Типы популяций. Типы популяций
 Скачать 47.39 Kb. Скачать 47.39 Kb.
|

|
Типы популяций В природе существуют различные популяции. С точки зрения зоологов и ботаников широкого профиля, в том числе систематиков и биогеографов, в рамках вида прежде всего имеются географические популяции. К ним принадлежат группы особей данного вида, которые населяют географически однородную, но биогеоценологически разнообразную местность. Это — внутривидовые группировки обширных географических пространств. Например, один из видов божьих коровок — семиточечная (Coccinella septempunctata)—обладает огромным ареалом, охватывающим Европу, Азию и Северную Африку, но образует всего четыре географические популяции. Они различаются между собой по выносливости к температуре, по количеству генераций в течение года, по времени размножения и периоду высокой активности. На обширной области распространения большой синицы (Parus major) известно пять ее географических (популяций (рис. 46). У некоторых из них существуют симпатрические территории, но скрещивания не происходит. 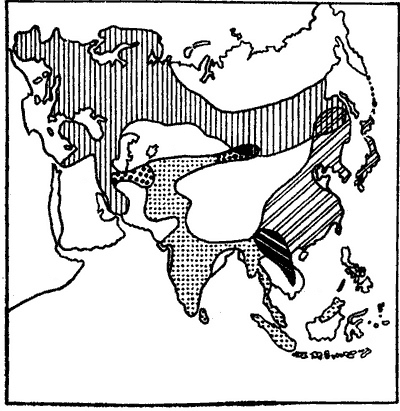 Рис. 46. Ареалы географических популяций большой синицы (по: Дажо, 1975). Наряду с географическими популяциями в их рамках известны экологические популяции. Их также называют местными, локальными, или ценопопуляциями. Именно эти экологические популяции и составляют предмет последующего изложения. Если говорить о малоподвижных, а тем более о прикрепленных видах, то тогда под экологической популяцией следует подразумевать совокупность всех особей данного вида, которые входят в какой-либо один биоценоз (отсюда термин «ценопопуляция»), Точнее сказать, экологическая популяция — пространственно ограниченная внутривидовая группировка, в основе которой лежит известная общность происхождения, сходство облика (фенотипа), единство экологических реакций на внешние воздействия. Эта последняя особенность — функциональное единство — важный критерий экологической популяции. В соответствии с ним все члены популяции реагируют на одинаковые воздействия среды не вразнобой, а единообразно и одновременно, причем в силу не только определенного наследственного родства и генетически обусловленных реакций, но и (особенно у птиц) высоко развитых подражательных рефлексов. Выше мы отмечали взаимное родство членов экологической популяции, однако ему не следует придавать того ведущего значения, которое подчеркивается в генетических популяциях. Дело в том, что, как будет показано дальше, в постнатальном онтогенезе птиц и других животных существует обязательный этап расселения молодняка, т. е. его дисперсии, а также наблюдается рассеяние в пространстве более взрослых особей. Все это приводит к тому, что видовое население в каждом пункте неизбежно становится генетически разнородным. По тем же причинам родственные связи не замыкаются в каких-либо узких территориальных и биологических пределах, а охватывают достаточно удаленные друг от друга группы особей. Популяция не просто занимает известное пространство, но при этом широко использует его жизненные ресурсы и возможности. В частности, среда обитания, как показал Н. П. Наумов, служит животным плацдармом для накопления и передачи взаимной информации, т. е. их биологическим сигнальным полем, наличие которого весьма способствует групповой консолидации членов популяции. В процессе своей жизнедеятельности животные более или менее глубоко влияют на окружающую природу — устраивают убежища, повреждают растения, делают запасы, роются в земле, изменяют почву и т. д. Следы этого воздействия также свидетельствуют о характере поведения животных и тем самым обогащают информацией биологические сигнальные поля соответствующих видов. Хотя экологическая популяция представляет собой единое целое, тем не менее она не самая мелкая, не самая дробная биохорологическая единица. Нередко, особенно в условиях мозаичного ландшафта, территория, занимаемая популяцией, бывает настолько пестрой, неоднородной, что это накладывает свой отпечаток на характер популяции — ее структуру, объем, адаптивные особенности и т. д. В подобного рода случаях приходится говорить о наличии в рамках экологической популяции более мелких — элементарных, или микропопуляций. Последние не являются самостоятельными формами существования вида. Микропопуляции обязательно взаимно связаны. Даже их население поддерживается лишь вследствие этих контактов. Сами по себе микропопуляции не обладают способностью к неограниченно длительному, автономному существованию. Оно обеспечивается за счет перемещения особей из одной микропопуляции в другую. Примером микропопуляций могут служить группировки лесных мышей (Apodemus sylvaticus) в лесостепи Зауралья. Там эти грызуны населяют три типа стаций: березовые колки (рис. 47), поля, заросли по берегам водоемов. Зверьки из упомянутых местообитаний различаются некоторыми морфологическими особенностями. Тем не менее, они не образуют самостоятельных популяций, поскольку группировка мышей каждой из стаций поддерживает свое существование лишь за счет взаимного обмена особями, зависящего от экологической обстановки. Особенно интенсивно перекочевки происходят весной и осенью. В общем все три группировки настолько тесно между собою контактируют, что фактически представляют единую, но сложно организованную экологическую популяцию. Рис. 47. Березовые колки в западносибирской лесостепи. Следовательно, микропопуляции являются структурными подразделениями экологической популяции. Микропопуляции, несущие на себе отпечаток влияния тех или иных особенностей среды обитания, наиболее характерны для растений и малоподвижных мелких животных. Их существование и единство поддерживается за счет перемещения особей из участков, где они могут выжить даже в самые неблагоприятные годы или сезоны. Недаром подобного рода участки получили название стаций переживания, или стаций-резерваций. Они играют очень важную роль в восстановлении численности животных после массовой их гибели в периоды депрессий. Знание стаций переживания необходимо не только в познавательных, но и в прикладных целях, например для научной организации борьбы с вредными насекомыми и грызунами. Именно здесь всего эффективнее и дешевле их истребление. |