Молекулярная биология. мол.биология. Учебнометодический комплекс Новосибирск 2015
 Скачать 187.18 Kb. Скачать 187.18 Kb.
|

|
Программа курса лекций. 1. Определение предмета "молекулярная биология". Этапы развития. Основные открытия. Доказательства генетической роли нуклеиновых кислот. Хронология открытий, подготовивших создание Уотсоном и Криком модели двойной спирали ДНК. 2. Принципы строения и основные функции биополимеров. Нуклеиновые кислоты. Нуклеозид, нуклеотид, полинуклеотид. Нерегулярные полимеры. Принципы строения двойной спирали ДНК. Виды ДНК. Параметры В-, А- и Z-форм ДНК. Виды РНК. Их роль в клетке. Функции ДНК. Информационная емкость. 3. Принципы строения и основные функции биополимеров. Белки Классификация аминокислот. Первичная и вторичная структура белка. Третичная и четвертичная структура белка. Глобулярные и фибриллярные белки. Денатурация и ренатурация белков. Фолдинг белков. Шапероны. Шаперонины. Прионы. Основные биологические функции белков. 4. Принципиальное строение биологической мембраны 5. Генетический код 6. Транскрипция у прокариот. Принципы транскрипции. Субъединичный состав РНК-полимеразы E.coli. Holo- и Core- фермент. Понятие об опероне. Особенности структуры промоторов у прокариот. Этапы транскрипции у прокариот. 7. Регуляция транскрипции у бактерий. Негативная индукция. Позитивная индукция. Негативная репрессия. Позитивная репрессия. Аттенуация в регуляции экспрессии триптофанового оперона E.coli. 8. Особенности транскрипции у эукариот. Множественность и специфичность РНК-полимераз эукариот. Понятие об экзонах и интронах. Cis-элементы транскрипции. Понятие об энхансерах . Trans-факторы транскрипции. Образование инициаторного комплекса транскрипции с участием РНК- полимеразы II. 9. Процессинг mРНК эукариот. Кепирование. Полиаденилирование. Сплайсинг. Редактирование. Различные механизмы сплайсинга. Автосплайсинг. Trans-сплайсинг. Альтернативный сплайсинг. 10.Строение иммуноглобулинов, их классификация и функции. Переключение классов иммуноглобулинов. Источники разнообразия антител. V-J рекомбинации при перестройке генов легких цепей иммуноглобулинов. V-D-J рекомбинации при перестройке генов тяжелых цепей иммуноглобулинов. 11-12. Трансляция. Структура tРНК. Рекогниция. Аминоацилирование tРНК. Структура рибосом про- и эукариот. Центры рибосом E.coli. Образование инициаторного комплекса трансляции у прокариот. Этапы трансляции у прокариот. Белковые факторы трансляции. Регуляция трансляции на примере фага MS2. Образование rРНК и белков рибосом у E.coli. Образование рибосом у эукариот. Понятие о ядрышке. 13. Репликация ДНК. Основные принципы и механизмы у про и эукариот. Принципы репликации ДНК. Доказательство полуконсервативного характера репликации. Ферментативная система синтеза ДНК in vitro. Активирование ДНК. ДНК-полимераза I из E.coli. Роль 3'5' и 5'3' гидролитических активностей. Схема непрерывной антипараллельной репликации Корнберга. Схема непрерывной параллельной репликации Кэрнса. Схема прерывистой антипараллельной репликации Оказаки. Сравнительная характеристика ДНК-полимераз I, II и III(core) из E.coli. ДНК-полимераза III*, holo-фермент. Их функции.Схема размножения фага М13 и доказательство наличия РНК-затравки при репликации ДНК. Модель «катящегося колеса». Праймаза и праймосома. Проблема денатурации матрицы при репликации ДНК . SSB. Геликазы. Принципы работы и биологические функции топоизомераз. Современная схема репликации ДНК E.coli . Репликация ДНК аденовируса человека. Репликация митохондриальной ДНК млекопитающих. Особенности репликации ядерных ДНК эукариот. Полирепликонность. 14. Проблема недорепликации 3'-концов линейных молекул. Теломеры и теломераза 15. Основные репарабельные повреждения в ДНК и принципы их исправления. 16. Уровни организации хроматина у эукариот. Общая характеристика гистонов. Нуклеосомный, супербидный, петлевой уровни компактизации ДНК эукариот. Метафазная хромосома. 17. Организация эукариотического генома. Геномы и кариотипы. Размеры и количество генов у разных таксонов. Гены "домашнего хозяйства" и гены "роскоши". Основы метода ренатурации ДНК в изучении структуры генома эукариот. Сателлитная ДНК. Особенности состава. Локализация в геноме. Палиндромы. Роль обращенных повторов в геноме. Умеренные повторы в геноме. Уники. 18. Понятие о мобильных генетических элементах. Классификация мобильных генетических элементов по механизму перемещения. Вирус иммунодефицита человека: структура провируса, белки, кодируемые вирусом. Особенности ретровирусоподобных (LTR-содержащих) ретротранспозонов Механизм обратной транскрипции ретровирусов и LTR – содержащих ретротранспозонов. Ретропозоны, не содержащие LTR (LINE и SINE элементы). Особенности организации ДНК-транспозонов. Примеры про- и эукариотических ДНК-транспозонов. Механизм интеграции ДНК-транспозонов в геном. Эффекты встройки мобильных элементов. Значение мобильных элементов в эволюции. 19. Молекулярные механизмы канцерогенеза. Этапы понимания молекулярных механизмов канцерогенеза.Многостадийность опухолевой трансформации. Основные этапы. Понятие онкогена и протоонкогена. Вирусные и клеточные онкогены. Ras онкоген, Myc онкоген. Механизмы активации протоонкогенов. Гены-супрессоры опухолеобразования План семинаров по молекулярной биологии Занятие 1. На первом занятии обсуждаются общие особенностей молекулярно-биологических методов и подходов, студенты знакомятся с классическими объектами в молекулярно-биологических иссдледованиях, обсуждаются особенности организации про- и эукариотических клеток. Затем студенты знакомятся с общими принципами клонирования ДНК. Контрольная работа на знание центрального постулата молекулярной биологии, принципов строения нерегулярных биополимеров, основных функций нуклеиновых кислот и белков. Занятие 2. Студенты знакомятся с принципами гель-электрофоретического фракционирования нуклеиновых кислот и рестрикционного анализа. Решают задачи на построение рестрикционных карт. Во время второго часа обсуждаются методы гибридизации нуклеиновых кислот (Саузерн-блот, Нозерн-блот, гибридизация insitu). Занятие 3. Студенты знакомятся с особенностями поведения аминокислот и полипептидов в растворе, вводится понятие «изоэлектрическая точка». Обсуждаются принципы гель-электрофоретического фракционирования белков, использование антител для детекции белков, Вестерн-блот анализ. Занятие 4. Первая часть занятия посвящена контрольной работе, во время которой студенты демонстрируют понимание молекулярно-биологических методов, обсуждавшихся на первых семинарах, а также знания по структуре и функциям биологических макромолекул. Вторая половина занятия посвящена решению задач на понимание свойств генетического кода. Занятие 5. Обсуждаются лекционный материал, посвященный организации оперонов E. Coli, регуляции транскрипции у прокариот. Сравнение про- и эукариотической организации транскрипции. Решаются задачи, направленные на лучшее понимание материала. Занятие 6. Первая часть семинара отводится контрольной работе, посвященной темам, связанным с транскрипцией и трансляцией. Затем обсуждается лекционный материал на тему трансляция у про- и эукариот. Занятие 7. Студенты знакомятся с принципами и практическими примерами применения полимеразной цепной реакции. Решаются задачи на тему «полиморфизм длин рестрикционных фрагментов». Обсуждается лекционный материал на тему «репликация ДНК», решаются задачи, направленные на лучшее усвоение материала. Занятие 8. Контрольная работа по теме «репликация ДНК». Затем обсуждается метод секвенирование ДНК по Сэнгеру. Образовательные технологии Формы организации учебного процесса: лекция, семинар, коллоквиум, самостоятельная работа студента, консультации, экзамен. Лекции читаются в количестве 4 часа в неделю в течение 10 недель 4-го семестра. Семинары проводятся в количестве 2 часа в неделю в течение 8 недель 4-го семестра. Используется традиционная система лекций и семинаров. На лекциях студенты получают знания о строении и функциях биополимеров, их компонентов и комплексов, об основных принципах кодирования, хранения и реализации генетической информации, структуре и функции генов и геномов. Решение задач на семинарских занятиях способствует более глубокому пониманию основных молекулярно-биологических процессов. Кроме того, на саминарах студенты знакомятся с основными принципами экспериментальных молекулярно-биологических подходов. Домашнее задание включает задачи, чтение лекций и дополнительных литературных источников. Для проверки промежуточных знаний проводятся 4 контрольные работы, на которых студенты в течение часа отвечают на теоретические вопросы и самостоятельно решают предложенные задачи. Для проверки итоговых знаний проводятся 2 коллоквиума, на которых каждый студент в подгруппе отвечает на предложенный теоретический вопрос, остальные дополняют. 6. Учебно-методическое обеспечение самостоятельной работы студентов. Оценочные средства для текущего контроля успеваемости, промежуточной аттестации по итогам освоения дисциплины Примеры контрольных работ Контрольная работа № 2 Верны ли следующие утверждения? 1 При электрофорезе в агарозном геле отдельные фрагменты ДНК мигрируют со скоростью, обратно пропорциональной их молекулярной массе: чем крупнее молекулы, тем сильнее они тормозятся сложной пространственной сеткой геля и тем медленнее продвигаются от старта. 2 Ферменты, называемые рестриктазами, разрезают двуцепочечную спираль ДНК по специфическим последовательностям, состоящим, как правило, из четырех-восьми нуклеотидов, являющихся палиндромами. 3 Плазмидными векторами, используемыми при клонировании, могут быть небольшие молекулы ДНК, которые содержат уникальные сайты рестрикции, чтобы включить чужеродную ДНК, иметь свою точку начала репликации ДНК, а так же ген, сообщающий клетке устойчивость к какому-либо антибиотику. ………………………………………. 4 Расположите олигонуклеотиды по порядку возрастания температуры плавления: АААTTGC GGG GCGCGCG AAAAAAAAAAAAAAA TTTAACG CCC CGCGCGC TTTTTTTTTTTTTTT 5 Что получится при электрофорезе смеси фрагментов ДНК: (T)150, (G≡C)150 и (T=A)150? 6 Будет ли этот фрагмент ДНК разрезаться рестриктазами EcoRI (5’-GAATCC), AluI (5’-AGCT), PstI (5’-CTGCAG)? Если да, то сколько фрагментов получится? 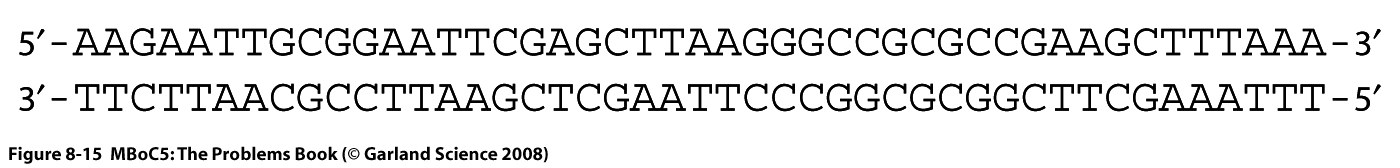 7 Линейный фрагмент ДНК обработали рестриктазой TaqI и нанесли на электрофорез, при этом получили два бэнда. Один двигался в районе 7 kb, а второй в районе 6 kb и при этом был в два раза ярче первого. Этот же фрагмент ДНК обработали рестриктазой PstI и так же разогнали на электрофорезе. При этом получили два одинаковых по яркости бэнда размером 9kb и 10 kb. При обработке этого фрагмента TaqI и PstI одновременно получают набор фрагментов 3kb, 6kb и 7 kb, при этом на электрофорезе самый короткий бэнд светится ярче остальных. Необходимо построить рестрикционную карту фрагмента ДНК. 8 Расскажите о клонировании фрагментов генома, используя перечисленные термины и по-возможности поясняя их значение Выделение ДНК Фрагментация ДНК Рестриктаза Вектор Плазмида Липкие концы Гомологичное спаривание Лигаза Репортерные гены Устойчивость к антибиотикам Репортерный ген LacZ, β-галактозидаза, Х-Gal Селективные среды Клеточные клоны Библиотека клонов Определение первичной последовательности ДНК 9 ДНК некоторых вирусов может встраиваться в геном хозяина. Вы исследуете структуру встроенной ДНК определенного вируса. Для этого вы получаете образцы ДНК свободного вируса и ДНК из клеток хозяина-носителя вируса. Эту ДНК вы гидролизуете рестриктазами, для которых вам известны сайты рестрикции (Рис. А). Далее вы разделяете полученные фрагменты при помощи электрофореза и визуализируете полосы (бэнды), соответствующие вирусной ДНК, при помощи Саузерн-блот гибридизации, используя в качестве зонда радиоактивно-меченную вирусную ДНК. Результат изображен на рис. В. Попробуйте определить, в каком из отмеченных участков (а-е) происходит разрыв кольца при интеграции вируса в геном хозяина. 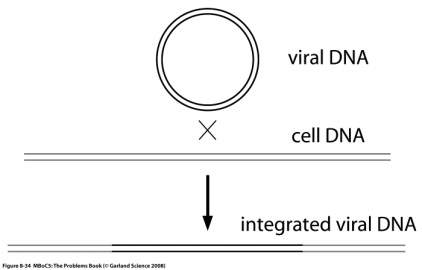 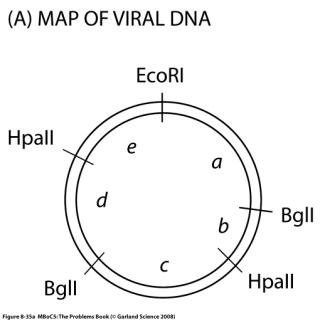 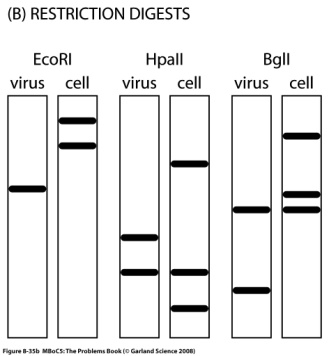 Контрольная работа № 3 Место образования субъединиц рибосом, наблюдаемое в световой микроскоп, называется ________________________(у эукариот) и ______________(у прокариот). Ферменты, называемые ______________, присоединяют каждую аминокислоту к соответствующей молекуле тРНК, образуя молекулу ________________. Генетический код называют ______________, потому что большинство аминокислот кодируется более чем одним кодоном. _____________ катализирует синтез РНК-копии на цепи ДНК в ходе процесса, называемого _____________. Субстратом для фермента_________________ являются дезоксирибонуклеозидтрифосфаты. Процесс транскрипции состоит из стадий: узнавание промотора, осуществляемое _______________, образование первой диэфирной связи или ____________, __________ и терминация. Для узнавания промотора необходим -фактор, который диссоциирует после ______________. _______ субъединица РНК-полимеразы обеспечивает прочное связывание с ДНК. В ___________ имеются два участка связывания молекулы тРНК: ___________, или Р-участок, удерживающий молекулу тРНК, присоединенную к растущему концу полипептидной цепи, и ______________ или А-участок, предназначенный для удерживания молекулы тРНК, нагруженной аминокислотой. Образование пептидной связи катализируется ферментом _________, каталитическая активность которой управляется крупной молекулой рРНК, входящей в состав ________ субъединицы рибосомы. Белки, называемые факторами __________, связываются со________-кодоном в А-участке рибосомы, в результате чего пептидилтрансфераза гидролизует связь, которая соединяет растущий пептид с молекулой тРНК. Во всех прокариотических клетках первую аминокислоту, с которой начинается любая белковая цепь, доставляет молекула особой _____________, узнающей кодон AUG и несущей аминокислоту _____________. …………………………………. Верны ли утверждения: Модифицированные нуклеотиды, особенно часто встречающиеся в молекулах тРНК, образуются в результате ковалентной модификации стандартных нуклеотидов перед их включением в РНК-транскрипты. Каждый комплекс аминокислоты с тРНК активирован не для его присоединения, а для присоединения очередной аминокислоты к растущей полипептидной цепи. Главная функция большой субъединицы рибосомы - связывание мРНК и различных тРНК; малая субъединица рибосомы катализирует образование пептидной связи. Поскольку стартовым кодоном для начала синтеза белка является AUG, то метионин обнаруживается только на N-концах полипептидных цепей белков. В любом месте двойной спирали ДНК обычно только одна цепь ДНК используется как матрица. В клетках бактерий транскрипцию РНК всех классов осуществляет РНК-полимераза одного типа, тогда как в клетках эукариот используются три разных типа РНК- полимераз. …………………………….. Схематически изобразите два противоположно направленных оперона E. coliи транскрипты с этих оперонов. Обозначьте кодирующие цепи, матричные цепи, их 5’ и 3' концы. Расскажите о регуляции Lac оперона E. Coli Перечислите основные этапы экспрессии про- и эукариотических генов. Примеры вопросов для коллоквиума Принципы строения двойной спирали ДНК. Структура промоторов у про- и эукариот Аттенуация Рекогниция. Аминоацилирование tРНК. Редактирование РНК Современная схема репликации ДНК E.coli Гибридизация нуклеиновых кислот. Саузерн-блот. Нозерн-блот. ПЦР. Основной принцип, примеры использования в молекулярной биологии и медицине Предположим, мы отсеквенировали геном нового организма. Какую информацию можно получить на основании только анализа этой последовательности ДНК (без дополнительных экспериментальных процедур), а так же сравнивая ее с уже известными последовательностями из международных банков данных? Свойства генетического кода Субъединичный состав РНК-полимеразы E.coli. Аттенуация Центры рибосом E.coli. Процессинг мРНК Секвенирование ДНК по Сэнгеру. Рестриктазы. Примеры использования рестриктаз в различных мол. биол. методах Предположим, мы отсеквенировали геном нового организма. Какую информацию можно получить на основании только анализа этой последовательности ДНК (без дополнительных экспериментальных процедур), а так же сравнивая ее с уже известными последовательностями из международных банков данных? |