А-Догель - Зоология беспозвоночных_2. Тип губки (spongia, или porifera)
 Скачать 14.77 Mb. Скачать 14.77 Mb.
|

|
Патогенное значение моногеней. Моногенеи, паразитирующие на рыбах, нередко становятся причиной их заболеваний и даже гибели, особенно в условиях прудовых рыбных хозяйств. Наиболее важное практическое значение имеют многие представители сем. Dactylogyridae, живущие на жабрах пресноводных рыб. Так, Dactylogyrus, например, нападает на молодь карпов в таком числе, что на одной рыбешке встречается до 500 этих мелких (1—3 мм) паразитов. Они питаются слизью, эпителием или (реже) кровью хозяина, вызывая иногда массовую гибель рыб. К числу опасных паразитов относятся и представители семейства Gyrodactylidae. Гибель рыб в результате заражения моногенеями иногда происходит в природных условиях. Так, вид Nitzchia sturionis, паразитирующий на Хозяин не размножается Хозяин размножается 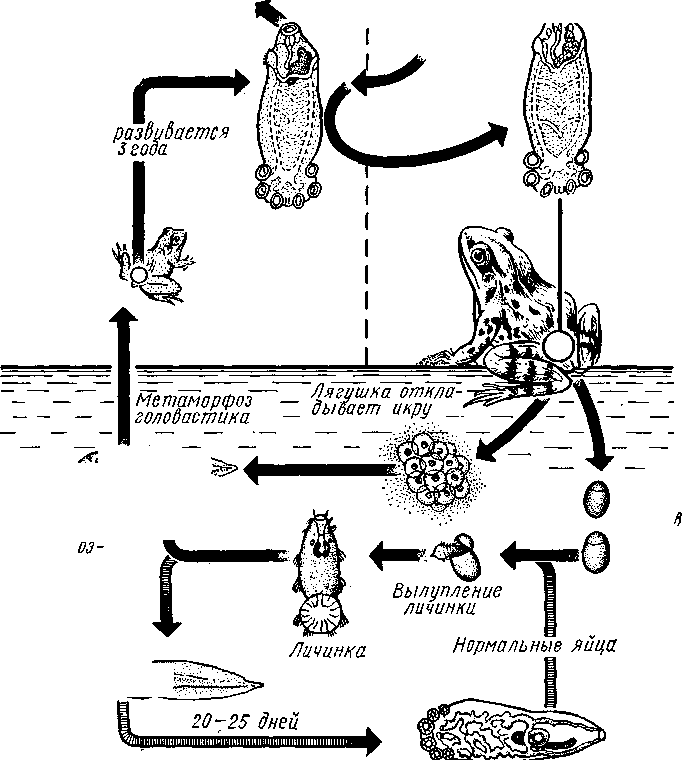 I I Половые гормоны хозяина стимулируют созревание Неполовозрелые полистомы Личинка Пол истома достигает л олово-зрелости, когда хозяин попадает 6 о од у Дичинка переходит д моче -бой пузырь jv^^s^iiK?4- ПрИкр'еТГ "^JtesasS ля юте я на -^ нппмп/и.нпр внутренние • ЗЖ/Р жМры fo/iee - поОеОение старших " растоо Яйца выделяются . воду Личинка прикрепляется к наружным кадрам молодо- €*£?^&?g. •го голоВас- •••••••••"**•• тика „ Жаберная форма " полистомы откладыоает яйца Рис. 152. Ход жизненного цикла лягушачьей многоустки Polystoma integerrimum (ВО жабрах осетровых рыб Волги и случайно завезенный при акклиматизации севрюги в Арал, перешел там на местных осетровых (шип) и вызвал среди них массовую гибель. Классификация моногеней основывается на особенностях строения их прикрепительного аппарата. В настоящее время Monogenoidea разбиваются но этому признаку на два подкласса, насчитывающие в общей сложности 9 отрядов. ПОДКЛАСС I. НИЗШИЕ МОНОГЕНЕИ (POLYONCHOINEA) К подклассу относятся моногеней, прикрепительный диск которых вооружен сильно развитыми крючьями, несет присоски или же может целиком превращаться в одну мощную присоску. Личинки (см. рис. 150) с глазами, несут на прикрепительном диске 14—16 мелких краевых крючочков. В основном—паразиты морских и пресноводных рыб. Это названные выше семейства Dactylogyridae, Gyrodactylidae и др. Представители семейства Polystomatidae паразитируют на амфибиях и рептилиях. Многие виды, например лягушачья многоустка (см. рис. 148), перешли к паразитированию во внутренних органах хозяина, т. е. стали эндопаразитами (с. 17S). Как правило, это крупные формы, характеризующиеся наличием разветвленного кишечника и множественных семенников (с. 177). Прикрепительный диск их обычно несет 6 мускулистых присосок. ПОДКЛАСС II. ВЫСШИЕ МОНОГЕНЕИ (OLYGONCHOINEA) Характерной чертой моногеней этого подкласса, паразитирующих только на рыбах, является наличие специализированных органов прикрепления — клапанов, действующих по принципу капкана (см. рис. 151). С появлением клапанов крючья утрачивают значение главных органов прикрепления и у взрослых червей могут иногда отсутствовать. У личинок обычно имеется 10 краевых крючочков. К этому подклассу относится рассмотренный выше Diplozoon paradoxum (см. рис. 151). КЛАСС IV, ЛЕНТОЧНЫЕ ЧЕРВИ (CESTODA) Ленточные черви — плоские черви, на которых паразитический образ жизни сказался значительно сильнее, чем на сосальщиках и моноге-неях. В половозрелом состоянии цестоды встречаются в кишечнике позвоночных животных; молодые стадии цестод живут в полости тела и внутри различных органов как беспозвоночных, так и позвоночных. Число видов превышает 3000. Строение и физиология взрослых ленточных червей. Тело обыкновенно бывает сильно вытянутым в длину, лентовидным и в большинстве случаев поделено па значительное число члеников, или проглоттид (рис. 153). Редко тело бывает цельным, нерасчлененным. Передний конец образует небольшую головку, или сколекс, за которым следует нерасчлененная шейка, а за ней идут проглоттиды. Головка несет органы прикрепления, построенные по типу присосок или по типу крючков (рис. 154). Присоски имеются всегда, тогда как крючья являются менее постоянным их дополнением. Присоски устроены, в общем, так же, как у сосальщиков, и чаще всего имеются в числе четырех по краю переднего конца головки. Реже вместо типичных присосок головка снабжена двумя вытянутыми в продольном направлении щелевидными присасывательными ямками. Крючья помещаются или прямо на поверхности головки, или на особом переднем выпячивании ее — хоботке, образуя один или несколько венчиков. Хоботок втяжной. В редких случаях (Tetrarhynchus) имеется 4 хоботка, длинных, усаженных многочисленными крючками и способных глубоко вворачиваться в особые влагалища. Проглоттиды имеют обычно четырехугольную форму, причем число их варьирует от 3 шт. до нескольких тысяч. Передние членики — самые 181 маленькие, по направлению же кзади размеры их постепенно возрастают. Во все время жизни червя происходит рост и увеличение числа члеников. Рост идет в области шейки: она удлиняется, и от заднего конца ее отшнуровываются все новые членики. Таким образом, самые молодые членики занимают переднюю часть тела; чем далее кзади расположен членик, тем он старее. Все тело называют цепочкой или стробилой (последнее за сходство с процессом стробиляции сцифистом сцифоидных медуз). Величина взрослых ленточных червей колеблется между 1 мм и 10м. Окраска тела однообразная — белая или желтоватая, характерная для многих внутренностных паразитов. Рис. 153. Общий вид стробилы бычьего солитера (по Холодковскому) Цестоды обладают типичным кожно-мускульным мешком. Их покровы очень сходны с таковыми у трематод и моногеней и построены по тому же типу, что и погруженный эпителий турбеллярий. Тегумент це-стод слагается из безъядерного цитоплазматического слоя, при помощи тонких тяжей соединенного с погруженными участками цитоплазмы, несущими ядра. Отличительная особенность покровов цестод состоит в  182 том, что на поверхности наружного цитоплазматического слоя имеется бесчисленное множество волосковидных выростов (микротрихий), по-видимому, играющих роль в процессе питания (рис. 155). Рассмотреть их можно только с помощью электронного микроскопа. Их ультраструкту- 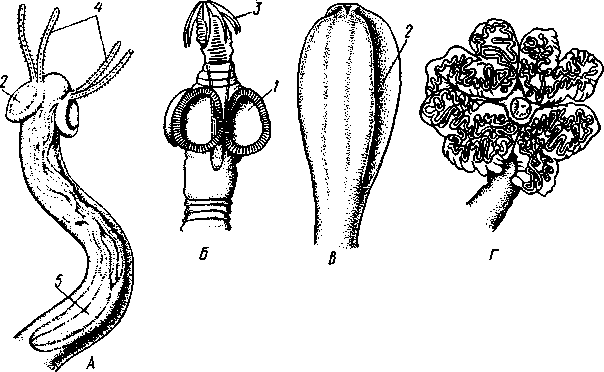 Рис. 154. Типы строения сколексов цестод (из разных авторов). А — Tetrarhynchus (Trypanorhyncha); Б — Hymenolepis (Cyclophyllidea); В — Diphyllobothrium (Pseudophyllidea); Г — Phyllobothrium (Tet- raphyllidea): / — присоски, 2 — присасывательные ямки, 3 — крючья, 4 — хоботки, вооруженные крючьями. 5 — влагалища, в которые втягиваются хоботки ра принципиально отличается от таковой микроворсинок, характерных для спороцист трематод. Непосредственно под базальной мембраной располагается наружный слой кольцевых и внутренний — продольных мускульных волокон. Часто к этим двум слоям присоединяется еще третий более глубокий слой кольцевых мышечных волокон (рис. 156). Кроме того, имеется система спинно-брюшных мышечных пучков, пронизывающих паренхиму. В последней у большинства цестод разбросаны микроскопические округлые конкреции углекислой извести — так называемые известковые тельца. Их происхождение и функция до конца не выяснены, но имеется предположение, что известковые тельца возникают как экскреты и выполняют роль своеобразной буферной системы, предохраняющей ленточных червей от вредного влияния кислой среды (например, при миграции личиночных стадий через желудок хозяина). В паренхиме ленточных червей откладывается значительное количество гликогена, в результате анаэробного расщепления которого це-стоды (подобно трематодам) получают энергию, необходимую для жизнедеятельности. Наиболее характерный признак ленточных червей заключается в полном отсутствии у них пищеварительной системы — факт, объясняющийся, вероятно, их образом жизни. Для цестод, обитающих в просвете кишечника другого животного (хозяина), нет необходимости самостоятельно захватывать и переваривать пищу. Последняя переводится в растворенное и доступное для усвоения состояние пищеварительными ферментами самого хозяина. Эту уже в значительной мере переваренную пищу паразиты воспринимают всей поверхностью тела. По-видимому, 183 особенности строения цитоплазматической пластинки тегумента, несущей микротрихии, способствуют осуществлению этого процесса. Важно отметить, что характер диеты хозяина, в особенности содержание в ней углеводов, влияет на состояние, интенсивность роста и развитие ленточных червей. Нервная система, как и вообще у паразитов, у цестод развита слабо. Органы чувств представлены разбросанными по поверхности тела чувствительными клетками, наибольшее количество их концентрируется на сколексе. Центральная нервная система состоит из парного мозгового узла, лежащего в головке и посылающего от себя назад несколько пар нервных стволов, соединенных поперечными перемычками (ортогон). Два ствола, расположенных по бокам тела, развиты сильнее остальных (рис. 157). От стволов отходят тонкие веточки, образующие под кожей довольно густое нервное сплетение. 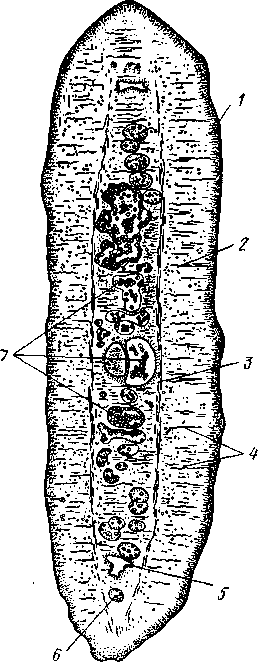 Выделительная система — протоне-фридиального типа (рис. 157, 158). По бокам вдоль всего тела, непосредственно кнутри от нервных стволов идут два главных выделительных канала. Они начинаются на заднем конце тела, затем на-  Рис. 155. Схема строения покровов цестод по данным электронной микроскопии (по Бегину): / — волосовидные выросты, 2 — наружный слой цитоплазмы с митохондриями (3)и различными включениями, 4 — цитоплазматиче-ские тяжи, соединяющие наружный слой цитоплазмы с погруженной частью тегумспта, 5— погруженные клеточные тела тегумента с ядрами (6), 7—продольные мышцы, 8 — кольцевые мышцы, 9 — базальная мембрана (поперечный срез) Рис. 156. Поперечный разрез через членик бычьего солитера Taeniarhynchus sagi-natus (по Стрелкову): / — кожно-мускульный мешок, 2—паренхима, 3 — слой внутренних кольцевых мышц, 4 — дорзовентральные мышцы, 5 — боковой сосуд выделительной системы, в — боковой продольный нервный ствол, 7 — перерезанные части половой системы 184 правляются кпереди, достигают головки, заворачиваются назад и вновь доходят до заднего конца, заканчиваясь общим выделительным отверстием. Вследствие этого часто получается впечатление, что у цестод имеются 4 продольных канала, тогда как их всего 2, но петлеобразно изогнутых у переднего конца тела. При соединении на заднем конце тела оба канала нередко образуют небольшой общий сократимый мочевой пузырь. У членистых цестод боковые каналы как в головке, так и у заднего края члеников соединяются при помощи поперечных перемычек; выделительная система получает вид лестницы. Когда последний членик цепи отвалится (вследствие периодического отрывания задних кусков стробилы), нового мочевого пузыря уже не образуется, и каждый боковой канал открывается теперь наружу особым отверстием. Многочисленные веточки каналов пронизывают паренхиму и на концах своих замыкаются звездчатыми клетками с мерцательным пламенем. Половая система (рис. 159, 160) ленточных червей гермафродитна и, в общем, напоминает таковую сосальщиков. Лишь у некоторых из нерасчлененных цестод (Caryophylla-eus) половой аппарат одиночен. У других, например у ремнецов, имеется продольный ряд половых аппаратов, тогда как у членистых цестод в каждой проглоттиде развивается своя половая система. Отдельные части полового аппарата у разных ленточных червей довольно сильно варьируют, так что мы для конкретности возьмем 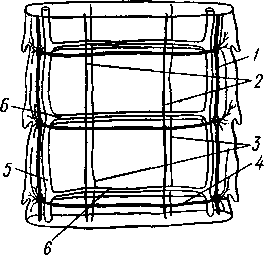 Рис. 157. Часть стробилы солитера с нервными стволами и выделительными каналами (по Фурману) : / — главный боковой нервный ствол, 2 •— спинные нервные стволы, 3 — брюшные нервные стволы, 4 — нервнее кольцо, 5 — продольный боковой канал выделительной системы, 6 — поперечная перемычка между продольными выделительными каналами Рис. 158. Выделительная система солитера (по Шим- кевичу): / — сколекс, 2 — проглоттиды, 3 — продольные боковые каналы выделительной системы, 4 — поперечные перемычки между продольными выделительными каналами, 5 — разветвления выделительных каналов (в двух последних члениках показаны мельчайшие ответвления, заканчивающиеся звездчатыми клетками) 185 один частный случай, например бычьего, или невооруженного, солитера (Taeniarhynchus saginatus) (рис. 159). В молодых передних члениках стробилы половые органы еще не развиты и начинаются приблизительно лишь с 200 членика. В следующих члениках с вполне развитой половой системой мужской отдел последней состоит из многочисленных семенников, разбросанных в паренхиме. Тонкие семявыносящие протоки семенников соединяются вместе и образуют общий семяпровод. Последний направляется к одной из узких боковых граней тела и там пронизывает собой совокупительный орган, имеющий вид мускулистой трубки, которая своим концом вдается в глубокую ямку на боковой грани тела — половую клоаку. Женский отдел системы состоит из ветвистого яичника, проток которого, яйцевод, впадает в оотип, как у трематод. В оотип же поступает и содержимое непарного желточника — сетевидной железы, прилегающей к задней стенке членика. Кроме того, от оотипа отходят два канала. Один, влагалище, тянется рядом с семяпроводом и открывается подле него в половую клоаку. Другой, более широкий, направляется от оотипа вперед по срединной линии членика и заканчивается слепо, это — матка. Яйцеклетки поступают в оотип, куда проникают и спермин через влагалище. В оотипе яйца оплодотворяются, окружаются скорлупой и переводятся в матку, где проходят первую часть своего развития. У цеп-ней вследствие отсутствия выводного отверстия яйца остаются в матке долго и выходят наружу лишь при разрыве стенок членика. Яйца настолько переполняют матку, что последняя сильно разрастается, дает от своего главного ствола много боковых ветвей в обе стороны и занимает значительную часть членика (см. рис. 162). К этому времени все остальные части половой системы заканчивают свою функцию и подвер- 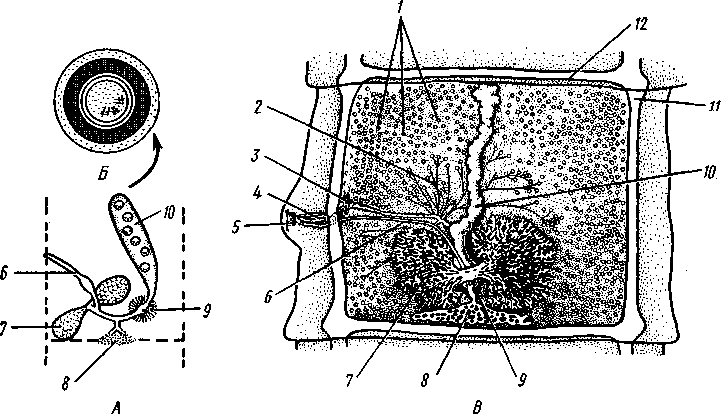 Рис. 159. Строение половой системы бычьего солитера Taeniarhynchus saginatus (отр. Cyclophyllidea). A — схема строения женской половой системы; Б — яйцо с заключенной внутри онкосферой (по Смиту); В — гермафродитный членик бычьего солитера (по Полянскому): / — семенники, 2 — семявыносящие протоки, 3 — семяпровод, А — совокупительный орган, 5 — половая клоака, 6 — влагалище, 7 — яичник, 8 — желточник, 9 — оотип, 10 — матка, // — продольный выделительный канал, 12—поперечная перемычка, соединяющая продольные выделительные каналы. Матка слепо замкнута. Яйца лишены крышечки, развиваются в матке. Свободного корацидия нет 186 гаются большей или меньшей атрофии. Членики, в которых осталась лишь сильно разветвленная и набитая яйцами матка, называют «зрелыми». Зрелые членики занимают задний конец цепочки и периодически отрываются целыми группами. Наиболее крупные из цестод образуют в течение своей долгой жизни поистине колоссальное количество яиц. Так, например, паразитирующий в кишечнике человека невооруженный це-пень (Taeniarhynchus saginatus) в среднем живет 18—20 лет и за год продуцирует до 600 млн. яиц, следовательно, за всю свою жизнь цепень производит около 11 млрд. яиц. У других цестод может быть ряд разнообразных отступлений. Так, матка, слепо замкнутая у цепня, нередко (например, у Diphyllobothrium и др.) открывается наружу на одной из плоских сторон членика (рис. 160). У таких видов яйца по мере заполнения ими матки выходят из нее в кишечник животного-хозяина. Мужское отверстие и отверстие влагалища могут при этом смещаться на одну из плоских сторон членика. Любопытным изменением является наблюдаемое у некоторых форм частичное или полное удвоение полового аппарата в каждом членике (например, у тыквенного цепня — Dipylidium caninum). Размножение и развитие. Оплодотворение у ленточных червей происходит как перекрестно, так и посред- 2 ю t9 g 1 ством самооплодотворения, причем совокупительный орган одного чле- 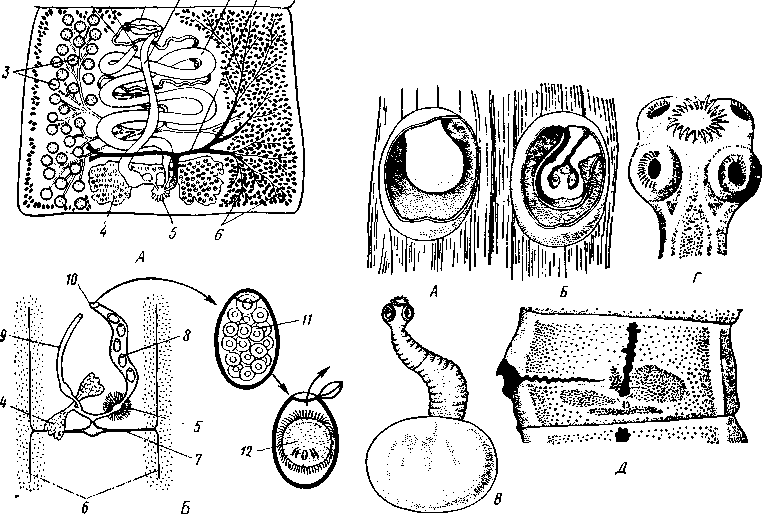 Рис. 160. Строение полового аппарата широкого лентеца Diphyllobothrium la-tutn (отр. Pseudophyllidea). A — герма-фродитный членик (из Шульца и Гвоздева) ; 5 — схема строения женской половой системы (по Смиту): / — совокупительный орган, 2— семяпровод, 3 — семенники, 4—яичник, 5 — оотип. б — желточники, 7— желточный проток, в—матка, 9 — влагалище, 10 — отверстие матки. Сложные яйца (//), снабженные крышечкой, выходят в воду, где в них развивается свободная личинка — корацидий (12) Рис. 161. Стадии развития свиного солитера Taenia solium (по Пфуртшеллеру). А, Б — финны в свином мясе; В — вывернутая финна; Г — сколекс взрослого червя; Д — гер-мафродитный членик 187 ника вводится во влагалище другого или даже, изгибаясь, во влагалище того же самого членика. Жизненный цикл цестод мы разберем сначала на частном случае свиного солитера — Taenia solium, паразитирующего в половозрелом состоянии в кишечнике человека (рис. 162). Яйца выходят наружу с испражнениями или посредством разрыва стенок члеников, или вместе с члениками, группы которых периодически отрываются от стробилы. Для дальнейшего развития яйца, как у сосальщиков, должны попасть во вполне определенного промежуточного хозяина, которым для Т. solium служит свинья. Заражение свиней происходит вследствие их нечистоплотности, так как они охотно роются в отбросах, нечистотах и т. п. У яиц, попавших в кишечник свиньи, скорлупа разрушается, и из яйца выходит личинка — онкосфера, или шестикрючный зародыш — маленький многоклеточный шарик, снабженный шестью хитиноидными крючками. Крючочки эти не соответствуют тем крючкам, которые у многих цестод образуются на сколексе, и отбрасываются при дальнейшем развитии. При помощи крючков онкосфера вбуравливается в стенки желудка или кишки, попадает в лимфатические или кровеносные сосуды и 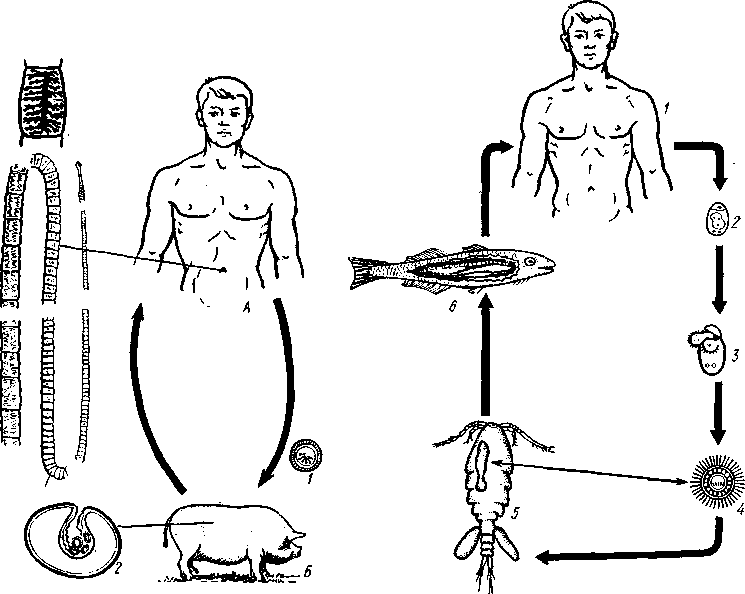 Рис. 162. Жизненный цикл свиного" солитера. А — окончательный хозяин (человек), в котором паразитирует половозрелая стадия; Б — промежуточный хозяин (свинья), пожирающая яйца (по Ноблю, изменено): / — яйцо цепня с онкосферой внутри, 2 — финна, развивающаяся в мышцах свиньи, 3—половозрелая стадия, 4 — отдельный зрелый членик, выходящий из кишечника человека. Заражение человека возможно при употреблении в пищу непрожаренной свинины Рис. 163. Схема жизненного цикла широкого лентеца: 1 — окончательный хозяин (человек), в котором паразитирует половозрелая стадия, 2 — выходящие из кишечника человека во внешнюю среду яйца лентеца, 3 — вылупление корацидия, 4 — свободный корацидий, 5 — первый промежуточный хозяин (циклоп) с процеркоидом в полости тела, 6 — второй промежуточный хозяин (рыба) с пле-роцеркоидом в мускулатуре. Человек заражается, употребляя в пищу плохо проваренную рыбу с плероцеркоидами 188 током крови заносится в различные внутренние органы: чаще всего — в печень, мышцы, реже — в легкие, мозг и др. Здесь онкосфера застревает, останавливается и испытывает превращение в стадию финны (рис. 161, А, Б).Превращение начинается с быстрого роста онкосферы, которая достигает размера крупной горошины. Тело личинки становится полым, представляя собой пузырь, наполненный жидкостью. Это и есть финна, или пузырчатая глиста. Стенки пузыря образуют в одной точке впячивание, на дне которого на внутренней поверхности появляются в виде 4 ямок зачатки присосок, а между ними, на самом дне, венчик небольших крючков. Это впячивание — зачаток головки ленточной глисты, но только ввороченной внутрь пузыря финны. В теле свиньи дальнейшее развитие финны не имеет места, но финна может несколько лет просуществовать в своем промежуточном хозяине, не погибая. Для достижения половозрелого состояния финна должна попасть в кишечник окончательного хозяина, т. е. человека. Попав вместе с недостаточно прожаренным или просоленным и т. п. мясом в кишечник человека, финны Т. solium освобождаются из мяса под влиянием пищеварительных соков, а затем головка финны (главным образом под действием желчи) выворачивается наружу, причем крючья и присоски принимают свое нормальное положение. Сморщенный пузырь финны некоторое время висит на конце шейки сколекса в виде хвостового пузыря, а затем отпадает и разрушается. Головка с шейкой начинают усиленно расти и посредством поперечных насечек отшнуровывать на заднем конце последней все новые членики, образуя длинную цепь проглоттид. Итак, цикл развития Т. solium (рис. 162) связан со сменой хозяев и со сложным превращением — онкосферы в финну, а финны в ленточную половозрелую стадию. 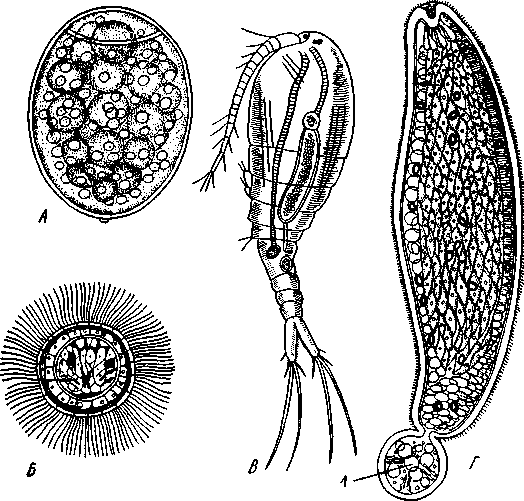 Рис. 164. Стадии развития широкого лентепа (по Скрябину, Шульцу, Розе- ну). А — сложное яйцо; Б — корацидий; В — процеркоид в полости тела зараженного циклопа; Г — процеркоид; Д — плероцеркоид: |