Водоросли как сборная группа животных. Подходы к систематике
 Скачать 0.61 Mb. Скачать 0.61 Mb.
|

|
Водоросли как сборная группа животных. Подходы к систематике. Раньше водоросли являлись сборной группой организмов, «не подходящих» для других групп животных и растений. В них входили: вирусы; бактерии, разнесённые по двум разделам: большинство – в специально отведённый для них раздел, часть – в раздел «Водоросли»; водоросли; миксомицеты; грибы и лишайники. Сейчас благодаря методологическим и идейным изменениям систематика водорослей претерпела сильные изменения. В систематическом отношении водоросли делятся на множество самостоятельных отделов, различающихся по окраске, зависящей от набора пигментов, по организации клетки и структуре талломов. Для систематизирования водорослей используют: А) Внешние признаки, такие как окраска, структура таллома, внешний вид и т.д. Б) Филогенетический критерий: организмы объединяют на основе общности их происхождения, которое сейчас устанавливается главным образом на основе метода геносистематики (науки, целью которой является исследование генов изучаемых организмов и построение по сравнительным данным достоверных эволюционных деревьев) В) Структурно-морфологические признаки: например, наличие или отсутствие ядра, жгутиков и т.д., строение хлоропластов; благодаря данному признаку эукариотов делят на одноклеточные, многоклеточные, тканевые. Г) Эколого-трофический критерий: водоросли делят по типу питания (авто-, гетеротрофы, хемо-, фотосинтезирующие) Типы морфологической организации водорослей. Явление морфологического параллелизма. Вегетативное тело водорослей представлено слоевищем (талломом), не дифференцированным на органы. •Представляют одно-, много-, неклеточные (сифональные) и сифонокладиальные организмы, существующие одиночно или образуюшие разнообразные соединения (колонии) • Размеры тела колеблются от 1 мкм в диаметре (для цианобактерий) до нескольких десятков метров. • Форма: шаровидная, каплевидная, сложно расчлененная. • Окраска (11 цветов): зеленая, розовая, красная, оранжевая, фиолетовая, голубая, сине-зеленая, желто-зеленая, желтая, бурая. • Все разнообразие водорослей сводится к нескольким типам морфологической структур. Выделяют 11 типов арогенно возникших типов структур.  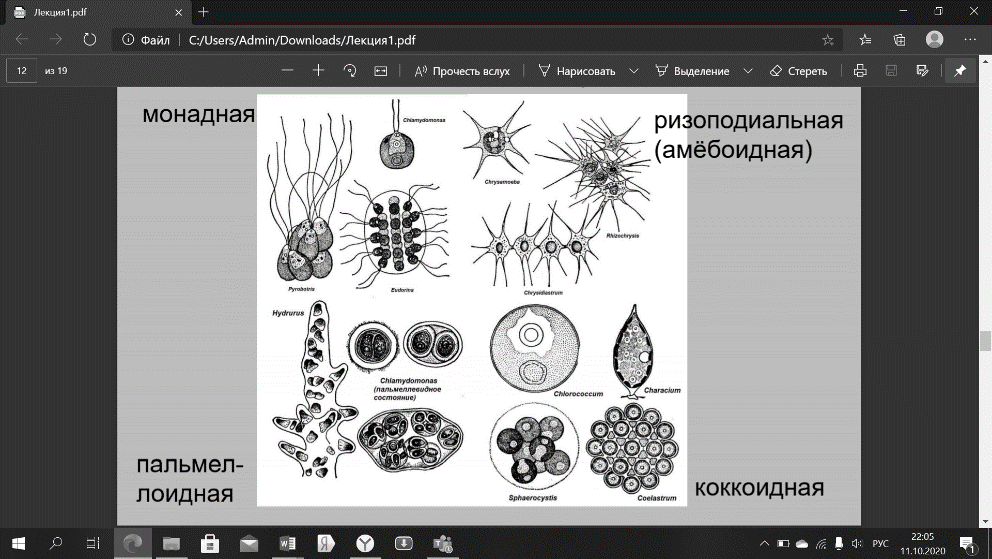 Монадный тип структуры Характерен для колониальных и одноклеточных представителей, способных в вегетативном состоянии активно передвигаться с помощью жгутиков. Доминируют среди эвгленовых, динофитовых, криптофитовых, рафидофитовых, золотистых, желтозеленых, зеленых водорослей. • Во время репродукции: у бурых и диатомовых. • Отсутствует полностью: у красных. • Монадные организмы способны осуществлять кинетические, векторные и фобические движения. Амебоидный тип (ризоподиальный) • Признаки: отсутствие прочных клеточных покровов и способность к амебному движению (т. е. с помощью временных цитоплазматических выростов - псевдоподий). Выделяют: ризоподии - длинные, тонкие, разветвленные; лобоподии - широкие, с закругленными концами; аксоподии – напоминают щупальца. Особенности клеток амебоидных • 1. Имеют ядра, пластиды, митохондрии и другие органеллы, характерные для эукариот. • 2. Паразитический тип питания. • 3. Хлоропласты: не проявляют признаков деградации или наблюдаются беспластидные формы. • 4. Могут вести прикрепленный образ жизни: - строить домик различной формы; - образовывать колонии со слизью и без неё 5. Соединение клеток в колониях происходит с помощью: - ризоподиев – сетчатые колонии (сетчатые плазмодии); - соединение амебоидов всей поверхностью – агрегированные плазмодии; - полное слияние амебоидов – слитный плазмодий. Распространение амебоидных • Среди золотистых, желто-зеленых, динофитовых. • Являются выражением крайней специализации эволюции, возникшей в результате приспособления монадных форм в биотопах, богатых органикой. Пальмеллоидный тип (гемимонадный) • Клетки, подобные монадным и имеющие характерные для монадного типа органеллы, но ведущие неподвижный образ жизни. Часто образуют колонии, состоящие из слизистых выделений и погружённых в эту слизь клеток. Коккоидный тип строения • Встречается у одноклеточных и колониальных представителей, неподвижных в вегетативном состоянии (за исключением десмидиевых и диатомовых), у которых отсутствуют жгутики и клетки всегда окружены клеточной стенкой. • Исходный тип для возникновения многоклеточных слоевищ, сифонального и сифонокладального типов структур. Трихальный (нитчатый) тип строения • Характеризуется расположением клеток в один или несколько рядов, образующихся в результате вегетативного деления клеток преимущественно в одной плоскости. Нити не ветвятся. Послужил отправным пунктом для развития других, более сложных типов структур. Гетеротрихальный тип строения • Нити ветвятся и способны к выполнению различных функций: опорной, прикрепительной, ассимиляторной и др. Ложнотканевой (псевдопаренхиматозный) тип строения • Клетки не являются сестринскими, цитоплазма не связана, клетки дифференцированы. Характеризуется образованием крупных объемистых слоевищ в результате срастания нитей. Представлен у бурых, красных и зеленых водорослей. Тканевой (паренхиматозный) тип строения • Образуется за счет того, что клетки способны делиться в трех плоскостях, образуя объемные слоевища с тканями, выполняющими различные функции. Характерен для бурых и частично зеленых и красных водорослей. • Закрепило господство водорослей в растительном мире водной среды. • Высшая ступень морфологической дифференциации тела водорослей. Сифональный (неклеточный) тип строения. Огромная многоядерная сильно расчленённая клетка без перегородок. Тупиковое направление, не выдержавшее конкуренции с магистральным эволюционным путем, возникшим на основе вегетативного деления клеток. Сифонокладиевый тип строения • Сифональная структура разбивается на отдельные многоядерные фрагменты. Только у зеленых водорослей. • Побочная, тупиковая ветвь морфологической эволюции растений. Морфологический параллелизм • Каждый из рассмотренных типов структуры вегетативного тела водорослей представлен в нескольких отделах. Идея впервые была изложена в работах А. Пашера (первая половина XX века): различно окрашенные водоросли, берущие начало от разных предков, проходили в эволюционном развитии сходные этапы – ступени морфологической дифференциации слоевища. Явление универсально: проявляется как на одном и том же структурно-эволюционном уровне в пределах одной ветви. Вместе с тем является помехой в классификации водорослей. Различные формы пластид у водорослей, пути эволюции Пластида – органелла, покрытая двумя мембранами, внутри которой находится матрикс с собственной системой репродукции. Пластиды характерны только для клеток фотосинтезирующих эукариот. В зависимости от строения и основных функций описаны разные типы пластид (например, хлоропласты, лейкопласты), основным из которых является хлоропласт. Хлоропласт (= хроматофор) – мембранная органелла, в которой происходят фотосинтетические процессы. Хлоропласт ограничен от цитоплазмы мембранами, внутри находится строма. В строме располагаются элементы авторепродукции хлоропластов (ДНК, РНК, рибосомы) и локализованы ферменты, участвующие в метаболических процессах. Внутри стромы имеются мембранные образования: тилакоиды – структуры, образованные за счет внутренней мембраны хлоропласта, которые могут формировать ламеллы – стопки тилакоидов, расположенные по всей длине хлоропласта или граны – стопки тилакоидов, в которых мембраны соседних тилакоидов сливаются. Хлоропласты многих водорослей содержат пиреноиды – участки хлоропласта, содержащие фермент темновой фазы фотосинтеза (рубиско). У некоторых водорослей, помимо хлоропластов, в клетках встречаются лейкопласты и их разновидность – амилопласты. Лейкопласт – бесцветная пластида, покрытая двумя мембранами, внутренняя из которых образует немногочисленные тилакоиды. В лейкопласте имеются ДНК, рибосомы и ферменты, осуществляющие синтез и гидролиз запасных веществ. Амилопласт – пластида из группы лейкопластов, в которой откладывается крахмал. Хлоропласты имеют симбиотическое происхождение. Согласно теории симбиогенеза, протопластидой была фотосинтезирующая цианобактерия, содержащая все необходимые для этого процесса компоненты. Пластиды Пигменты в клетках эукариотных водорослей находятся в пластидах, как и у всех растений. У водорослей встречается два типа пластид: окрашенные хлоропласты (хроматофоры) и бесцветные лейкопласты (амилопласты). Хлоропласты водорослей в отличие от таковых высших растений значительно разнообразнее по форме и по строению. У евгленовых и большинства динофитовых хлоропласт окружен тремя мембранами, а у гетероконтных и криптофитовых – четырьмя. УЧЕБНИК!!!!!!!!!!! Пигментные системы разных таксономических групп, особенности фотосинтеза Пигменты Все водоросли хорошо различаются по набору фотосинтезирующих пигментов. Такие группы в систематике растений имеют статус отделов. Основным пигментом всех водорослей является зеленый пигмент хлорофилл. Известно четыре типа хлорофилла, которые различаются по своей структуре: хлорофилл a – присутствует у всех водорослей и высших растений; хлорофилл b – встречается у зеленых, харовых, евгленовых водорослей и у высших растений: растения, содержащие этот хлорофилл, всегда имеют ярко-зеленую окраску; хлорофилл c – встречается у гетероконтных водорослей; хлорофилл d – редкая форма, встречается у красных и синезеленых водорослей. Большинство фотосинтезирующих растений содержат два разных хлорофилла, одним из которых всегда является хлорофилл a. В некоторых случаях вместо второго хлорофилла присутствуют билипротеины. У синезеленых и красных водорослей встречаются два вида билипротеинов: фикоцианин – синий пигмент, фикоэритрин – красный пигмент. Обязательными пигментами, входящими в фотосинтетические мембраны, являются желтые пигменты – каротиноиды. Они отличаются от хлорофиллов спектром поглощаемого света и, как полагают, выполняют защитную функцию, предохраняя молекулы хлорофилла от разрушительного воздействия молекулярного кислорода. Кроме перечисленных пигментов у водорослей встречаются: фукоксантин – золотистый пигмент; ксантофилл – бурый пигмент. Большинство водорослей являются фототрофами, которые осуществляют оксигенный фотосинтез (то есть с выделением кислорода) и используют в качестве донора электронов воду. Однако некоторые виды синезеленых водорослей способны обитать в анаэробных условиях и переходить на аноксигенный фотосинтез, а в качестве донора электронов использовать сероводород. Типы строения и состав клеточных покровов разных групп водорослей Клетка водорослей покрыта плазматической мембраной (═ плазмалеммой). В отличие от клеток высших растений, не у всех водорослевых клеток снаружи от плазмалеммы образуется клеточная стенка. В целом, строение клеточных покровов у водорослей довольно разнообразно 1) Клетка может быть покрыта только плазмалеммой и не иметь никаких дополнительных покровов. 2) Над плазматической мембраной могут формироваться дополнительные структуры. К таким структурам относятся, например, клеточная стенка, кремнеземный панцирь диатомей, различные слизистые чехлы и т.д. а) Клеточная стенка – это клеточный покров, секретируемый кнаружи от плазматической мембраны. Клетки многих водорослей имеют клеточную стенку. Набор веществ, слагающих клеточную стенку, различается у разных систематических групп водорослей. Так, например, основным компонентом клеточных стенок зеленых водорослей является целлюлоза, красных – целлюлоза и агароидные вещества и т.д. б) Кремнеземный панцирь – это клеточный покров, секретируемый кнаружи от плазматической мембраны и состоящий из кремнезема. Помимо аморфного кремнезема панцирь включает органические и неорганические соединения. Снаружи и изнутри панцирь покрыт слоем, состоящим из пектиновых веществ. Панцирь по строению напоминает коробку, сложенную донышком и крышкой, и состоит из двух половинок: большей – эпитеки («крышка») и меньшей гипотеки («донышко»). Такой тип покрова характерен для диатомовых водорослей. в) Слизистые чехлы располагаются на поверхности клетки и представлены различными полисахаридами. Слизи могут включать неорганические вещества (например, соли железа, марганца) и за счет этого быть окрашенными. Такие покровы характерны для цианобактерий и некоторых других водорослей. 3) Под плазматической мембраной могут формироваться дополнительные структуры. К таким структурам относится, например, покров эвгленовых водорослей, который называется пелликула. Пелликула – это клеточный покров, который включает в себя плазматическую мембрану и расположенные под ней белковые полосы и систему микротрубочек. Пелликула у некоторых видов эластична и может сокращаться, и растягиваться, поэтому клетки многих видов эвгленовых водорослей способны изменять свою форму. 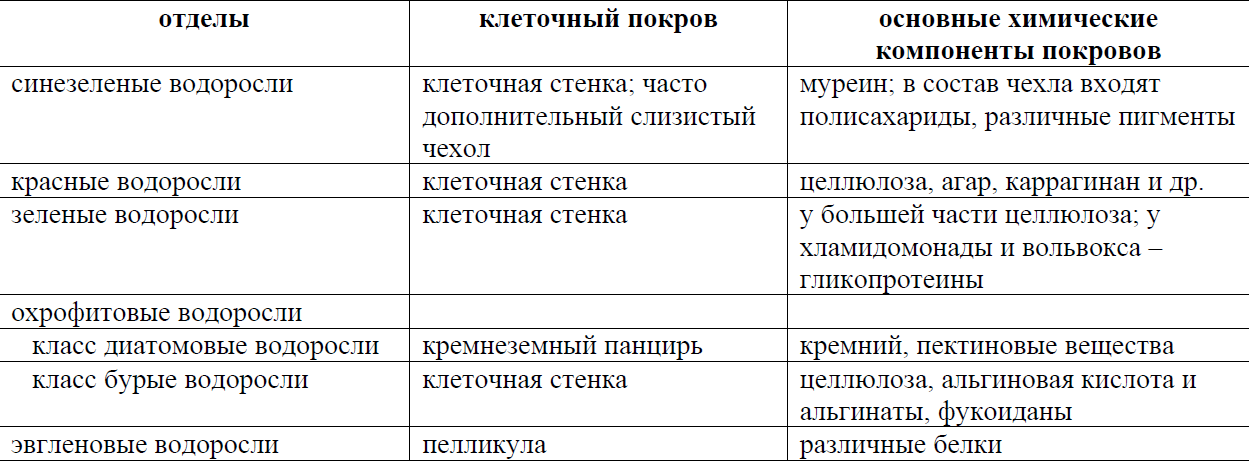 Движение водорослей. Двигаться к свету умеют многие растения. Водоросль хламидомонада, например, использует особую тактику, чтобы изменить направление, — начинает вращать жгутиками в разные стороны. Положительный фототаксис (так называется движение к источнику света) приводит туда, где можно фотосинтезировать. Род вольвокс — более высокоорганизованные родственники хламидомонады. Но если с движением хламидомонады все более или менее понятно, то над тем, как вольвокс движется к свету, ученые ломают голову уже более ста лет. Дело в том, что хламидомонада состоит всего из одной клетки. Поэтому ее движение достаточно простое. Вольвокс — многоклеточная водоросль. И жгутиков у нее как минимум 5 тыс. Чтобы такой организм двигался, вроде бы необходима координационная связь между клетками. Но такой связи у вольвокса нет. Что совершенно не мешает ему двигаться к свету. По словам ученых, каждая клетка вольвокса имеет два жгутика для движения и специальные «глазки» (или глазные пятна), которые воспринимают свет. «Вольвокс имеет форму сфероида. При помощи жгутиков он движется вперед и вращается вокруг своей оси против часовой стрелки, совершая один оборот в одну-три секунды», — рассказывает Уэки. Передняя и задняя часть вольвокса не одинаковы по своему строению. Вольвокс имеет, по словам ученых, два полюса. У одного (в передней части) находится множество глазных пятен. Свет воспринимают лишь они. К тому же постепенно количество пятен по направлению к противоположному полюсу уменьшается. Зафиксировав свет, передние клетки дают команду жгутикам двигаться и так задают направление. Эвглена двигается при помощи пелликулы. Некоторые двигаются при помощи слизи. Типы полового размножения, особенности в разных отделах. Способы полового размножения можно разделить на два типа: половой процесс без образования гамет и с образованием гамет. Клетка, в которой формируются половые клетки (гаметы), называется гаметангий. Вне зависимости от способа полового размножения, в результате слияния гаплоидных клеток (гамет) образуется диплоидная зигота, которая дает начало новому поколению. Различают следующие типы полового процесса. 1) Половой процесс без образования гамет. К такому типу полового процесса относится конъюгация – процесс слияния протопластов двух вегетативных клеток. Конъюгация характерна для некоторых зеленых водорослей, например, спирогиры. 2) Половой процесс с образованием гамет. К этому типу полового процесса относятся: а) изогамия – процесс слияния морфологически одинаковых и подвижных за счет жгутиков гамет; хламидомонада, улотрикс; б) анизогамия – слияние морфологически разных и подвижных за счет жгутиков гамет; при этом гамету большего размера называют условно «женской», меньшую – «мужской»; у некоторых зеленых и бурых водорослей в) оогамия – слияние неподвижной женской гаметы, которую называют яйцеклеткой, с мужской гаметой. Мужская гамета может иметь жгутики и в этом случае ее называют сперматозоид; если мужская гамета не имеет жгутиков, то ее называют спермаций. Женский гаметангий при этом называется оогоний, мужской – антеридий. бурых водорослей ламинарии и фукуса, а также красных водорослей. Не у всех водорослей в жизненном цикле известно половое размножение. Например, не описан половой процесс у таких водорослей, как синезеленые и эвгленовые. Жизненные циклы красных водорослей. Особенности в разных порядках. Мужские гаметы красных водорослей - спермации - развиваются по одному в органах размножения - сперматангиях. Спермации лишены хлорофилла. У некоторых красных водорослей спермации голые и способны к амебоидному движению. Женский репродуктивный орган называют карпогоном. Это особая клетка, густо заполненная протоплазмой и в большинстве случаев лишенная хлоропласта. После оплодотворения карпогона развиваются специальные нити - гонимобласты, которые несут карпоспоры. В жизненном цикле красных водорослей половое и бесполое размножение сменяют друг друга. Но обычно строгого чередования полового и бесполого поколений не происходит. В большинстве случаев гаметофит и спорофит могут воспроизводить себя в течение нескольких поколений. Органы полового и бесполого размножения, как правило, развиваются на разных растениях. Моно, би- и тетраспоры образуются на растениях-спорофитах, а оогонии и сперматангии - на гаметофитах. При этом у большинства красных водорослей существуют мужские и женские гаметофиты. Таким образом, половая и бесполая форма развития представлены обычно самостоятельными свободноживущими поколениями. Однако в ряде случаев наблюдается возникновение одной формы развития на другой, и тогда на одном и том же индивиде образуются и тетраспоры, и органы полового размножения. Гаметофит, спорофит, тетраспорофит. Кроме описанного цикла развития, в котором явно присутствует три формы развития, у красных водорослей существует другой, в котором имеется гаметофит с паразитирующим на нем карпоспорофитом- диплоидная бесполая фаза, а тетраспорофит настолько редуцирован, что ранее его считали отсутствующим. В таких случаях предполагали, что редукционное деление в этом цикле происходит сразу после оплодотворения - в самом начале прорастания зиготы - и весь цикл развития проходит в гаплоидной фазе. Однако недавние исследования показали, что единственное место, где происходит редукционное деление ядра - тетраспорофит. Таким образом, цикл развития красных водорослей может быть представлен двумя типами. Если гаметофиты и тетраспорофит имеют совершенно одинаковое строение, то такой цикл развития относят к типу Polysiphonia. Если слоевище тетраспорофита редуцировано по сравнению с гаметофитом и сильно отличается от него по строению (часто в таких случаях тетраспорофит представлен стелющимся или корковидным растением), то говорят о типе Porphyra. (У видов рода Porphyra Ag. пластинчатые крупные гаметофиты сменяются микроскопическими стелющимися спорофитами, построенными из однорядных нитей.) Редукция гаметофитов в цикле развития красных водорослей встречается гораздо реже. Обычно она выражается в недоразвитии гаметофита и прорастании его на тетраспорофите. Таким образом, в цикле развития красных водорослей происходит смена трех форм развития и существует смена ядерных фаз, но не всегда эти формы развиты одинаково и являются свободноживущими организмами. Жизненные циклы бурых водорослей. Особенности в разных порядках. Большинство бурых водорослей встречается в виде двух самостоятельных форм развития - спорофита и гаметофита (или гаметоспорофита), которые могут быть морфологически сходны или различны. Следовательно, для бурых водорослей характерны как изоморфная, так и гетероморфная смена форм развития. При гетероморфной смене форм развития одна из них бывает микроскопической. Чаще всего - это гаметофит. Развитию вертикальных макроскопических слоевищ бурых водорослей (в основном это спорофиты) нередко предшествует образование стелющихся нитей или дисков, - т. н. протонем. Если на протонеме развиваются органы размножения, а также со временем образуется вертикальное слоевище, то ее называют плетизмоталлюсом. Образование зооспор и гамет у бурых водорослей происходит во вместилищах двух типов - одногнездных и многогнездных. Многогнездные вместилища могут функционировать как спорангии и как гаметангии. Внешне они при этом не отличаются так же, как зооспоры и гаметы. Одногнездные вместилища чаще бывают спорангиями. У наиболее примитивных бурых (Puncturiariales, Chordariales, Ectocarpales) смена форм развития носит нестойкий характер. Из спор, производимых спорофитом, могут развиваться слоевища гаметофитов или гаметоспорофитов, или же слоевища спорофитов. Примером могут служить виды рода Ectocarpus Lyngb. Цикл развития видов этого рода может быть изоморфным или гетероморфным. При этом виды, считавшиеся самостоятельными, иногда оказываются формами развития одного и того же вида. Возникновение той или иной формы развития в значительной степени определяется экологическими условиями Жизненные циклы зелёных водорослей. Особенности в разных систематических группах. В жизненном цикле зеленых водорослей встречаются все основные типы смены форм развития. У них широко распространено как изоморфное, так и гетероморфное чередование поколений. Большинство зеленых водорослей в вегетативном состоянии гаплоидны, сравнительно небольшая часть диплоидна. Наиболее просто организованные зеленые водоросли порядков Polyblepharidales и Volvocales (Dunaliella salina Teod., Chlamydomonas proboscigera) характеризуются изоморфной сменой сравнительно простых форм развития. Для D. salina - типичного представителя порядка Polyblepharidales, половой процесс представляет собой типичную гологамию, в результате которой образуется диплоидная зигота. При прорастании зиготы происходит редукционное деление ядра, и вегетативные клетки D. salina имеют гаплоидный набор хромосом. В данном случае спорофит представлен одноклеточной зигоспорой, а вегетативные клетки являют собой гаметофиты. Таким образом, все поколения D. salina существуют в стадии гаметофита. Сходный цикл развития характерен и для Pyrobotrys gracilis Korsch. (Volvocales): 16-клеточные ценобии этой водоросли, вегетатирующие в течение всего теплого времени года, представляют собой гаметофиты; в конце вегетационного периода в клетках ценобиев вместо зооспор развиваются изогаметы; они попарно копулируют и дают зигоспору, которая переживает неблагоприятные условия до следующего года. Зигоспора - спорофит характерна и для холодолюбивого вида Stephanosphaera pluwialis Cohn (Volvocales). Она формируется в результате попарной копуляции изогамет, образующихся внутри клеток ценобия. В ценобии бывают от 4 до 32 изогамет. Зигоспора вегетирует на протяжении всего лета на поверхности тающего снега и льда в овражках и расщелинах скал каменистой тундры, накапливает гематохром, приобретает красную окраску и переходит в состояние покоя. Для более высокоорганизованных порядков зеленых водорослей характерна гетероморфная смена форм развития, в частности для некоторых Ulotrichales Циклы воспроизведения без чередования поколений, зависимость способа размножения от условий внешней среды. Циклы воспроизведения без чередования поколений (гаплобионта и диплобионта)(некоторые грибы и водоросли). 1) Гаплофазный цикл (диплоидна только зигота). Многие зеленые водоросли, в том числе улотрикс. 2) Диплофазный цикл (гаплоидны только гаметы). Немногие грибы и водоросли, в том числе фукус. I. Гаплофазный жизненный цикл без чередования поколений. Одна и та же особь производит или споры или гаметы. Вз R! Гаметы (+n, -n) ------ Зигота (n) ----- Взрослая особь (n) Характерные черты этого жизненного цикла: нет чередования поколений редукция зиготическая взрослая особь в гаплоидной фазе (гаплонт) Этот жизненный цикл характерен для сцеплянок, харовых водорослей, примитивных вольвоксовых , некоторых представителей хламидомонадовых, примитивных протококковых, одноклеточных представителей отдела желто-зеленые водоросли, улотрикса При неблагоприятных условиях в некоторых клетках водоросли образуются многочисленные мелкие подвижные гаметы со жгутиками. В воде они попарно сливаются, образуя зиготу. Обычно сливаются гаметы, возникшие в клетках нитей разных водорослей. Зигота покрывается толстой оболочкой и может долго находиться в состоянии покоя. При наступлении благоприятных условий зигота делится на 4 клетки — безжгутиковые споры. Каждая из них, опустившись на подводный предмет, может дать начало новой нитчатой водоросли улотриксу. Диплофазный жизненный цикл без чередования генераций R! Взрослая особь (2n) ---- гаметы (+n, -n) ---- зигота (2n) ---- взрослая особь (2n) Характерные особенности данного жизненного цикла: чередования поколений нет, отсутствует бесполое поколение редукция гаметическая Этот жизненный цикл встречается у животных, диатомовых водорослей, класса циклоспоровые из отдела бурые водоросли, порядка хлорококковые рода хлорохитриум, кладоферы (класс сифоновые из отдела зеленые водоросли). Бесполое размножение, типы спор. При бесполом размножении формируются специальные споры бесполого размножения. Клетка, в которой образуются споры, называется спорангий. У одних водорослей любая вегетативная клетка может стать спорангием, у других – только специализированная. Споры бесполого размножения водорослей можно классифицировать по разным признакам, например, способны споры к активному движению (то есть, имеют ли они жгутики) или нет; по типу деления, который предшествовал образованию спор и т.д. (таблица 10). 1) По способности к движению различают: а) апланоспоры – неподвижные споры и б) зооспоры – подвижные споры, которые двигаются за счет жгутиков. Клетка, в которой формируются зооспоры, называется зооспорангий. При бесполом размножении апланоспоры образуются, например, у хлореллы, всех красных водорослей; зооспоры – у хламидомонады и улотрикса. 2) По типу деления различают: а) митоспоры – перед образованием спор происходит митоз(–ы) и б) мейоспоры – первое деление, предшествующее образованию спор – мейоз. У хламидомонады, хлореллы, улотрикса при бесполом размножении образуются митоспоры, у ламинарии – мейоспоры. Не все водоросли способны к образованию спор бесполого размножения. Например, оно отсутствует у диатомовых и эвгленовых водорослей, а также у бурой водоросли фукус. Приспособление водорослей к планктонному образу жизни. Приспособительные черты в строении планктонных водорослей. Размеры клеток – это важная экологическая характеристика планктонных организмов. Чем больше клетка, тем отношение площадь ее поверхности к объему становится меньше, следовательно, тем тяжелее клетке плавать. Удельная масса у клеток водорослей больше, чем у воды, поэтому для обитания в толще воды необходимы определенные приспособления. С одной стороны, эти приспособления связаны с морфологией (наличие жгутиков, образование слизистых и плоских колоний, вытянутая форма клеток, мелкие размеры, различные выросты клеток и т.д.). С другой стороны, приспособления могут быть связаны с физиологией, например, клеточными включениями (капли масла и т.д.). Приспособление водорослей к бентосному образу жизни. Приспособительные черты в строении бентосных водорослей. Размеры бентосных организмов сильно варьируют, и, в отличие от планктона, многие бентосные формы являются макрофитами (их талломы могут достигать нескольких десятков метров в длину). Для обитания на дне водоема у бентосных, также как и у планктонных форм, существуют приспособления, связанные с морфологией и физиологией: крупные талломы крепятся ко дну с помощью ризоидов; микроскопические водоросли могут образовывать различные пленки или слизистые колонии и так далее |