А-Догель - Зоология беспозвоночных_2. Тип губки (spongia, или porifera)
 Скачать 14.77 Mb. Скачать 14.77 Mb.
|

|
Отряд. 1. Caryophyllidea. Типичный представитель отряда — гвоздичник (Caryophylla-eus),живет в кишечнике карповых рыб. Тело паразита не расчленено и имеет единственный половой аппарат (рис. 169, А).Фестончатый передний конец функционирует как головка. Личинки (снабженные хвостовым придатком процер-коиды) живут в полости тела малощетинковых червей. Близкий родич гвоздичника Archi-getes (рис. 169, Г) сохраняет в течение всей жизни хвостовой придаток и достигает половой зрелости в полости тела малощетинковых червей. Это единственный представитель цес- торого протекает в беспозвоночных. Возможно, что Arcki-getes — это неотеническая личинка, взрослая стадия которой утрачена. Отряд 2. Pseudophyllidea объединяет ленточных червей, сколекс которых несет всего две присасывательные ямки и иногда одиночные крючья. Матка открывается наружу само- Рис. 169. Анатомия нерасчлененных Cestoda. А — об- тод- половое размножение ко - -• - щий вид Caryophyllaeus laticeps из кишечника карповых рыб; Б — его личинка (по Фурману); В — детали строения его заднего конца тела; Г — Archigetes ар-pendiculatus из полости тела пресноводных олигохет (по Мрачеку): / — сколекс. 2 — семенники, 3 — желточники, 4 — семяпровод, 5—сумка копулятивного аппарата, 6 — яйцевод, 7 — желточный проток, 8 — скорлуповые железы, 9 — яичник, 10 — семяприемник, // — влагалище, 12 — матка, 13 —женское половое отверстие, 14 — мужское половое отверстие, 15 — копу-лятивный орган, выпяченный наружу, 16 — хвостовой придаток, 17 — личиночные крючья 196 стоятельным отверстием (см. рис. 160, Б).Наибольшее значение имеют два семейства: ремнецы — Ligulidae и лентецы — Diphyllobothriidae. Ремнецы характеризуются наличием длинного лентовидного тела, содержащего многократно повторяющийся половой аппарат, но не всегда разбитого на членики; головка обособлена неявственно. У семейства лентецов тело расчленено, головка же в качестве органов прикрепления несет две боковые щелевидные ямки (см. рис. 154, В), Отряд 3. Cyclophyllidea. Включает цестод, имеющих сколекс с четырьмя присосками и венчиком кутикулярных крючьев. Матка не имеет отверстия (см. рис. 159, В) и наполненные яйцами членики целиком отделяются от стробилы. Сюда относится обширное семейство солитеров, или цепней (Taeniidae), включающее много важных паразитов человека и животных. КЛАСС V. ЦЕСТОДООБРАЗНЫЕ (CESTODARIA) К этому классу относится небольшое число видов, сгруппированных в два отряда. Представители цестодообразных во многих отношениях сходны с 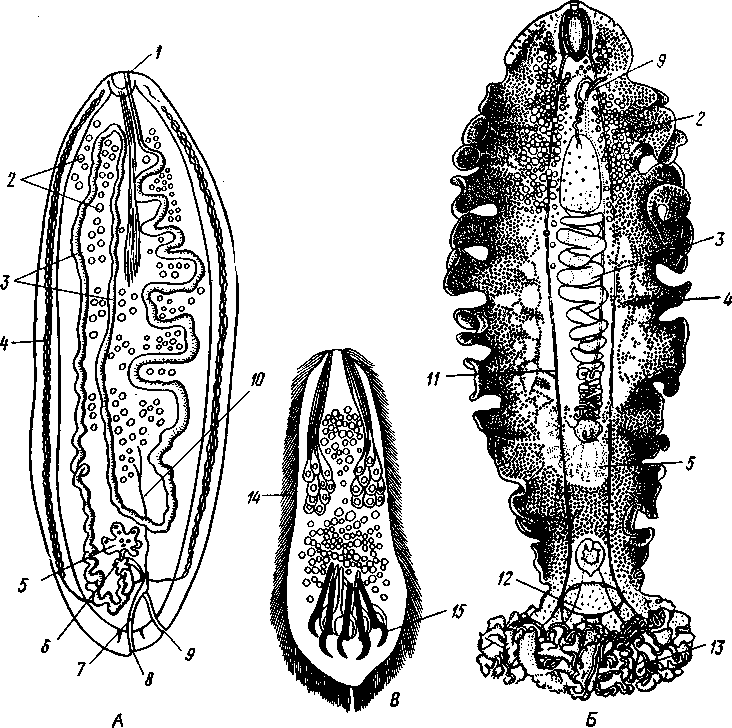 Рис. 170. Представители класса Cestodaria. А — анатомия амфилины Amphilina foliacca из полости тела стерляди (по Гейну); Б — строение Gyrocotyle urna из кишечника химеры (по Линчу); В — личинка Gyrocotyle (из Шульца и Гвоздева): / — отверстие матки, 2 — семенники, 3—матка, 4 — желточники, 5—яичник, 6 — оотип, 7 — влагалище, 8 — отверстие влагалища, 9—мужское половое отверстие, 10 — семяпроводы, // — нервный ствол, 12 — нервное кольцо, 13 — прикрепительная розетка. 14 — ресничный покров личинки, Ib — эмбриональные крючочки 197 цестодами и лишены, подобно последним, пищеварительной системы. Это обстоятельство послужило причиной того, что цестодообразных нередко относят к классу Cestoda как подкласс. Между тем цестодообраз-ные обладают рядом признаков, отличающих их от ленточных червей. Прежде всего это касается строения личинки — ликофоры (рис. 170.В), которая несет не 6 как онкосфера цестод, а 10 эмбриональных крючочков. В отличие от цестод церкомер ликофоры часто не отбрасывается, а сохраняется и у половозрелых особей. Все Cestodaria — нерасчлененные формы, обладающие единичным половым аппаратом. Половая система, как и у прочих плоских червей, гермафродитна и представлена многочисленными фолликулярными семенниками и одним двухлопастным яичником. Желточники развиты, матка открывается наружу самостоятельным отверстием. Имеется влагалище. Все прочие системы органов устроены принципиально так же, как у ленточных червей. Отряд 1. Gyrocotylidea. Паразитирует в кишечнике древних акуловых рыб — химер. Тело гирокотилид уплощено, края имеют фестончатые разращения (рис. 170,5). На заднем конце тела расположен прикрепительный диск в виде складчатой розетки, на переднем — небольшая присоска. Прикрепительная розетка иннервируется мощным нервным кольцом, которое соединяет боковые нервные стволы. Развитие идет с метаморфозом. Вылупляющаяся из яйца личинка имеет на заднем конце диск с крючочками (церкомер), из которого в дальнейшем возникает прикрепительная розетка взрослой формы. Своеобразие отр. Gyrocotylidea заключается в том, что представители его сочетают признаки классов моногеней и ленточных червей, занимая как бы промежуточное положение между ними. Отряд 2. Amphilinidea. Наибольший интерес представляет вид Amphilina fo-liacea (рис. 170, Л) с овальным, листовидным телом, достигающим 5 см длины. Паразит встречается в осетровых рыбах Волжско-Каспийского бассейна и рек Сибири. В половозрелом состоянии живет не в кишечнике, а в полости тела хозяина. Промежуточными хозяевами для амфилины служат некоторые виды бокоплавов (отр. Amphipoda) и расщеп-ленноногих рачков (отр. Mysidacea). В полости их тела живет личинка типа процеркоида, которая после съедания рачков рыбой превращается в половозрелую стадию. В связи с необычным местообитанием взрослой Amphilina (полость тела вместо кишечника) некоторые ученые трактуют взрослую амфилину как пеотеническую личинку типа плероцер-коида, приобретшую способность размножаться половым путем. Филогения плоских червей и вопрос о происхождении паразитизма Плоские черви связаны в своем происхождении с предками примитивных кишечнополостных. В основании филогенетического древа Plathelminthes, несомненно, стоит класс ресничных червей (Turbellaria), возможные пути происхождения которых от гипотетических планулооб-разных организмов рассмотрены выше (с. 161). Примитивным строением, как указывалось, характеризуются бескишечные турбеллярии (Асое-1а), которые и дали, по-видимому, начало остальным ресничным червям. Однако наибольший интерес для понимания филогенетических взаимоотношений в пределах типа Plathelminthes в целом представляют прямокишечные турбеллярии. Схематизируя описанную выше (с. 159) организацию Rhabdocoela, мы получаем своего рода обобщенный прототип всех плоских червей в виде животного с простой прямой кишкой, мозгом, 2—6 нервными стволами, парой протонефридиев, парными гонадами и хитинизированиым копулятивным аппаратом. От этого прототипа могут быть произведены другие, более специализированные в разных отношениях группы Plathelminthes. По-видимому, все современные паразитические плоские черви ведут свое начало от свободноживущих форм, общих с предками прямокишечных турбеллярии и морфологически близких к ним. Эволюция этих предковых форм шла в трех оспов- 198 ных направлениях: одна ветвь дала начало современным Rhabdocoela, две другие — пошли по пути приспособления к паразитическому образу жизни. Многим современным турбелляриям и, в частности, прямокишечным свойственна способность к сожительству (симбиозу) с другими животными. Известны виды Rhabdocoela, находящие себе убежище в выводковой камере или жаберной полости некоторых ракообразных. Есть и настоящие паразитические виды, поселяющиеся в моллюсках, морских звездах, рыбах и т. д. Это позволяет предполагать, что и предки прямокишечных турбеллярий при возникновении подходящих условий могли становиться паразитами. Переход свободноживущих животных к паразитизму может осуществляться несколькими путями. Важное значение в этом плане имеют различные типы симбиоза, в том числе так называемое «квартирантст-во», при котором один из партнеров, отличающийся более мелкими размерами, находит себе убежище на теле другого более крупного животного. Такими «квартирантами», возможно, были турбелляриеобраз-ные предки современных моногеней. Оседая на поверхности жабр, кожи или плавников рыб, они первоначально питались оседавшими здесь же мелкими беспозвоночными или слизью, выделяемой покровами рыбы. Вероятно, что такие квартиранты вместо того, чтобы довольствоваться не всегда имеющейся посторонней пищей, начнут делать ранки на теле хозяина и найдут в нем неистощимый источник питательного материала. Постепенно перейдя к питанию кровью и тканями хозяина, они станут, таким образом, настоящими эктопаразитами. Переход древних моногеней к паразитизму был связан, вероятно, с появлением хрящевых рыб (химер и акул), следовательно, это могло быть приурочено к силуру или девону. Главное направление эволюции кл. Monogenoidea выражается в совершенствовании прикрепительного аппарата, обеспечивающего постоянную связь паразита с телом хозяина. В отдельных случаях эктопаразитизм может далее вести к внутреннему паразитизму. Так, многие моногеней всю жизнь проводят на жабрах рыб. Такой же образ жизни ведут на жабрах головастиков личинки лягушачьей многоустки и поколение червей этого вида, успевающее завершить развитие до окончания метаморфоза хозяина-. При зарастании жабр головастика личинки следующей генерации многоусток мигрируют в мочевой пузырь молодой лягушки. Процесс, который совершается здесь в течение индивидуального развития одного из поколений, надо думать, имел место и в филогении рода Polystoma, предки которого жили либо на рыбах, либо на водных амфибиях, всю жизнь дышащих жабрами. Важно отметить и то, что некоторые виды современных моногеней паразитируют в ротовой полости, глотке и пищеводе рыб. Очевидно, в ходе эволюции имело место постепенное перемещение червей, исходно паразитировавших на жабрах, в ротовую полость и далее по пищеварительному тракту. В филогенезе — это вероятный путь перехода от экто-паразитизма к паразитированию в кишечнике хозяина. Именно таким путем от древних моногенообразных предков могли возникнуть классы цестод и цестодэобразных. Последние представляют в этом смысле особый интерес, так как в организации паразитирующего в кишечнике химер Gyrocotyle удивительным образом сочетаются признаки строения моногенетических сосальщиков и ленточных червей. По-видимому, Gyrocolyle можно рассматривать как промежуточное звено между кл. Cestoda и кл. Monogenoidea. Филогенетическая близость этих трех классов находит подтверждение в наличии у них церкомера, отсутствую- 1-99 щего у трематод и турбеллярий. Крайне характерно для эволюции цестод и цестодообразных вызванное эндопаразитическим образом жизни упрощение их организации, проявляющееся в полном исчезновении пищеварительной системы и органов чувств и в значительном увеличении продуктивности их половой системы. Жизненные циклы представителей Monogcnoidca, Cestoda и Cestoda-ria проходит с метаморфозом, но без гетерогонии. В отдельных случаях (эхинококк) вторично возникает метагенез как средство для увеличения потомства. В ходе эволюции цестод и некоторых цестодообразных (ам-филина) возникли промежуточные хозяева, играющие роль передаточного звена, необходимого для заражения окончательных хозяев — позвоночных. В расселении эктопаразитов участие промежуточных хозяев не дает никаких преимуществ. Вероятно, именно с этим связано отсутствие промежуточных хозяев в жизненном цикле моногеней. Становление в филогенезе современного класса трематод шло, по-видимому, несколько иначе, хотя в основе приспособления к паразитизму и здесь лежали симбиотические отношения. Жизненный цикл современных сосальщиков не может осуществляться без обязательного участия моллюсков. Вероятно, что именно моллюски и были филогенетически первыми хозяевами трематод. Нетрудно представить возникновение симбиоза между моллюсками и свободпоживущими предками ди-генетичсских сосальщиков. Личинки последних, похожие на прямокишечных турбеллярий, вероятно, обитали вблизи дна, под камнями, или использовали в качестве укрытия раковины моллюсков. При этом они могли легко попадать в жаберную полость моллюсков (с. 454) и поселяться там, становясь, таким образом, квартирантами. Разумеется, такой переход от свободного существования к квартирантству должен был совершаться очень медленно, и личинки на протяжении длительного времени сохраняли еще связь с внешней средой. В дальнейшем такие квартиранты, первоначально питавшиеся, вероятно, независимо от моллюсков, могли перейти к питанию его кровью, становясь, таким образом, на путь адаптации к паразитизму. Постепенно это должно было привести к переселению паразитов внутрь тела моллюска. Защищенные организмом хозяина от неблагоприятных воздействий внешней среды, они получили вместе с тем постоянный источник пищи в виде соков и тканей хозяина. Это обусловило новое направление эволюции предков трематод, выразившееся в постепенном и все более глубоком упрощении их организации и ускорении созревания. Личинки, которые раньше покидали своего хозяина и достигали половой зрелости во внешней среде, перестали выходить наружу и начали размножаться в теле хозяина. Так как, по-видимому, у предков трематод долгое время еще сохранялось и свободноживущее поколение, то жизненный цикл приобрел характер гетерогонии, т. е. чередования двух различающихся половых поколений. Разница же заключалась в том, что особи паразитического поколения под влиянием паразитизма упростились и из гермафродитов превратились в партеногенетических самок, тогда как свободное поколение оставалось гермафродитным. С появлением костистых рыб в верхнем триасе жизненный цикл предков трематод начал осуществляться с участием не одного, а двух хозяев. Столь «поздний» переход гермафродитного поколения к паразитизму находит свое отражение в том, что оно сохранило еще все черты организации, свойственные свободноживущим плоским червям (сложные половая и выделительная системы, пищеварительная система и т. д.). Жизненный цикл с участием двух хозяев — моллюска и позвоночного, характерный, например, для печеночной двуустки, обычно рассмат- 200 ривают как исходный примитивный тип жизненного цикла. По-видимому, лишь много позднее установился характерный для большинства современных дигенетических сосальщиков жизненный цикл, который проходит с участием трех хозяев. При этом появление второго промежуточного хозяина, служащего пищей окончательному, рассматривают как приспособление, обеспечивающее заражение последнего. Дополнение к типу Plathelminthes. Mesozoa Сюда относятся несколько небольших групп эндоиаразитических животных, многоклеточных, но устроенных, .несомненно, значительно проще, чем все прочие Metazoa. Они не обнаруживают дифференцировки на типичные зародышевые листки, так как под наружным эпителием, одевающим их тело, имеется или всего одна осевая клетка, или же просто скопление половых клеток. Вплоть до начала XX в. Mesozoa придавали большое филогенетическое значение как формам, переходным между Protozoa и Metazoa. В настоящее время более или менее установился взгляд, что это животные, упростившиеся под влиянием эндопаразитического образа жизни. Согласно одним авторам, это упрощенные Coelenterata, на что указывает некоторое сходство их с личинкой планулой, по другим — Mesozoa — какие-то упрощенные плоские черви. Второе мнение кажется нам более основательным: Во всяком случае группу Mesozoa следует считать провизорной, сохраняющейся до тех пор, пока не удастся связать их генетически с какими-нибудь высшими Metazoa. ,К Mesozoa принадлежат два класса (Orthonectida и Dicyemida), содержащие всего около 60 видов. КЛАСС !. ОРТОНЕКТИДЫ (ORTHONECTIDA) Небольшая группа мелких (меньше 1 мм) организмов, паразитирующих в полости тела или в половых железах различных морских животных (Turbellaria, Nemcrtini, Poly-chacta, Mollusca и Ophiuroidea). Orthonectida — раздельнополы. Самцы и самки имеют продолговатое тело, состоящее из мерцательного наружного эпителия и внутреннего скопления мужских или женских половых клеток (рис. 171). Половые особи покидают хозяина и выходят в воду, где и происходит оплодотворение. 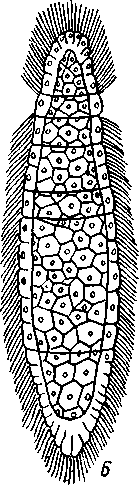 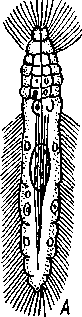 Яйца развиваются внутри оплодотворенных самок в мелких мерцательных личинок; последние покидают материнский организм и активно проникают в подходящего хозяина. Наружный слой клеток личинки при этом погибает, а внутреннее скопление клеток сливается в небольшой плазмодий, питание которого происходит с помощью многочисленных выростов, внедряющихся в ткани хозяина. Число ядер плазмодия увеличивается. Большая часть ядер имеет вегетативный характер, но некоторые увеличиваются в размерах и вокруг них обособляется участок более плотной цитоплазмы. Получившиеся клетки начинают дробиться и в одних плазмодиях разиваются в самцов, в других — в самок. Клетки, дающие начало самцам и самкам, по нашему мнению, являются партеногенетическими яйцами. В таком случае жизненный цикл Orthonectida представляет собой гетерогонию, т. е. чередование типичного полового (самца и самки) и партеногене-тического (плазмодии) поколений. Некоторые авторы, однако, считают генеративные клетки Orthonectida за особые бесполые клетки, или агаметы, способные бесполым путем давать начало половому поколению. Представители: Rhopalura (в офиурах), Stocchar-thrum (в полихетах). Рис. 171. Rhopalura ophiocomae (класс Orthonccthida). A — самец; Б — самка (из Клауса] 201 |