, губки являются мощными биофильтраторами. Этим они способствуют очистке воды от механического и органического загрязнения. 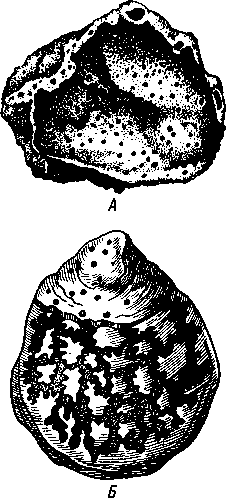 Губки часто сожительствуют с другими организмами, причем в одних случаях это сожительство носит характер простого комменсализма (квартирантство), в других приобретает характер обоюдно полезного симбиоза. Так, колонии морских губок служат местом поселения большого числа разных организмов — кольчатых червей, ракообразных, змеехвосток (иглокожие) и др. В свою очередь, губки часто поселяются на других, в том числе подвижных, животных, например на панцире крабов, раковинах брюхоногих моллюсков и т. п. Для некоторых, в особенности пресноводных губок, характерен внутриклеточный симбиоз с одноклеточными зелеными водорослями (зоохло-реллами), которые служат дополнительным источником кислорода. При избыточном развитии водоросли частично перевариваются клетками губки. кой (из Колтуна). А — по- п верхность раковины с мно- еще неясен. В растворении извести, по-види- Г0численными мому, существенную роль играет выделяемая губкой углекислота. Своеобразную экологическую группу представляют сверлящие губки (род Ciiona).Поселяясь на известковом субстрате (раковины моллюсков, колонии кораллов, известковые породы и т. п.), они образуют в нем ходы, открывающиеся наружу небольшими отверстиями (рис. 88). Через эти отверстия выступают Рис. 88. Раковины устрицы, выросты тела губки, несущие оскулумы. Меха- пораженные сверлящей губ-низм действия сверлящих губок на субстрат отверстиями, просверленными губкой; Б — ходы и каналы, проде- ,-, , ланные сверлящей губкой в Практическое значение губок невелико. толще ракновины (верхний слой раковины удален) В некоторых южных странах имеется промысел обладающих роговым скелетом туалетных губок, используемых для мытья и различных технических целей. Их ловят в Средиземном и Красном морях, Мексиканском заливе, Карибском море, Индийском океане, у берегов Австралии. Промысел стеклянных губок (преимущественно Euplectella), используемых в качестве украшений и сувениров, существует также у берегов Японии (см. рис. 87). Классификация. Классификация типа губок базируется на составе и строении скелета. Различают три класса. 111 КЛАСС I. ИЗВЕСТКОВЫЕ ГУБКИ (CALCAREA, ИЛИ CALCISPONGIA) Скелет слагается из игл углекислой извести, которые могут быть четырехосными, трехосными или одноосными. Исключительно морские, преимущественно мелководные небольшие губки. Они могут быть построены по асконоидному, сиконоидному или лейконоидному типу. Типичные представители — роды Leucosolenia, Sycon, Leuconia (см. рис. 76). КЛАСС II. СТЕКЛЯННЫЕ ГУБКИ (HYALOSPONGIA) Морские преимущественно глубоководные губки высотой до 50 см. Тело трубчатое, мешковидное, иногда в виде бокала. Почти исключительно одиночные формы сиконоидного типа. Кремневые иглы, слагаю- Рис. Разрез 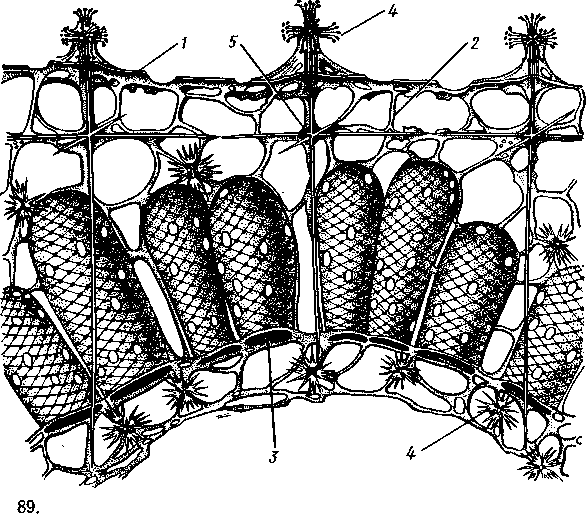 через стенку тела стеклянной губки Euplectella asper- gillum (по Шульце): 1 — поверхностный (дермальный) слой, 2 — синцитиальные перемычки в наружном слое тела, 3 — жгутиковые камеры, 4—мелкие иглы (микросклеры), 5 — крупные иглы (макросклеры) щие скелет, крайне разнообразны, в основе трехосные. Часто спаиваются концами, образуя решетки разной сложности (рис. 89). Характерная черта стеклянных губок — слабое развитие мезоглеи и слияние клеточных элементов в синцитиальные структуры. Типичный род Euplectella (см. рис. 80). У некоторых видов этого рода тело цилиндрическое, до 1 м в высоту, иглы у основания, втыкающиеся в грунт, достигают 3 м длины. КЛАСС III. ОБЫКНОВЕННЫЕ ГУБКИ (DEMOSPONGIA) К этому классу принадлежит большинство современных губок. Скелет кремневый, спонгиновый или сочетание того и другого. Сюда относится отряд четырехлучевых губок (Tetraxonia), скелет которых слагает- 112 I ся четырехосными иглами с примесью одноосных. Характерные представители: шаровидные крупные геодии (Geodia), ярко окрашенные оранжево-красные морские апельсины (Tzthya)^, комковидные яркие пробковые губки (сем. Suberitidae), сверлящие губки (сем. Clionidae) и многие другие (см. рис. 88). Второй отряд класса Demospongia — кремнероговые губки (Cornacuspongida). В состав скелета входит спонгин как единственный компонент скелета или в разных соотношениях с кремневыми иглами. Сюда принадлежат туалетные губки, немногочисленные представители пресноводных губок — бадяг из сем. Spongillidae (см. рис. 84), эндемичные байкальские губки сем. Lubomirskiidae. Филогения типа Spongia В организации губок много признаков большой примитивности: отсутствие настоящих дифференцированных тканей и органов, чрезвычайная пластичность клеточных элементов, отсутствие резко выраженной индивидуальности в колониях — все это свидетельство того, что губки— просто устроенные представители многоклеточных. Если принять теорию Мечникова о происхождении многоклеточных (с. 93), то легко видеть, что личинка, свойственная большинству губок,— паренхимула (см. рис. 86), по строению почти полностью соответствует гипотетической мечниковской фагоцителле. У нее имеется поверхностный, эктодермальный слой жгутиковых клеток и внутренний рыхлый слой клеток — энтодерма. Можно предположить, что фагоцителла перешла к сидячему образу жизни и таким путем дала качало типу губок. При этом, как уже отмечалось (с. 109), судьба клеточных слоев фагоци-теллы у губок оказалась иной, чем у прочих многоклеточных («извращение» зародышевых листков): наружный эктодермальный слой жгутиковых клеток у губок дал начало пищеварительному слою хоаноцитов, который вместе с тем осуществляет кинетическую мерцательную водо-движущую функцию; внутренние энтодермальные клетки зародыша, которые у других групп животных дают начало энтодермальной кишке, у губок превращаются в клетки поверхности тела (дермальные) и в клеточные элементы мезоглеи. Все эти факты говорят о том, что отделение губок от ствола многоклеточных произошло очень рано, еще до того, как определилась окончательная судьба двух основных клеточных пластов тела. Некоторые зоологи считают, что губки произошли от колониальных воротничковых жгутиконосцев независимо от прочих многоклеточных. Другие полагают, что многоклеточные происходят общим стволом, от которого очень рано отделились губки. Второй взгляд представляется более обоснованным потому, что личинка — паренхимула губок — сходна с планулой кишечнополостных. Это говорит об общности их происхождения. Губки — очень древние организмы. Их ископаемые остатки многочисленны в кембрийских морских отложениях. Встречаются они и в протерозойских породах. НАДРАЗДЕЛ EUMETAZOA |