|
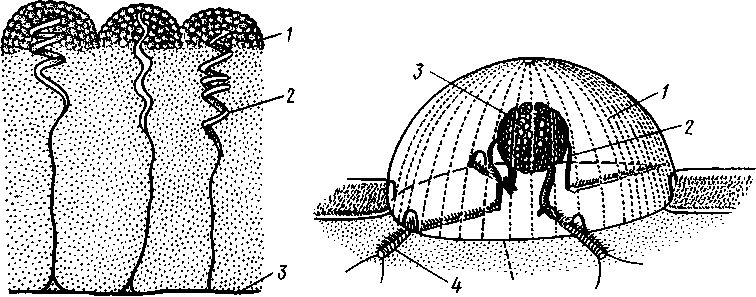
А-Догель - Зоология беспозвоночных_2. Тип губки (spongia, или porifera)
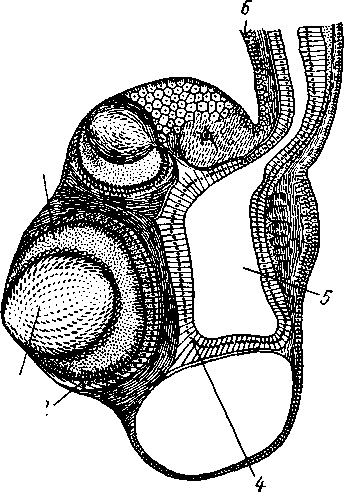
Происхождение сифонофор. При толковании строения сифонофор были высказаны две точки зрения. Одни ученые отстаивали их колониаль-ность, другие считали сифонофору единой особью, а все ее придатки лишь органами сложной особи.
Большинство современных зоологов считают сифонофор происшедшими от колоний полипов, приспособившихся к плавающему образу жизни.
Кроме истории развития, в пользу такого толкования свидетельствует и то, что в настоящее время найдено несколько плавающих колониальных полипов (в том числе один в Каспийском море — Moerisia) (рис. 104). На колониях таких гидроидов развиваются как полипоидные, так и медузоидные особи.
Можно представить себе, что сифонофоры произошли от подобных колониальных Hydroidea в результате возникновения полиморфизма особей и разделения функций между ними. При таком толковании строения сифонофоры воздушный пузырь, плавательные колокола и го-нофоры вполне легко и естественно подводятся под тип строения медуз, а кормящие полипы и пальпоны отвечают видоизмененным полипам.
КЛАСС II. СЦИФОИДНЫЕ МЕДУЗЫ (SCYPHOZOA)
К этому сравнительно небольшому классу (200 видов) относятся медузы, обитающие только в морях. Они значительно крупнее гидромедуз (рис. 105). Кроме того, они легко отличимы от последних по отсутствию паруса. В остальном у сцифомедуз типично медузоидное строение.
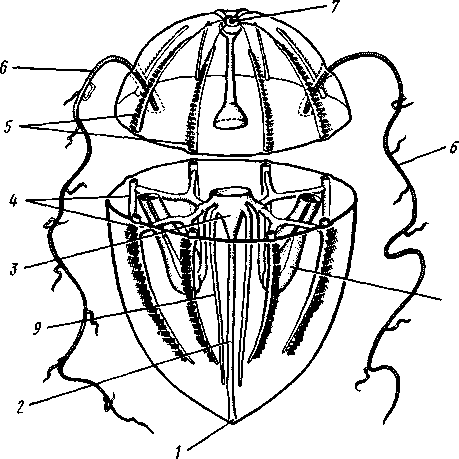
Строение и физиология. Тело в виде круглого зонтика или, при вытягивании его по главной оси, высокого колокола (рис. 106). Посреди нижней вогнутой стороны зонтика на конце ротового стебелька помещается четырехугольный рот. Углы рта вытягиваются в 4 желобовидных выро-
129
5—763
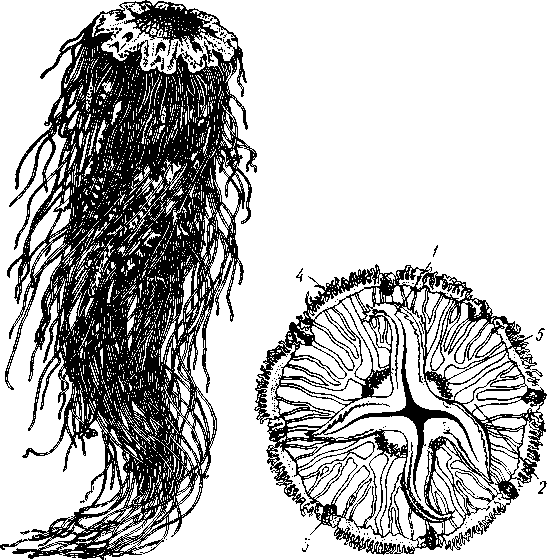
Рис. 105. Сцифоидные медузы. А — Cyanea capillata (рис. Кондакова); Б —Aurelia aurita (no Кюкенталю) :
/ — ротовая лопасть, 2 — радиальные каналы гастроваскулярной системы; 3 — ропалии, 4—щупальца по краю зонтика, 5 — гонады
 10 10
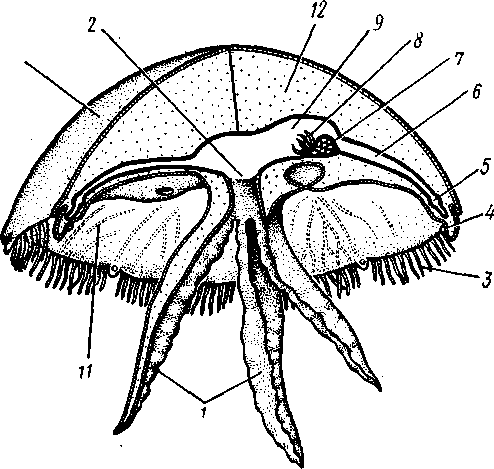
130
Рис. 106. Схема строения сцифоидной медузы (из Байера)] / — ротовые лопасти, 2—ротовое отверстие, 3 — щупальца, 4 — ро-палий, 5 — кольцевой канал, 6 — радиальный канал, 7 — гонада, 8 — гастральные нити, 9 — желудок, 10 — эксумбрелла, // — субумб-релла, 12 — мезоглея. Эктодерма показана штриховкой, энтодерма — черным
ста — ротовые лопасти, служащие для захвата пищи; у некоторых так называемых корнеротых сцифомедуз (отр. Rhizostomida) ротовые лопас-сти становятся складчатыми и срастаются, так что от ротового отверстия остаются лишь многочисленные мелкие поры, через которые проходит в кишечник пища — мелкие планктонные организмы (см. рис. 109).
J
Рот ведет в энтодермальный желудок, занимающий центр зонтика и образующий четыре неглубоких карманообразных выпячивания. В желудок вдаются с краев четыре валика с гастральными нитями, которые служат для увеличения всасывающей поверхности энтодермы.
1
От желудка расходится к краям тела система радиальных каналов. В простейшем случае их всего четыре, у других видов — восемь, у ряда сцифомедуз гастроваскуляр-ная система еще сложнее — состоит из 4 сильно ветвящихся каналов первого порядка, 4 ветвистых каналов второго порядка и 8 неразветвленных каналов третьего порядка. Каналы правильно чередуются в расположении, а своими наружными концами впадают в кольцевой канал, окаймляющий край зонтика.
Рис. 107. Продольный разрез чеоез ропалий медузы Carybdea. Налево разрез прошел через 2 главных глаза ропалия, пустое пространство внизу ропалия — статоцист (статолиты растворились при изготовлении препарата) (по Шевякову):
/ — хрусталик. 2 — светочувствительный
слой ретинальных клеток, 3 — стекловид
ное тело. 4 — энтодермальная выстилка
продолжающегося в ропалий радиального
канала (5), 6 — эктодерма ропалия
Край зонтика несет различное число щупалец. Некоторые из щупалец, расположенные у концов главных каналов первого и второго порядков, видоизменяются и превращаются в краевые тельца, или ропа-лии (рис. 107). При этом щупальца укорачиваются и утолщаются, а внутри них развиваются органы зрения « органы равновесия. Каждый ропа-лий чаще содержит один статоцист и несколько глазков разной степени
сложности строения; наряду с глазками, напоминающими таковые гидроидных медуз, здесь имеются и более сложные глаза типа глазного пузыря. Такой глаз получается посредством погружения глазной ямки под эпителий и отшнуровывания ее от поверхности тела, причем ямка замыкается под кожей в глазной пузырь. Кожный эпителий над пузырем остается тонким и прозрачным и называется роговицей. Дно и боковые стенки пузыря состоят, как и у гидромедуз, из пигментных и чувствительных клеток. Часть стенки пузыря, лежащая непосредственно под роговицей, сильно утолщается и образует двояковыпуклый хрусталик. Внутренность пузыря заполнена бесструктурным стекловидным телом, которое выделяется стенками пузыря. Несмотря на сложность строения, глаза, по мнению большинства авторов, служат лишь для различения света и тьмы. В связи с сильным развитием органов чувств центральная нервная система сцифомедуз тоже испытывает усложнение. На протяжении краевого нервного кольца (соответственно 8 ропалиям) возника-
5* 131
ют 8 скоплений нервных клеток, или ганглиев, — первый пример образования значительных нервных узлов.
Размножение и развитие. Медузы раздельнополы. Половые железы образуются из энтодермы нижней поверхности карманов желудка. Созревшие половые клетки выводятся наружу через рот медузы.
После полного и равномерного дробления яйца образуется бластула, а затем типичная мерцательная планула. Она сначала плавает, позднее же прикрепляется передним полюсом к морскому дну. На прежнем заднем, а теперь верхнем полюсе прорывается рот, который ведет внутрь образующейся к этому времени гастральной полости. Вокруг рта развивается венчик щупалец, число которых кратно четырем. Энтодерма гастральной полости дает 4 продольных желудочных валика. В результате этих изменений планула превращается в маленького одиночного полипа— сцифистому (рис. 108), немного похожего на гидру, но устроенного сложнее. Этот полип может путем почкования давать начало другим сцифистомам.
Главный процесс, совершающийся со сцифистомой, — это стробиля-ция: полип делится путем ряда поперечных перетяжек, которые постепенно врезаются с краев в тело полипа, пока из последнего не получится подобие стопки наложенных друг на друга тарелок, соединенных центральным стволом. На этой стадии развития полип называется стробилой.
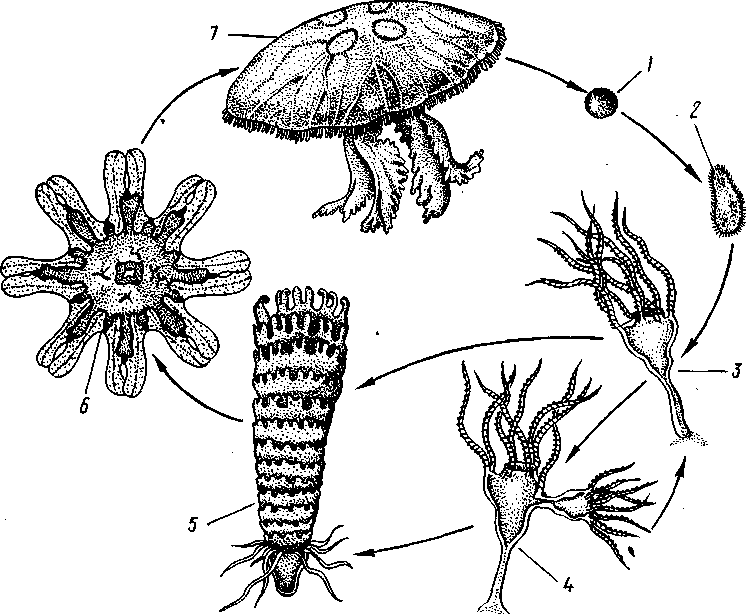
Рис. 108. Развитие сцифомедузы Aurelia:
1 — яйцо, 2— планула, 3—сцифистома, 4 — почкующаяся сцифистома, 5 — стробиляция, 6 — эфира,
7—взрослая медуза (по Ъайеру)
132
Образовавшиеся в процессе стробиляции диски представляют собой молодых медуз, расположенных вогнутыми сторонами их зонтиков кверху. Медузы постепенно, начиная с верхней, отрываются от сцифистомы, переворачиваются выпуклой стороной кверху и переходят к плавающему образу жизни. Они еще во многом отличаются от взрослых медуз, а потому считаются особой личиночной стадией, или эфирой. Край зонтика эфиры глубоко вырезан в виде 8 лопастей. Превращение во взрослую медузу сопровождается усиленным ростом и состоит главным образом в том, что края зонтика выравниваются, формируется сложная канальная система, появляются краевые щупальца и зачатки гонад.
Таким образом, сцифомедузы обладают ясно выраженным метагенезом, т. е. чередованием полового и бесполого поколений, причем в противоположность Leptolida из гидрозоев у них наиболее развито медузоид-ное поколение.
Экология. Сцифомедузы плавают при помощи сокращений зонтика, число которых может доходить до 100—140 в 1 мин. Некоторые медузы широко распространены; так, Aurelia aurita, обитающая почти во всех умеренных и тропических' морях, заходит и в арктические воды. Другие сцифоидные медузы распространены более ограниченно.
Тело большинства медуз прозрачно, что зависит от большого количества содержащейся в тканях воды (особенно в мезоглее). У многих медуз вода составляет 97,5% общей массы тела.
Размеры сцифомедуз могут быть очень велики: Aurelia aurita достигают обычно 40 см в поперечнике, тогда как Cyanea capillata — иногда почти 2 м в диаметре при длине щупалец 10—15 м. Медузы — хищники. Они питаются различными планктонными беспозвоночными, а иногда и молодью рыб.
Расправленными щупальцами медузы облавливают большую площадь воды. Так, Drymonema, достигая 25 см в поперечнике, облавливает щупальцами круг в 150 м2.
Классификация. Сцифомедузы делятся на 5 отрядов.
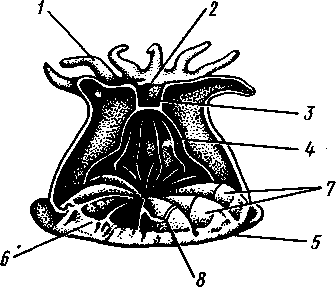
Отряд 1. Stauromedusae состоит из небольшого числа своеобразных форм, ведущих прикрепленный образ жизни. У них имеется стебелек, которым медуза прикрепляется к субстрату. Край зонтика образует глубокие вырезки, между ними на особых рукооб-разных выростах сидят пучки головчатых щупалец. Жизненный цикл Stauromedusae проходит без чередования поколений. Из планулы непосредственно развивается молодая медузка. Наиболее типичными представителями отряда могут служить Haliclystus (рис. 109, .4) и Lucernaria.
Отряд 2; Cubomedusae тоже невелик. Его представители имеют обыкновенно четырехгранный высокий зонтик с четырьмя ропалиями (см. рис. 107) и четырьмя простыми (как у Carybdea) либо разветвленными щупальцами. Кубомедузы встречаются в мелководье теплых морей. Некоторые тропические виды, частые у берегов Австралии и Индонезии (например, Chiropsalmus), могут вызывать у человека тяжелые, иногда смертельные «ожоги».
Отряд 3. Coronata. Зонтик делится кольцевой бороздкой на центральный диск и периферическую «корону». Ропалии и щупальца сидят на особых студенистых цоколях. Немногочисленные представители отряда (Atolla, Periphylla) встречаются преимущественно на больших глубинах.
Большинство сцифомедуз относится к двум последним отрядам.
Отряд 4. Semaeostomeae. Характерно наличие дисковидно сплющенного зонтика, несущего по краю многочисленные щупальца. Сюда принадлежат Aurelia aurita (см. рис. 105, Б) и обыкновенная медуза наших северных морей—Cyanea capillata (см. рис. 105, А). Некоторые медузы, особенно Pelagia, способны сильно светиться в темноте.
Отряд 5. Rhizostomida (корнеротые медузы). Обычно это крупные медузы, наибо-ле,е многочисленные в тропических морях. В Черном море встречается представитель этого отряда Rhizostoma pulmo (рис. 109, Б).У корнеротых нет щупалец по краю зонтика. Функция захвата добычи целиком осуществляется ротовыми лопастями. Некоторые корнеротые медузы в странах Азии в соленом виде употребляются в пищу.
133
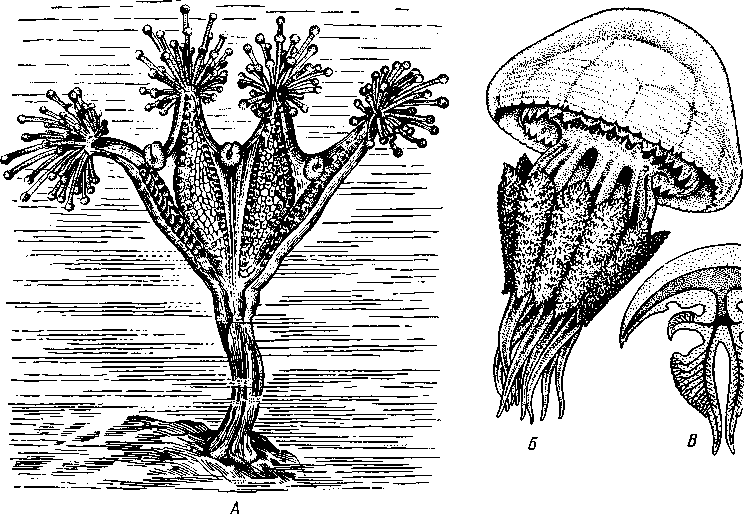
Рис. 109. Представители сцифомедуз. А — сидячая медуза Haliclystus (вид сбоку);
Б — корнеротая медуза Rhizostoma pulmo; В — схема продольного разреза корнеро-
той медузы (по Деляжу и Эреуару)
Мягкость тела и отсутствие скелета у медуз препятствуют хорошему сохранению их в ископаемом состоянии. Однако отдельные отпечатки попадаются, начиная с нижнего Кембрия. Наибольшее количество отпечатков медуз сохранилось в юрском сланце из Золеигофена.
КЛАСС III. КОРАЛЛОВЫЕ ПОЛИПЫ (ANTHOZOA)
Коралловые полипы бывают только полипоидной формы. Они совершенно не обнаруживают чередования поколений. Это морские животные, иногда одиночные, большей частью колониальные, причем колонии могут достигать значительной величины. Это наиболее крупный класс ки-шечнополостных, охватывающий 6000 видов.
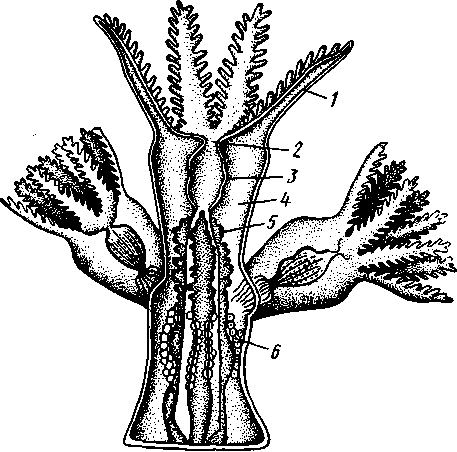
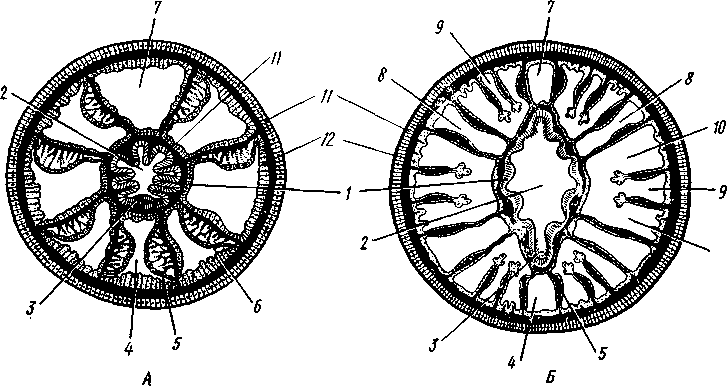
Строение и физиология. Коралловые полипы напоминают гидроидных, но устроены значительно сложнее. Тело отдельной особи имеет форму цилиндра (рис. ПО). Нижний конец одиночных полипов уплощен в подошву, служащую для прикрепления к субстрату, у колониальных форм он соединен со стволом или ветвью колонии. Ротовое отверстие помещается в центре противоположного конца тела. Вокруг рта располагается венчик полых щупалец, число которых у одних полипов (подкл. Octocorallia) равно восьми, в других же (подкл. Hexacorallia) представляет число, кратное шести.
Рот ведет сначала в длинную трубку — глотку, которая свешивается в гастральную полость. Глотка образуется впячиванием стенки ротового диска, вследствие чего выстлана внутри эктодермальным эпителием. Глоточная трубка сплющена в одном направлении, так что просвет глотки получает вид более или менее широкой щели. На одном или обо-
134
Рис. 110. Схема части колонии восьмилучевого коралла Alcyonium (по Л. и В. Шимкевич): ; — щупальца. 2 — ротовое отверстие. 3 — глотка, 4 — септа, 5 — мезентериальные нити, 6 — яйца
их концах этой щели расположены ротовые желобки — сифоноглифы (рис. 111), которые несут клетки с очень длинными ресничками. Находясь в непрерывном движении, реснички гонят воду внутрь гастральной полости полипа, тогда как на остальном участке глотки вода выводится из гастральной полости обратно наружу. Таким путем обеспечивается постоянная смена воды, имеющая важное значение для жизни полипа. В кишечную полость поступает свежая, богатая кислородом вода, а у полипов, питающихся мелкими планктонными организмами, и пища. Вода, отдавшая кислород тканям полипа и насыщенная углекислотой, выносится наружу вместе с непереваренными пищевыми остатками.
Собственно гастральная полость выстлана энтодермой и поделена на участки (камеры) особыми радиальными перегородками или септами. Число камер соответствует числу щупалец. Септы состоят из мезоглеи, одетой энтодермой. В верхней части полипа перегородки прирастают
 ю ю
Рис. 111. Поперечные разрезы через восьмилучевой (А) и шестилучевой (Б) коралловые полипы (Л — по Хиксон, Б — по Хайман):
1 — глотка, 2—полость глотки, 3 — сифоноглиф, 4 — вентральная направительная камера, 5 — септа, 6—мускульный валик септы, 7 — дорзальная направительная камера, 8 — внутренние камеры, расположенные между двумя септами первого порядка, 9 внутренние камеры, образующиеся между вторично возникающими септами, 10—промежуточные камеры, 11 — эктодерма,
12 — энтодерма. Мезоглея зачернена
135

одним краем к стенке тела, другим к глотке. В нижней части полипа (ниже глотки) они прикрепляются лишь к стенке тела, вследствие чего центральная часть гастральной полости — желудок — остается неразделенной. Число септ соответствует числу щупалец. Свободные края септ утолщены, они называются мезентериальными нитями (см. рис. ПО). Последние играют важную роль в пищеварении, так как несут железистые энтодермальные клетки, выделяющие пищеварительные ферменты. Гистологическое строение коралловых полипов усложняется по сравнению с гидроидными — вместо тонкой базальной мембраны у них толстый слой мезоглеи, кроме того, вместо эпителиально-мышечных клеток под эктодермой расположен слой обособившихся мускульных клеток (продольных и кольцевых). Произошла дифференциация мускульной ткани. Сильно развитые, энтодермального происхождения мускульные клетки имеются и в септах. Они образуют проходящие вдоль каждой септы (с одной из ее сторон) утолщения — мускульные валики (рис. 111). Подкожное нервное сплетение у коралловых полипов выражено гораздо сильнее, чем у гидроидных.
Рис. 112. Известковые иглы (спикулы) восьмилучевых кораллов
Рис. 113. Схема строения шестилуче-
вого кораллового полипа: образова
ние склеросепт (по Пфуртшеллеру):
/ — щупальца, 2 — ротовое отверстие. 3 — глотка, 4 — септы, 5—подошвенная пластинка, 6— чашечка, 7 — склеросепты, впячивающие мягкие ткани полипа (8)в гастральную полость
136
Только одиночные коралловые полипы, да и то не все, лишены скелетных образований. Напротив, у колониальных кораллов есть скелет, чаще всего состоящий из углекислой извести, реже (у части восьми-лучевых кораллов Octocorallia) из рогоподобного вещества. У восьми-лучевых кораллов известковый скелет залегает внутри мезоглеи и в простейшем случае состоит из разбросанных микроскопических известковых игл (рис. 112). Последние, как и у губок, формируются внутри особых клеток — склероблас-тов. У благородного коралла известковых игл (спикул) так много, что большинство их сливается в плотную массу, образуя твердый скелет. У шестилучевых кораллов (рис. 113) известковый скелет устроен иначе. У молодой особи сначала клетками эктодермы выделяется кнаружи подошвенная пластинка, а затем кругом тела полипа скелетная чашечка, или тека. Далее от теки внутрь тела врастают скелетные перегородки (склеросепты), впячивающие перед собой стенку полипа глубоко в его гастральную полость.
У колоний, состоящих из громадного числа особей, чашечки соседних полипов часто сливаются. Итак, у подкл. Octocorallia скелет внутренний (залегает внутри мезоглеи), у подкл. Hexacorallia по происхождению он внешний, так как лежит кнаружи от эктодермы, представляя продукт ее жизнедеятельности.
Склеросепты и настоящие мягкие перегородки имеют вполне определенное число и расположение, служащие важным систематическим признаком.
Восьмилучевые кораллы Octocorallia (см. рис. 111, Л) имеют 8 перегородок, делящих гастральную полость на 8 периферических камер; две из этих камер, приходящиеся против узких краев глоточной трубки, называются направительными. На септах в строго определенном порядке расположены мускульные валики. В результате в полость одной из направительных камер (условно называемую вентральной) обращены два мускульных валика. В другую направительную камеру (дорзальную) мускульные валики не вдаются.
У шестилучевых кораллов расположение септ сложнее (см. рис.'111, Б).Число расположенных попарно перегородок кратно шести, но по меньшей мере их 12. Перегородки возникают не все сразу. Сначала образуются шесть пар перегородок первого порядка, которыми гаст-ральная полость делится на 12 камер. Камеры, лежащие между двумя перегородками одной пары, называются внутренними, расположенные же между перегородками различных пар — промежуточными. Дальнейшие перегородки возникают парами, образуясь в пределах промежуточных камер.
Склеросепты развиваются всегда во внутренних камерах и не препятствуют закладке новых мягких септ, образующихся только в промежуточных камерах.
Физиологические отправления исследованы преимущественно у актиний— одиночных крупных коралловых полипов, лишенных скелета. Актинии в противоположность колониальным формам могут медленно ползать при помощи своей мускулистой подошвы. При раздражении актинии сильно сокращаются, втягивают щупальца и превращаются в небольшой твердый комок. Актинии очень чувствительны, особенно их щупальца.
Крупные актинии — большие хищницы, питаются раками, моллюсками и т. п. Добыча захватывается щупальцами, которые выпускают заряд стрекательных нитей, затем рот раскрывается, часть глотки выворачивается из него в виде широкого мешка и обхватывает пойманное животное.
Пищеварение актиний так же, как у гидр, представляет сочетание внутриклеточного с полостным. •
Размножение и развитие. Коралловые полипы размножаются бесполым и половым путем. Одиночные мягкие актинии иногда размножаются делением, у колониальных форм наблюдается почкование. Коралловые полипы, как правило, раздельнополы. Половые железы формируются в перегородках, между их энтодермой и мезоглеей. При половом размножении сперматозоиды прорывают эпителий септы мужских особей, выходят через рот наружу и через рот же проникают в женские особи, где и происходит оплодотворение яйца. Начальные стадии развития проходят в мезоглее септ. У многих актиний все развитие вплоть до образования полипа происходит в гастральной полости материнского организма. У некоторых Anthozoa оплодотворение наружное.
Развитие коралловых полипов идет сравнительно просто. Яйца испытывают полное равномерное дробление; сначала формируется бластула, потом мерцательная планула, которая плавает, а затем оседает перед-
137
ним концом на дно и, подобно плануле гидрозоев, превращается в молодого полипа.
Коралловые рифы и роль коралловых полипов в образовании земной коры. В тропических частях Атлантического, Индийского и Тихого океанов распространены рифообразующие (мадрепоровые) кораллы, отличающиеся иногда очень крупными' размерами (более 2 м в диаметре; ветви коралла Асгорога достигают высоты свыше 4 м). У них массивный известковый скелет, они образуют на мелководье густые поселения — коралловые рифы. Наиболее известны рифы побережья Австралии, Индонезии и островов Океании, многие из которых целиком слагаются из кораллового известняка. Дальше всего на север коралловые рифы заходят в Красном море. Самого «цветущего» состояния они достигают там, где зимняя температура воды не опускается ниже 20°С. Вместе с тем эти кораллы имеют ограниченное распределение по вертикали, опускаясь лишь до глубины 50 м. Это связано с тем, что в тканях рифообра-зующих мадрепоровых кораллов живут симбиотические одноклеточные водоросли — зооксантеллы, которые нуждаются в солнечном свете. Роль зооксантелл в жизнедеятельности мадрепоровых кораллов до конца еще не выяснена. Предположение о том, что кораллы способны переваривать эти водоросли, в последнее время не подтвердилось. По-видимому, зооксантеллы необходимы для нормального образования скелета кораллов. Рост скелета резко замедляется в темноте или у кораллов, искусственно лишенных зооксантелл. Все рифообразующие кораллы нуждаются в морской воде с нормальной океанической соленостью, т. е. содержащей около 3,5% солей. В связи с этим они никогда не поселяются в устьях
Рис. 114. Участок кораллового рифа во время отлива (по Гентшелю)
рек и других опресненных участках моря. Мадрепоровые кораллы нуждаются также в чистой прозрачной воде, достаточно насыщенной кислородом. Коралловый риф служит местом обитания и развития многих морских организмов (pile. 114). Здесь в большом количестве поселяются водоросли, моллюски, черви, ракообразные, иглокожие и представители других групп донных кишечнополостных. В зарослях коралловых рифов обитают многочисленные коралловые рыбки. Все эти животные и растения в совокупности образуют своеобразное сообщество, или биоценоз, кораллового рифа. Часть членов этого сообщества обладает мощным известковым скелетом и наряду с мадрепоровыми кораллами принимают участие в образовании рифа. Другие же находят здесь убежище и питаются за счет рифообразующих организ-мов- Животные кораллового биоценоза имеют причудливую
138
пеструю расцветку, которая помогает им укрываться на фоне ярко окрашенных колоний мадрепоровых кораллов.
Рифы распадаются на три разновидности: береговые, барьерные и атоллы. Береговые окаймляют берег суши, барьерный риф расположен параллельно берегу, но на некотором от него расстоянии. Особенно знаменит Большой Барьерный риф, тянущийся на протяжении 1400 км вдоль восточного берега Австралии.
Атолл — коралловое кольцо, выдающееся из моря на небольшую высоту, внутри кольца находится озеро морской воды, лагуна. На такой атолл приносятся водой или ветром семена разных растений (в том числе кокосовые орехи) и он превращается в цветущий островок.
Происхождение береговых рифов не требует объяснений. Труднее объяснить возникновение атоллов, так как они со всех сторон окружены большими глубинами, не подходящими для поселения кораллов.
Ч. Дарвин, наблюдавший коралловые рифы и острова во время путешествия на «Бигле», предполагал, что все разновидности рифов возникли из береговых вследствие постепенного опускания дна океана. Если на месте острова, окруженного береговым рифом, морское дно начнет медленно понижаться, то остров станет уходить под уровень воды и уменьшаться в размерах. Кораллы, его окружающие, по мере опускания рифа будут надстраивать его кверху, так как живут лишь на небольших глубинах, более же глубокие части рифа начнут отмирать. Таким образом, в то время как сам остров будет опускаться, риф, его окаймляющий, останется у поверхности воды, постепенно отделится от сократившегося в размерах острова и превратится в барьерный. При полном опускании острова на его месте остается мелководная лагуна, а риф превращается в атолл.
Другими учеными (Агассиц и др.) возникновение барьерных и атолловых рифов объясняется иначе. Они считают, что кораллы покрывают самые вершины находящихся под водой горных кряжей и пиков. Эти горы сначала выдавались над водой, но потом были настолько разрушены атмосферными влияниями, что ушли под уровень моря, а на месте самых высоких точек их образовались мелководные участки моря, очень удобные для поселения кораллов. Бурение атоллов показало, однако, что на несколько сотен метров в глубину их основание состоит из отмершего кораллового полипняка. Это подкрепляет не взгляды Агассица, а теорию, высказанную Дарвином.
Некоторые авторы (Меррей) принимают, что атоллы и барьерные рифы возникают вследствие растворяющего известь действия морской воды. Наружные, обращенные к открытому океану части рифа омываются водой, богатой солями извести, что способствует росту полипняков, внутренние же части рифа пользуются водой, которая обеднена известью, но зато обогащена углекислотой — продуктом дыхания коралловых полипов, способствующей растворению извести. Вследствие этого отмирающие внутренние части рифа постепенно растворяются морской водой, тогда как периферические продолжают надстраиваться. Однако в результате последующих гидрохимических исследований моря теория Меррея не получила подтверждения. Оказалось, что углекислота, содержащаяся в воде лагуны, так же, как и по внешнему краю рифа, в условиях высокой температуры тропиков известь не растворяет. Таким образом, ни одна из более новых теорий происхождения коралловых рифов и островов не опровергла основных положений Дарвина.
Что касается способа происхождения надводной части рифа, то все рифы сначала подводные, затем постепенно море обламывает краевые части рифа и обломки выкидывает на поверхность. Слой обломков рас-
139
тет, промежутки между крупными полипняками заполняются мелким обломочным материалом и, наконец, на месте бывшего подводного рифа получается слой слежавшейся известковой массы до 4 м и более высоты.
Мощная созидательная деятельность полипов объясняется отчасти их огромным количеством, отчасти легкостью процесса усвоения извести из морской воды в тропических водах.
Коралловые рифы существовали, начиная с древнейших геологических эпох, но состав животных-рифообразователей менялся. Рифы кайнозоя и мезозоя были построены из кораллов, сходных с современными. В палеозое строителями рифов были вымершие коралловые полипы из подклассов Rugosa и Tabulata. Кроме того, большое участие в создании рифов принимали Strornatoporoidea — вымершие представители Hydro-zoa, близкие к современному подотр. Hydrocorallia.
Классификация. К классу Anthozoa относятся два современных и три целиком вымерших подкласса.
ПОДКЛАСС I. ВОСЬМИЛУЧЕВЫЕ КОРАЛЛЫ (OCTOCORALLIA)
Представители подкласса имеют 8 щупалец и 8 септ, скелет Octoco-rallia развивается в мезоглее (рис. 115). Объединяет отряды: альционарии (Alcyonaria), горгонарии, или роговые кораллы (Gorgonaria), и морские перья (Pennatularia). К альционариям принадлежат наиболее просто устроенные коралловые полипы. Скелет их состоит из разбросанных в мезоглее спикул, никогда не сливающихся в осевой стержень. Колонии альционарии образуют густые заросли на небольших глубинах тропических морей, но встречаются также в умеренных и полярных водах. В северных и дальневосточных морях СССР наиболее обычна разветвленная
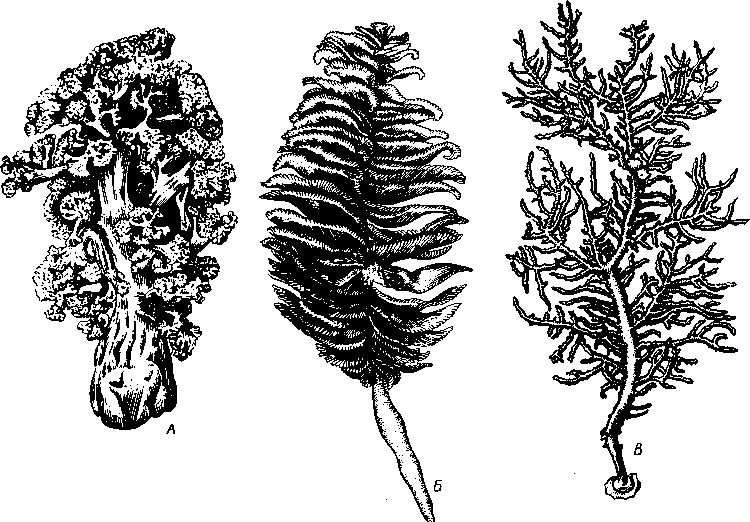
Рис. 115. Различные восьмилучевые кораллы. А — альционария Gersemia; Б — морское перо Pennatula; В — колония рогового коралла Leptogorgia
140
Gersemia. У Gorgon aria кроме спикул по оси древовидной колонии залегает роговой стержень, иногда частично или полностью обызвествлен-ный. Некоторые роговые кораллы замечательны тем, что их скелет содержит значительное количество иода. Этим, вероятно, объясняется применение их в средние века в качестве лекарственного средства. Особое значение имеет благородный коралл (Corallium rubrum); встречается на глубине свыше 50 м в Средиземном море. Известковый скелет благородного коралла, окрашенный в розовый или кир.пично-красный цвет, служит для изготовления украшений и мелких поделок. Изящные морские перья с правильным двухрядным расположением полипов на прямом стволе оранжевой, розовой или фиолетовой колонии относятся к отряду Pennatularia. Главный ствол такой колонии образован первичным полипом; в толще ствола нередко залегает роговой опорный стержень. Колонии морских перьев могут внедряться своим основанием в песчаный или илистый грунт. Некоторые из них обладают способностью к свечению.
ПОДКЛАСС II. ШЕСТИЛУЧЕВЫЕ КОРАЛЛЫ (HEXACORALLIA)
Шестилучевые кораллы делятся на пять отрядов, из которых наиболее обширны отряды актиний (Actiniaria) и мадрепоровых (Madrepora-ria). Актинии — одиночные полипы, способные медленно ползать при помощи подошвы. Это лишенные скелета крупные полипы (иногда свыше 60 см в диаметре), отличающиеся правильностью формы и красотой расцветки. Некоторые актинии (Sagarlia и др.) живут в симбиозе с раками-отшельниками, на раковинах которых они поселяются (рис. 116). При этом рак служит для актиний средством передвижения, тогда как близкое соседство актиний, вооруженных стрекательными капсулами, защищает рака от нападения хищников. Вырастая и переселяясь в более крупную раковину, рак пересаживает актиний клешнями с прежней раковины на новую. Мадрепоровые кораллы (Mad-reporaria) образуют колонии с массивным наружным извест- Рис' 116' Актиния на Раковине, занятой раком-
FJ .. отшельником (по Макарову)
ковым скелетом. Мадрепоро
вые кораллы — основные обра-
зователи коралловых рифов; известны, начиная с триаса. Кроме того, имеются отряды одиночных (Ceriantharia) и колониальных (Zoantharia и Antipatharia) полипов.
ПОДКЛАСС III. ЧЕТЫРЕХЛУЧЕВЫЕ КОРАЛЛЫ (TETRACORALLIA, ИЛИ RUGOSA)
Это вымершие палеозойские, большей частью одиночные (рис. 117, А),реже колониальные кораллы; чаще имеют вид расширенного к верхнему концу бокала или рога. В молодом возрасте Tetracorallia обнаруживают, судя по их скелетам, закладку
141
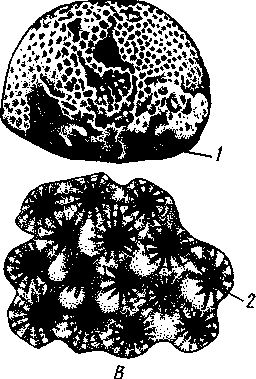


Рис. 117. Ископаемые кораллы. А—одиночный четырехлучевой коралл Omphiutna (из Деляжа); Б — колония кораллов Tabulata (из Друскина и Обручевой); В — колония Heliolitidae (из Байера): / — общий вид, 2 — участок колонии 6 первичных перегородок, что говорит в пользу их родства с подкл. Hexacorallia. Следующие склеросепты закладываются не во всех шести, а только в четырех секторах. Отсюда и название этих кораллов — четырехлучевые. Второе наименование — Rugosa — связано с характерными кольцевыми морщинами на наружной поверхности скелета. Четырехлучевые кораллы распространены с ордовика до пермской эпохи, но главного развития достигают в каменноугольную эпоху, когда за их счет образовывались значительные коралловые рифы, ПОДКЛАСС IV. ТАБУЛЯТЫ (TABULATA)Вымершие колониальные (рис. 117, Б),реже одиночные кораллы с полипами незначительных размеров. Число склеросепт обычно кратно 6, но, как правило, они недоразвиты или имеют вид расположенных рядами шипиков. Древнейшие табуляты известны с кембрия, но наиболее широкое распространение получили в ордовике, силуре и девоне, участвуя в образовании коралловых рифов. К началу мезозоя табуляты полностью вымерли. ПОДКЛАСС V. ГЕЛИОЛИТИДЫ (HELIOLITIDAE)Большая группа вымерших палеозойских кораллов, исключительно колониальных. Они характеризуются наличием 12 склеросепт. Гелиолитиды были широко распространены и поэтому имеют большое значение как руководящие формы при определении возраста палеозойских отложений. Филогения типа CoelenterataНизшими из кишечнополостных являются, без сомнения, гидрозои (Hydrozoa). Это доказывается тем, что у них тело представляет собой двухслойный мешок, оба слоя которого соприкасаются по краям рта. Исходной формой гидрозоев следует, по-видимому, считать морских гидроидных полипов, которые уже вторично дали начало медузам, отличающимся гораздо более сложной организацией. Появление медуз имело прогрессивное значение, так как они играли большую роль в расселении вида. Однако у современных Hydrozoa в ряде случаев произошла вторичная утрата медузоидного поколения. Пресноводные Hydrida не могут рассматриваться как примитивные в филогенетическом смысле формы, так как особенности их строения и 142
жизненного цикла, по-видимому, вторично изменены в связи с переходом к пресноводному образу жизни.
Среди гидрозоев сифонофоры представляют, вероятно, группу более позднего происхождения, у -которой произошла значительная и разно-направленная специализация отдельных особей колоний, в результате чего получились полиморфные колонии.
В процессе эволюции Scyphozoa полипоидная стадия (сцифистома) усложнилась незначительно, тогда как сцифомедузы достигли гораздо более высокой степени сложности, что связано с их свободноплаваюшим образом жизни. Эволюция Anthozoa, жизненный цикл которых проходил без метагенеза, шла в направлении приспособления к сидячему образу жизни, для которого очень характерно размножение почкованием и образование колоний. Этот класс кишечнополостных проделал значительную прогрессивную эволюцию в направлении усложнения строения, так как пищеварительный аппарат Anthozoa дифференцируется уже на две части: эктодермальную глотку и энтодермальную среднюю кишку. Кроме того, у Anthozoa и Scyphozoa пищеварительная система испытывает усложнение и в смысле образования стенками желудка перегородок, увеличивающих поверхность всасывания пиши. Ввиду сходства плана строения этих классов с таковым гидрозоев можно считать их генетически связанными с этим низшим классом.
ТИП ГРЕБНЕВИКИ (CTENOPHORA)
Гребневики — морские свободноплавающие, реже ползающие или сидячие радиально-симметричные (двухлучевые) животные. Тело, как и у кишечнополостных, состоит из двух слоев клеток — эктодермы и энтодермы, между которыми имеется толстый слой мезоглеи. Характерная черта типа — наличие клейких клеток. Движение осуществляется работой видоизмененных ресничек.
Тип Ctenophora, объединяющий около 90 видов, имеет единственный класс, носящий то же название.
КЛАСС ГРЕБНЕВИКИ (CTENOPHORA)
Строение и физиология. Тело чаще всего мешковидно, причем на одном конце мешка помещается рот; этот полюс тела называется ротовым, оральным, противоположный же — аборальным (рис. 118). Главная ось тела проходит через оба полюса; через главную ось можно провести две различные плоскости симметрии, т. е. тело гребневиков построено по радиальному, а именно по двухлучевому типу. Однако следует заметить, что многие органы (ряды гребных пластинок, радиальные каналы, половые железы) расположены вокруг главной оси в числе 8. Таким образом, общее строение тела гребневиков обнаруживает сочетание двух типов симметрии: двухлучевого и восьмилучевого.
Рот ведет в эктодермальную глотку, имеющую вид сильно сплющенной в одном направлении трубки; кверху глотка переходите энтодермаль-ный желудок — мешок, сплющенный в направлении, перпендикулярном к плоскости сплющивания глотки. Плоскость, по которой сплющена глотка, называется глоточной, а та, в которой происходит сплющивание желудка, — щупальцевой, так как в ней же лежит пара щупалец (см. ниже); эти плоскости и являются плоскостями симметрии (рис. 118). От желудка отходят каналы гастроваскулярной системы. Один канал направлен к полюсу, противоположному рту, и непосредственно под аборальным полюсом распадается на 4 короткие ветви: две слепые и две,
143
заканчивающиеся узкими отверстиями. Другие два канала отходят от желудка в щупальцевой плоскости и в направлении, перпендикулярном
8
главной оси тела, — это каналы первого порядка; каждый из них на пути к краю тела дважды дихотомически ветвится, давая 2 канала второго и потом 4 канала третьего порядка. Получившиеся таким образом 8 каналов третьего порядка впадают в соответственное число меридиональных каналов, идущих параллельно поверхности тела от або-рального полюса к оральному; оба конца меридиональных каналов замкнуты слепо. От желудка отходят еще два канала, идущие к оральному полюсу по сторонам глотки. Соответственно меридиональным каналам на
Рис. 118. Схема строения гребневика, перерезанного поверхности тела прохо-
поперек (по Гертвигу): дят 8 ребер, или валиков,
/ — ротовое отверстие, 2 — глотка, 3 — желудок, 4 — мери- ня КОТОПЫХ СИЯЯТ Г ЛЯ RHhTP
диональные каналы, 5 - ряды гребных пластинок, 6 - щу- "<* КО ШрЫХ СИДЯТ ГЛаВНЫе
пальца, 7 — аборальный орган, 8 — влагалища щупалец, ОрГЭНЫ ДВИЖ6НИЯ Гребне-
9 - каналы, идущие к оральному полюсу BHKQB _ гребные пластин_
ки. Расходясь от абораль-
ного полюса, ребра доходят почти до ротового. Каждое ребро усажено рядом поперечно поставленных четырехугольных небольших пластинок: они прозрачны, тонки и на конце расщеплены наподобие гребешка. Пластинки бьют по воде в одном направлении, действуя наподобие множества маленьких весел, и передвигают животное ротовым полюсом вперед. Гистологическое строение пластинок показывает, что каждая состоит из ряда слившихся друг с другом крупных ресничек.
От рядов пластинок берут начало бороздки, покрытые ресничками (мерцательные шнуры), которые сливаются затем попарно и в числе четырех направляются к -аборальному полюсу.
Таким образом, гребневики в течение всей жизни движутся при помощи измененных ресничек, тогда как у кишечнополостных этот способ движения имеется лишь у личинок, во взрослом же состоянии заменяется мускульным движением.
Большая часть гребневиков снабжена парой щупалец, служащих для захвата добычи. Щупальца имеют вид длинных обычно разветвленных арканчиков и отходят от боков тела в щупальцевой плоскости, одно против другого. Они могут втягиваться в особые ямки — щупальцевые влагалища. Захват пищи осуществляется при помощи особых клейких клеток эпителия щупалец. Наружная поверхность клейкой клетки имеет вид полушария, покрытого каплями липкого секрета. От основания полушария внутрь щупальца отходит спирально закрученная нить, прикрепленная другим концом к пучку продольных мышц, проходящих по оси щупальца и его ветвей (рис. 119). Мелкие морские орга-
!44
низмы при соприкосновении со щупальцами приклеиваются к клейким клеткам. Если добыча пытается вырваться, то клейкие клетки отходят
Рис. 119. Строение клейкой клетки
гребневика (из Наумова): / — полушария с каплями липкого секрета, 2 — спиральная нить, 3 — мускульный тяж щупальца
Рис. 120. Схема строения аборального органа (по Кестнеру):
/ — колпачок (колокол) из слившихся ресничек, 2 — дужки, 3—статолит, 4 — мерцательные бороздки
от поверхности щупальца, но остаются соединенными с ним посредством эластичной спиральной нити, которая, как пружина, амортизирует рывки захваченного животного. Пойманная щупальцами добыча захватывается подвижными краями широкого рта. Нервная система гребневиков состоит из поверхностного сплетения нервных клеток, которые под рядами гребных пластинок и мерцательными шнурами образуют более плотные тяжи, идущие к аборальному полюсу. Здесь расположен своеобразный орган чувств, получивший название аборального органа (рис. 120). Основу его составляет утолщение эктодермы в виде подушечки, прикрытой прозрачным колпаком, образующимся из венчика слипшихся между собой длинных ресничек. Внутри колпака над поверхностью подушечки расположены 4 крючковидные дужки, образованные длинными слившимися жгутиками, отходящими от рецепторных клеток, связанных с аборальным органом. На дужках подвешен статолит, состоящий из склеенных между собой зерен фосфорнокислого кальция. Основания дужек сообщаются при помощи мерцательных жгутов с рядами гребных пластинок. Хотя весь этот аппарат трактуется как орган равновесия, он служит также и для регуляции движения. У животного с удаленным аборальным органом движение гребных пластинок нарушается. Мезоглея гребневиков хорошо развита, прозрачна и водяниста, как у медуз. Гребневики не отличаются яркостью окраски, иногда имеют слегка розовый оттенок. Бьющие, прозрачные гребные пластинки переливаются на общем фоне тела всеми цветами радуги. Размножение и развитие. Гребневики гермафродиты, причем половые клетки их дифференцируются в энтодерме. По бокам каждого меридионального канала пищеварительной системы залегает с одной стороны колбасовидный яичник, с другой — такой же семенник. Зрелые половые клетки выходят посредством прорыва разделяющей их стенки в просвет меридиональных каналов, а оттуда через рот наружу, где и происходит оплодотворение. Жизненный цикл гребневиков простой, без метаморфоза. Дробление 145
полное, но не равномерное, один полюс зародыша состоит из быстро
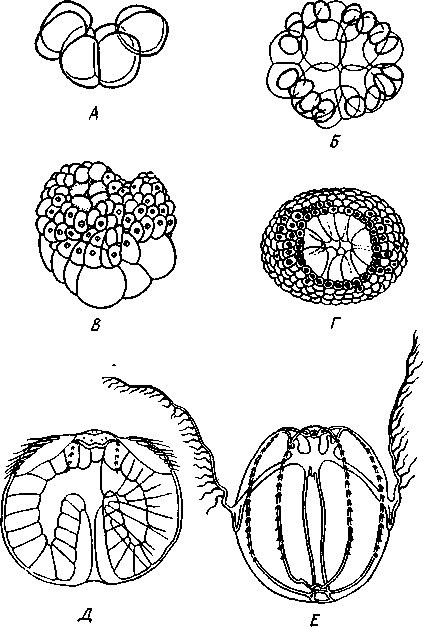
Рис. 121. Эмбриональное развитие гребневика Во-linopsis (из Байера). А — ранняя стадия дробле-вия; Б — образование микромеров; В, Г — стадии гасгруляции; Д — продольный разрез молодой личинки; Е — сформированная личинка
дробящихся мелких микромеров, другой — из медленно делящихся макромеров (рис. 121). Микромеры дают эктодерму, макромеры — энтодерму. Микромеры обрастают книзу массу макромеров, которая к тому времени обнаруживает на нижнем полюсе впячивание — первичный рот и зачаток гаст-ральной полости. Получается гаструла. По краям бла-стопора эктодерма впячивается и образует глотку. В глубине гастральной полости часть клеток энтодермы уходит в промежуток между эктодермой и энтодермой (т. е. в остаток полости бластулы) и дает там крестообразную группу клеток. Из этих клеток впоследствии формируются клеточные элементы мезоглеи и мускульные оси щупалец. Интересно, что здесь мы впервые видим во время развития обособленные зачатки специального третьего клеточного зародышевого пласта — мезодермы. Этим слабым зачатком является вышеупомянутая крестообразная клеточная пластинка.
Классификация. Ctenophora делятся на два подкласса: Tentaculata и Atentaculata. Первые в течение всей жизни или только на ранних стадиях развития имеют щупальца, вторые лишены щупалец на всех стадиях. Большинство видов относится к щупальцевым (Tentaculata). Некоторые из них имеют сильно измененную форму тела, которое вытягивается в поперечном направлении (в глоточной плоскости) в ленту до 1,5 м длины; таков «Венерин пояс» — Cestus veneris (рис. 122,5). Большой интерес представляют Platyctenidea. Эти гребневики (рис. 122, В) сплющены по направлению главной оси и не только плавают, но могут и ползать по субстрату на уплощенной ротовой стороне. При переходе от плавающего к ползающему образу жизни организация гребневиков претерпевает глубокие изменения. Исключение среди них представляет арктическая форма Tjalfielta (рис. 122,Г), которая ведет сидячий образ жизни. Молодь ее имеет вид типичных гребневиков. Молодое животное садится ртом на субстрат, средняя часть рта зарастает, а его концы вытягиваются кверху в трубки, служащие для захвата пищи. К бесщупальцевым гребневикам относится обычный в северных и дальневосточных морях Вегое сисщтз (рис. 122, Д). Питается он другими гребневиками, заглатывая их широко раскрывающимся ртом.
146
Филогения типа Ctenophora. Гребневики обладают некоторыми при-митивными чертами, например движением при помощи измененных
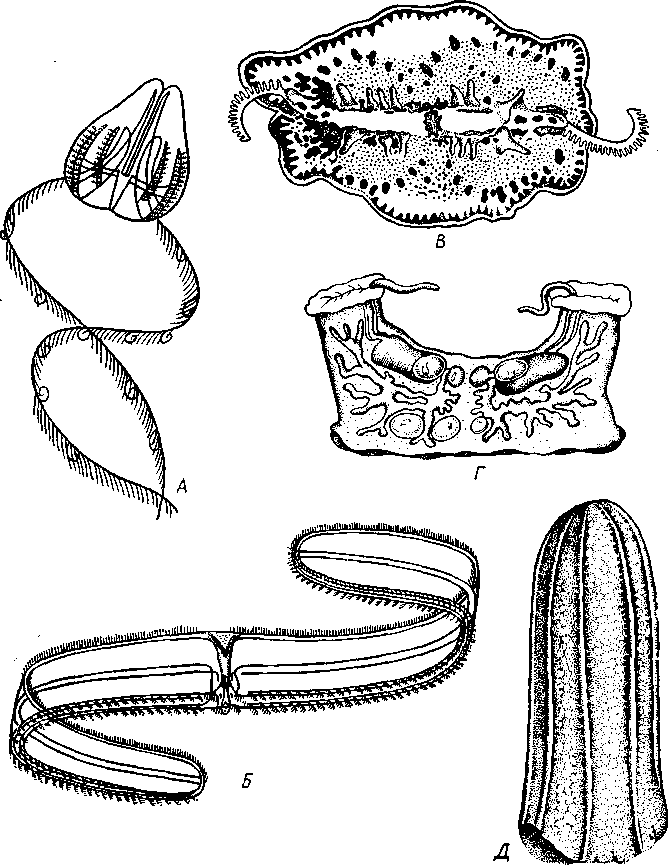
Рис. 122. Различные представители гребневиков. А — Г — щупальцевые гребневики: А — Cydippe (по Хуну): Б — «Венерин пояс» (Cestus veneris) (по Мейеру); В — ползающий гребневик Coeloplana (по Кюкенталю); Г — сидячий гребневик Tjalfiella (из Догеля); Д — бесщупальцевый гребневик Beroe cucumis (по Наумову)
ресничек. Развитие гребневиков обнаруживает следы закладки мезодермы— этим они приближаются к червям. Возможно, что гребневики отделились от общего с Coelenterata ствола развития до того, как последние стали сидячими животными,
|
|
|
 Скачать 14.77 Mb.
Скачать 14.77 Mb.