Документ Microsoft Word. Типы мышечных волокон и регуляция их функций
 Скачать 243.81 Kb. Скачать 243.81 Kb.
|

|
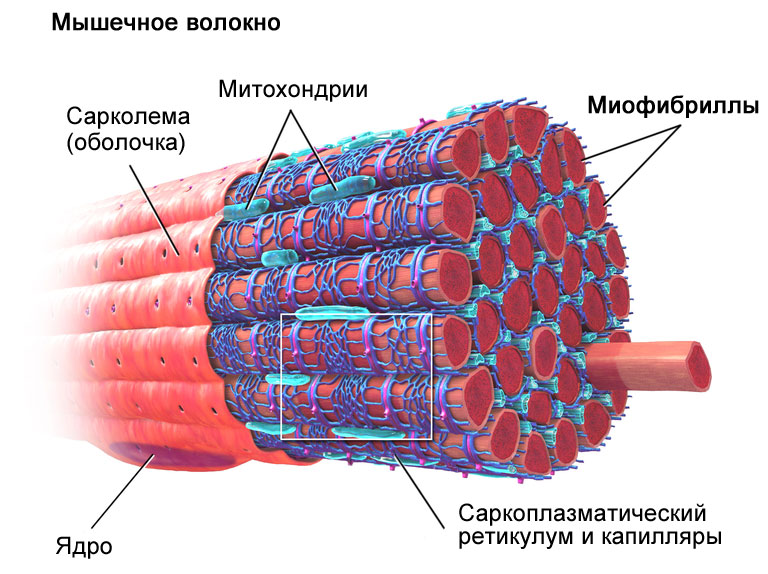
государственное бюджетное образовательное учреждение высшего профессионального образования «Тюменский государственный медицинский университет Министерства здравоохранения Российской Федерации» (ГБОУ ТюмГМУ Минздрава РФ) Кафедра гистологии с эмбриологией имени ЗДН РФ проф. Дунаева П.В. Заведующий кафедрой: д.м.н., профессор Соловьев Г.С. РЕФЕРАТ На тему «Типы мышечных волокон и регуляция их функций» Выполнил: студент 156 группы Осипов Никита Сергеевич Руководитель: ассистент кафедры Морозова Елена Васильевна Тюмень 2018 г. Содержание: Введение.......……………………………………………………………..…3 Состав мышечного волокна……………………………………………….4 Типы мышечных волокон…………………………………………………5 Быстрые и медленные мышечные волокна………………………………8 Гликолитические, промежуточные и окислительные волокна…………10 Миозин………………………………………………………………………15 Список литературы…………………………………………………………17 Введение Мышцы – это важнейшая составляющая опорно-двигательного аппарата, обладающая сократительной способностью. Именно благодаря возможности мышечных тканей сокращаться, человек может выполнять всяческие движения, начиная с самых простых (моргание и улыбка) и заканчивая максимально тонкими (как у ювелиров) и энергичными (как у спортсменов). Функциональность мышечного скелета напрямую связана с составом его главных структурных единиц – мышечных волокон. Волокна скелетных мышц соединяются со спинным мозгом посредством толстых нервных волокон. После попадания в мускул каждое из нервных волокон делится на сотни разветвлений, которые снабжают сотни мышечных волокон. Соединение между нервом и волокном мышечной ткани называют синапсом, или нервно-мышечным соединением. Примечательно, что на каждом мышечном волокне может формироваться только один синапс. При соответствующем нервном сигнале возникает потенциал действия, который передается по нервам от спинного мозга к мускулам. От свойств мышечных волокон зависит то, как мускулатура адаптируется к повторяющимся сигналам. Именно типы волокон обуславливает предрасположенность спортсмена к той или иной тренировочной программе. Во время тренировки происходит гипертрофия мышечных волокон – увеличение их объема и массы. При этом важно понимать, что количество волокон не изменяется и обуславливается генетическими особенностями того или иного человека. Состав мышечного волокна В состав мышечного волокна входят:
Типы мышечных волокон У людей все волокна скелетных мышц имеют разные механические и метаболические свойства. Различные типы мышечных волокон определяют по максимальной скорости их сокращения (быстрой и медленной) и главного метаболического пути, который они используют для образования АТФ (окислительный и гликолитический). Мышечные волокна в целом делятся на:
Поскольку скорость сокращения самых быстрых мышечных волокон несколько выше, чем скорость сокращений волокон IIb типа, самые быстрые волокна называются в литературе волокнами IIх типа. Иногда выделяют волокна IIс типа — эти волокна не похожи на волокна ни I, ни II типа. Они проявляют как окислительную, так и гликолитическую активность и представлены лишь в небольшом количестве (около 1 %). В зависимости от типа тренировок они могут переходить в волокна I или II типа. Мышечные волокна возбуждаемые одним мотонейроном входят в состав одной двигательной единицы (ДЕ). Ске летные мышцы человека состоят из ДЕ всех трех типов. Одни из них включают преимущественно медленные ДЕ, другие — быстрые, третьи — и те, и другие.
Таблица 1. Критерии разделения на типы Быстрые и медленные мышечные волокна Классифицируются по активности фермента миозиновой АТФ-азы и, соответственно, по скорости сокращения мышц. Волокна, содержащие миозин с высокой активностью АТФазы, относят к быстрым волокнам, а те, что содержат миозин с более низкой активностью АТФазы, - к медленным. Активность АТФазы наследуется и тренировки не влияют на соотношение быстрых и медленных волокон. Освобождение энергии, заключенной в АТФ, осуществляется благодаря АТФ-азе. Энергии одной молекулы АТФ достаточно для одного поворота (гребка) миозиновых мостиков. Мостики расцепляются с актиновым филаментом, возвращаются в исходное положение, сцепляются с новым участком актина и делают гребок. Скорость одиночного гребка одинакова у всех мышц. Для очередного гребка требуется новая молекула АТФ. В волокнах с высокой АТФ-азной активностью расщепление АТФ происходит быстрее, и за единицу времени происходит большее количество гребков мостиками, то есть мышца сокращается быстрее и, соответственно, сильнее. Медленные окислительные волокна содержат множество митохондрий и обладают высокой способностью к окислительному фосфорилированию. Эти волокна могут содержать значительное количество липидов, но меньшее количество гликогена. Большая часть АТФ, произведенного такими волокнами, зависит от снабжения крови кислородом и топливных молекул. Эти волокна окружают многочисленные капилляры. Они также содержат большое количество связывающего кислород миоглобина, который увеличивает поглощение кислорода тканями и способствует небольшому внутриклеточному накоплению кислорода. Миоглобин придает темно-красный цвет, поэтому окислительные волокна часто называют красными мышечными волокнами. 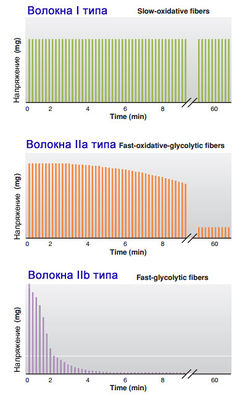 В быстрых волокнах, также названных гликолитическими волокнами, напротив, содержится мало митохондрий, но они обладают высокой концентрацией гликолитических ферментов и большим запасом гликогена. Из-за ограниченного использования кислорода их окружает относительно небольшое количество капилляров, и они содержат мало миоглобина. Их называют белыми мышечными волокнами вследствие их более светлого цвета по сравнению с красными окислительными волокнами. Гликолитические, промежуточные и окислительные волокна Гликолитические волокна, как правило, намного больше в диаметре, чем окислительные волокна. Чем больше диаметр, тем больше максимальное растяжение, которого они могут достичь (т.е. тем они сильнее). Классифицируются по окислительному потенциалу мышцы, то есть по количеству митохондрий в мышечном волокне. Митохондрии – это клеточные органеллы, в которых глюкоза или жир расщепляется до углекислого газа и воды, ресинтезируя АТФ, необходимую для ресинтеза креатинфосфата. Креатинфосфат используется для ресинтеза миофибриллярных молекул АТФ, которые используются для мышечного сокращения. Вне митохондрий в мышцах также может происходить расщепление глюкозы до пирувата с ресинтезом АТФ, но при этом образуется молочная кислота, которая закисляет мышцу и вызывает ее утомление. По этому признаку мышечные волокна подразделяются на три группы:
У не тренирующихся людей обычно быстрые волокна - гликолитические и промежуточные, а медленные – окислительные. Однако при правильных тренировках на увеличение выносливости быстрые мышечные волокна превращаются из гликолитических в промежуточные. Также возможен переход промежуточных волокон в окислительные. При силовых тренировках промежуточные волокна могут переходить в гликолитические. При этом соотношение медленных и быстрых волокон генетически предопределено практически не меняется вне зависимости от тренировок (переход не более 1-3%).
Рис. 2 Срез мышечных волокон I (темные) и IIa (светлые) и IIb/IIx типа (А) у пожилых и (B) молодых людей. С возрастом количество волокон II типа сокращается. Окраска АТФазы миозина. Двигательная единица - это один мотонейрон и иннервируемые им мышечные волокна. Когда МО мотонейрон стимулирует свои волокна, сокращается гораздо меньше мышечных волокон, чем когда свои волокна стимулирует мотонейрон БГ. Следовательно, двигательные БГ волокна достигают пикового напряжения быстрее и, взятые вместе, развивают большее усилие, чем МО волокна. Волокна скелетных мышц различаются также по их способности противостоять усталости. Утомление БГ волокон происходит быстрее, тогда как МО волокна очень устойчивы к усталости. Быстро окисляющиеся волокна обладают промежуточной способностью сопротивляться усталости. Все мышцы человека обладают разным процентным соотношением БГ и МО мышечных волокон. В зависимости от доли имеющихся типов волокон, мышцы могут значительно различаться по максимальной скорости сокращения, силе и утомляемости. Например, в икроножных мышцах наблюдается преобладание БГ волокон, придающее им способность к сильному и быстрому сокращению, которое используется, например, при прыжках. С другой стороны, в камбаловидной мышце больше МО мышечных волокон, и она используется при длительной активности мышц ног. В целом, МО мышечные волокна обладают высоким уровнем аэробной выносливости. Способность поддерживать мышечную активность в течение длительного времени известна как мышечная выносливость. Так как МО волокна обладают высокой аэробной выносливостью, они чаще всего задействуются во время нагрузок на выносливость (например, в марафонском беге) и во время большинства повседневных занятий, где требования к мышечной силе невысоки (например, ходьба). БГ мышечные волокна, с другой стороны, обладают относительно низкой аэробной выносливостью. При нормальной, малоинтенсивной деятельности БГ волокна используются довольно редко, но при «взрывных» нагрузках они преобладают. Предполагается, что они активизируются, когда во время физической нагрузки оказывается превышен анаэробный порог; тогда уровень молочной кислоты в крови и в мышечных волокнах начинает повышаться немного раньше. БГ двигательные единицы генерируют гораздо большую силу, чем МО двигательные единицы, но они легко устают из-за своей ограниченной выносливости. Поэтому БОГ волокна, по всей видимости, используются в основном при непродолжительной интенсивной нагрузке на выносливость, например при пробежке на 1 милю или заплыве на 400 м. Как было доказано, спортивные тренировки не меняют относительное соотношение БГ и МО волокон. По всей видимости, напротив, - это почти полностью определяется генетическим наследованием, и это, в свою очередь, может определять основные спортивные способности разных людей. С практической точки зрения двигательный нерв определяет тип мышечных волокон в двигательной единице. Если нерв, иннервирующий медленную двигательную единицу, отрезать и соединить с другим нервным волокном, иннервирующим быструю двигательную единицу, эта прежде быстрая двигательная единица может постепенно измениться и стать медленной. В мышечном веретене мышц также содержатся совершенно разные типы мышечных волокон. Эти структуры воспринимают напряжение мышц. Чувствительность мышечных веретен может быть отрегулирована при сокращении их особых интрафузальных мышечных волокон. Веретена расположены параллельно основной мышце или экстра-фузальным волокнам. Уровнем сокращения интрафузальных мышечных волокон в веретенах управляют гамма-мотонейроны, тогда как альфа-мотонейроны регулируют экстра-фузальные мышечные волокна, которые непосредственно отвечают за сокращение мышц. Высокопороговые и низкопороговые волокна Классифицируются по уровню порога возбудимости двигательных единиц. Мышца сокращается под действием нервных импульсов, которые имеют электрическую природу. Каждая двигательная единица (ДЕ) включает в себя мотонейрон, аксон и совокупность мышечных волокон. Количество ДЕ у человека остается неизменным на протяжении всей жизни. Двигательные единицы имеют свой порог возбудимости. Если нервные импульсы, посылаемые мозгом, имеют частоту ниже этого порога, ДЕ пассивна. Если нервные импульсы имеют пороговую для этой ДЕ величину или превышают ее, мышечные волокна активируются и начинают сокращаться. Низкопороговые ДЕ имеют маленькие мотонейроны, тонкий аксон и сотни иннервируемых медленных мышечных волокон. Высокопороговые ДЕ имеют крупные мотонейроны, толстый аксон и тысячи иннервируемых быстрых мышечных волокон. Медленные окислительные волокна относятся к низкопороговым (возбуждаются при незначительной нагрузке). Быстрые волокна относятся к высокопороговым (включатся только при интенсивной нагрузке). Миозин Существенное различие разных видов мышечных волокон обуславливает значительную гетерогенность мышечных тканей и их способность к выполнению разнообразных функциональных задач. Биохимический и иммуногистохимический анализ скелетных мускулов показывает, что структурное и функциональное разнообразие мышечных волокон обуславливается широким спектром изоформ миозина. Миозином называется фибриллярный белок, выступающий одним из главных компонентов сократительных мышечных волокон. Он составляет от 40 до 60% общего количества мышечного белка в организме. При соединении миозина с актином (еще один мышечный белок) образуется актомиозин – основной элемент сократительной системы мускулов. В состав молекулы миозина входит две тяжелых цепи (MyHC) и четыре легких (MyLC). Тяжелые цепи имеют несколько изоформ, свойства которых обуславливают силовые и скоростные показатели мышечных волокон. Наиболее важными считаются четыре изоформы: MyHCI, MyHCIIA, MyHCIIX/IID, и MyHCIIB. Каждая изоформа имеет специфическую скорость сокращения и позволяет развить определенное усилие. Волокна, в состав которых входит MyHCI, по сравнению с волокнами, содержащими другие формы тяжелой цепи миозина, медленнее сокращаются и развивают меньшее усилие. Наиболее быстрыми и сильными считаются волокна, содержащие MyHCIIB изоформу тяжелой цепи. За ними следует MyHCIIX и MyHCIIA форма. Физическая активность может привести к весомым изменениям сократительных свойств мускулов. Принято считать, что при тренировке на выносливость увеличивается количество медленных изоформ миозина. Вместе с тем во время силовой тренировки происходит увеличение количества MyHCIIA и уменьшение MyHCIIX. Кроме того, считается, что у основной массы людей, активность которых ограничивается простыми бытовыми делами, волокна, содержащие миозин в форме MyHCIIX, крайне редко вовлекаются в работу. В процессе физической тренировки они начинают задействоваться и постепенно переходят в MyHCIIA форму. Дело в том, что волокна, содержащие IIA изоформу тяжелой цепи миозина, имеют большую выносливость, по сравнению волокнами IIX типа. Во время тренировок выносливости или силы происходит весомое изменение гормонального фона скелетных мускулов, которое служит мощным сигналом, запускающим процесс изменения состава миозина в мускулах, подвергающихся нагрузке. Список литературы
|