Вопрос 1 Клеточная теория история и современное состояние. Значение клеточной теории для биологии и медицины
 Скачать 256.62 Kb. Скачать 256.62 Kb.
|

|
Вопрос 41 HLA-система. Методом хромосомной гибридизации установлено, что система HLA локализуется на коротком плече 6 аутосомной хромосомы человека. Размер комплекса HLA составляет 2 сантиморгана. Молекулярные классы HLA-региона – молекулы, кодируемые HLA-областью, разделены на три класса: I, II и III. Молекулы I класса – HLA-A, HLA-B и HLA-C – кодируются тремя отдельными парами генных локусов. Антигены I класса, впервые найденные на лейкоцитах (отсюда термин HLA), экспрессируются (синтезируются и выводятся на клеточную поверхность) почти во всех тканях (продукт четвертого локуса I класса, HLA-G, экспрессируется только в трофобласте). Молекулы I класса играют важную роль при распознавании антигена цитотоксическими T-клетками (CD8). Молекулы II класса кодируются тремя или более генными локусами (DR, DP и DQ). HLA-DR антигены известны также как Ia антигены по аналогии с антигенами иммунного ответа у мышей. Продуктами системы HLA являются антигены I и II классов. Антигены I класса являются мембранными гликопротеинами, состоящими из двух частей: глигозилированной полипептидной тяжелой цепи с молекулярной массой 44000 и В2-микроглобулина с молекулярной массой 12000. Молекулы HLA II класса также являются гликопротеинами, но состоят из двух нековалентно соединенных цепей А и В с молекулярной массой 34000 и 29000 соответственно.Антигены I класса содержатся почти на всех клетках органов и тканей организма, включая тромбоциты и стволовые гемопоэтические клетки. Антигены II класса имеют более ограниченное распространение, они выражены преимущественно на макрофагах, В-лимфоцитах, активированных Т-клетках и клетках-предшественниках гемопоэза. HLA выполняет в организме важные биологические функции. Первоначально полагали, что HLA имеет лишь непосредственное отношение к трансплантации органов и тканей. С помощью HLA-типирования удалось подтвердить общность некоторых расстройств или по-новому подойти к вопросу их классификации. Одни из них связаны с резистентностью или, наоборот, с восприимчивостью, а также со сроками возникновения болезней, другие с остротой их течения и, наконец, третьи – с продолжительностью жизни больных. Вопрос 42 Центральная догма молекулярной биологии. Центральная догма молекулярной биологии — обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении. Правило было сформулировано Френсисом Криком в 1958 году. 3 класса способов передачи информации, описываемые догмой Общие Специальные Неизвестные ДНК → ДНК РНК → ДНК белок → ДНК ДНК → РНК РНК → РНК белок → РНК РНК → белок ДНК → белок белок → белок Репликация ДНК (ДНК → ДНК) ДНК — основной способ передачи информации между поколениями живых организмов, поэтому точное удвоение (репликация) ДНК очень важна. Репликация осуществляется комплексом белков, которые расплетают хроматин, затем двойную спираль. После этого ДНК полимераза и ассоциированные с ней белки, строят на каждой из двух цепочек идентичную копию. Транскрипция (ДНК → РНК) Транскрипция — биологический процесс, в результате которого информация, содержащаяся в участке ДНК, копируется на синтезируемую молекулу мРНК. Транскрипцию осуществляют факторы транскрипции и РНК-полимераза. В эукариотической клетке первичный транскрипт (пре-иРНК) часто редактируется. Этот процесс называется сплайсингом. Трансляция (РНК → белок) Зрелая иРНК считывается рибосомами в процессе трансляции. В прокариотических клетках процесс транскрипции и трансляции не разделён пространственно, и эти процессы сопряжены. В эукариотических клетках место транскрипции — клеточное ядро — отделено от места трансляции (цитоплазмы) ядерной мембраной, поэтому иРНК транспортируется из ядра в цитоплазму. иРНК считывается рибосомой в виде трёхнуклеотидных «слов». Комплексы факторов инициации и факторов элонгации доставляют аминоацилированные транспортные РНК к комплексу иРНК-рибосома. Обратная транскрипция (РНК → ДНК) Обратная транскрипция — перенос информации с РНК на ДНК, процесс, обратный нормальной транскрипции, осуществляемый ферментом обратной транскриптазой. Встречается у ретровирусов, например, ВИЧ и в случае ретротранспозонов. Репликация РНК (РНК → РНК) Репликация РНК — копирование цепи РНК на комплемлементарную ей цепь РНК с помощью фермента РНК-зависимой РНК-полимеразы. Вирусы, содержащие одноцепочечную (например, пикорнавирусы, к которым относится вирус ящура) или двуцепочечную РНК реплицируются подобным способом. Прямая трансляция белка на матрице ДНК (ДНК → белок) Прямая трансляция была продемонстрирована в клеточных экстрактах кишечной палочки, которые содержали рибосомы, но не иРНК. Такие экстракты синтезировали белки с введённых в систему ДНК, и антибиотик неомицин усиливал этот эффект. Вопрос 43 Биосинтез белка. Биосинтез белка — сложный многостадийный процесс синтеза полипептидной цепи из аминокислот, происходящий на рибосомах с участием молекул мРНК и тРНК. Процесс биосинтеза белка требует значительных затрат энергии. (Процесс биосинтеза белка подразделяется на три этапа: Транскрипция,процессинг,трансляция. Транскрипция-процесс синтеза и-РНК происходит в ядре на одной из цепей ДНК,ферментативно. Процессинг-это созревание и-РНК в ядерном соке. Трансляция-синтез белка на и-РНК.) Вопрос 44 Транскрипция. Процессинг, сплайсинг. Отличие транскрипции у про- и эукариот. Процессинг РНК (посттранскрипционные модификации РНК) — совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта РНК в зрелую РНК. Наиболее известен процессинг матричных РНК, которые во время своего синтеза подвергаются модификациям: кэпированию, сплайсингу и полиаденилированию. Также модифицируются (другими механизмами) рибосомные РНК, транспортные РНК и малые ядерные РНК. Между транскрипцией и трансляцией молекула мРНК претерпевает ряд последовательных изменений, которые обеспечивают созревание функционирующей матрицы для синтеза полипептидной цепочки. К 5΄-концу присоединяется кэп, а к 3΄-концу поли-А хвост, который увеличивает длительность жизни иРНК. С появлением процессинга в эукариотической клетке стало возможно комбинирование экзонов гена для получения большего разнообразия белков, кодируемых единой последовательностью нуклеотидов ДНК, — альтернативный сплайсинг. Альтернативный сплайсинг — процесс, позволяющий одному гену производить несколько мРНК и, соответственно, белков. Большинство генов в эукариотических геномах содержат экзоны и интроны. После транскрипции в процессе сплайсинга интроны удаляются из пре-мРНК. А вот экзон может включаться (или нет) в состав конечного транскрипта. Таким образом, с помощью альтернативного сплайсинга можно получить множество транскриптов, а, следовательно, и белков. Объединение различных сайтов сплайсинга позволяет индивидуальным генам экспрессировать множество мРНК, которые кодируют белки, порой, с антагонистическими функциями. Экзон одного варианта сплайсинга может оказаться интроном в альтернативном пути. Разные варианты сплайсинга могут приводить к образованию разных изоформ одного и того же белка. Например, ген тропонина состоит из 18 экзонов и кодирует многочисленные изоформы этого мышечного белка. Разные изоформы тропонина образуются в разных тканях и на определенных стадиях их развития. РНК-полимераза прокариот обеспечивает транскрипцию генов, несущих информацию о последовательностях молекул РНК всех трех классов: рибосомной РНК (рРНК), транспорт-ной РНК (тРНК) и информационной (или матричной) РНК (мРНК). В эукариотических клетках имеются три различные РНК-по-лимеразы, каждая из которых специфически узнает промото-ры, контролирующие транскрипцию трех различных классов молекул РНК. РНК-полимераза I локализуется в ядрышке и синтезирует основные рибосомные РНК. РНК-полимераза III осуществляет транскрипцию транспортных РНК и одного ком-понента рибосом - 5SPHK. Транскрипция молекул мРНК, не-сущих информацию о структуре белков, осуществляется РНК-полимеразой II. Ферменты II и III типа локализуются в нуклеоплазме. Процессы образования мРНК в прокариотических и эукариотических клетках характеризуются существенными различиями. В эукариотических клетках после инициации транскрипции происходит модификация 5'-трифосфата в образующейся цепи за счет присоединения так называемого кэпа – метилированного остатка гуанозина. Кроме того, у большинства транскриптов происходит также модификация 3'-концов - по окончании транскрипции к ним присоединяется цепочка из остатков аденина, образующая характерный ро1уА-«хвост» (исключением из этого правила являются мРНК гистонных белков). У всех эукариот при транскрипции ДНК образуются молекулы РНК трех вышеназванных классов. Все они участвуют в процессе трансляции - третьей разновидности матричных процессов передачи информации - от РНК к белку. Вопрос 45 Трансляция - это процесс, в результате которого рибосомы считывают генетическую информацию матричных РНК и создают белковый продукт в соответствии с этой информацией. Специфические молекулы транспортных РНК (тРНК) служат посредниками между кодом мРНК и конечной белковой последовательностью. В их состав входит последовательность, узнающая код мРНК и соответствующая этому коду аминокислота. События трансляции разделяют на последующие события: инициацию, элонгацию и терминацию. На стадии инициации рибосома связывает мРНК и первая аминокислота присоединяется к рибосоме. Во время элонгации происходит рост полипептидной цепи. На стадии терминации рибосома отделяется от мРНК и процес трансляции заканчивается. У прокариот и эукариот процессы трансляции схожи, но имеются и существенные различия. Трансляция происходит в цитоплазме, где находятся рибосомы. В зависимости от дальнейшего преднозначения синтезируемых белков, они могут образовываться либо в цитозоле, либо на поверхности шероховатого эндоплазматического ретикулума.  Полипептидные цепи синтезируются однонаправленно: с амино-конца к карбокси-концу. 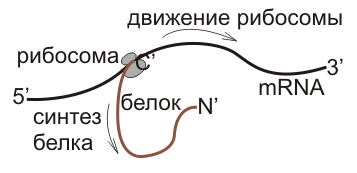 При инициации первая и вторая молекулы аминоацил-тРНК спариваются с первыми двумя кодонами мРНК. Далее трансляция продолжается в направлении 5'–>3' кодон за кодоном до тех пор, пока не достигнет стоп-сигнала, расположенного сразу же за кодоном, детерминирующим С-концевую аминокислоту. Модификация белков. Как правило, синтезированный полипептид подвергается дальнейшим химическим превращениям. Исходная молекула может разрезаться на отдельные фрагменты; затем одни фрагменты сшиваются, другие гидролизуются до аминокислот. Простые белки могут соединяться с самыми разнообразными веществами, образуя гликопротеины, липопротеины, металлопротеины, хромопротеины и другие сложные белки. Кроме того, аминокислоты уже в составе полипептида могут подвергаться химическим превращениям. Например, аминокислота пролин, входящая в состав белка проколлагена, окисляется до гидроксипролина. В результате из проколлагена образуется коллаген – основной белковый компонент соединительной ткани. Реакции модификации белков не являются реакциями матричного типа. Такие биохимические реакции называются ступенчатыми. Вопрос 46 Экспрессия генов — это процесс, в ходе которого наследственная информация от гена преобразуется в функциональный продукт — РНК или белок. Экспрессия генов может регулироваться на всех стадиях процесса: и во время транскрипции, и во время трансляции, и на стадии посттрансляционный модификаций белков. Регуляция экспрессии генов позволяет клеткам контролировать собственную структуру и функцию и является основой дифференцировки клеток, морфогенеза и адаптации. Экспрессия генов является субстратом для эволюционных изменений, так как контроль за временем, местом и количественными характеристиками экспрессии одного гена может иметь влияние на функции других генов в целом организме. На сегодняшний день известно более двухсот вариантов посттрансляционной модификации белков. Посттрансляционные модификации оказывают различные эффекты на белки: регулируют продолжительность их существования в клетке, ферментативную активность, взаимодействия с другими белками. Конститутивные гены – это гены с постоянной экспрессией, они постоянно включены, то есть функционируют на всех стадиях онтогенеза и во всех тканях. К конститутивным относятся гены, кодирующие тРНК, рРНК, ДНК-полимеразы, РНК-полимеразы, белки-гистоны, белки рибосом и т.д. Иначе говоря, это «гены домашнего хозяйства», без которых клетки не могут существовать. Ренулируемые гены – это гены с регулируемой экспрессией, они могут включаться и выключаться. У многоклеточных организмов индуцибельные гены их называют тканеспецифичными, потому что они по-разному функционируют в разных тканях на разных этапах онтогенеза. Регуляция активности генов осуществляется и на уровне транскрипции, и на уровне трансляции. Включение генов называется индукцией, а выключение – репрессией. Современная теория регуляции экспрессии генов у прокариот была предложена французскими исследователями Ф.Жакобом и Ж.Моно, которые исследовали биосинтез у E.сoli ферментов, метаболизирующих лактозу (β- галактозидазы, β-галактозидпермеазы и β-галактозидтрансацетилазы). Обнаружено, что при культивировании E.сoli на глюкозе содержание ферментов, метаболизирующих лактозу, минимально, но при замене глюкозы на лактозу происходит взрывоподобное усиление синтеза ферментов, разщепляющих лактозу на глюкозу и галактозу, и обеспечивают последующий метаболизм последних. У бактерий существуют ферменты 3-х типов: а) конститутивные, которые присутствуют в клетках в постоянных количествах, независимо от их метаболического состояния; б) индуцибельные – их количество в клетках при обычных условиях незначительно, но может увеличиваться в сотни и тысячи раз, если в культуральную среду добавлять субстраты этих ферментов; в) репрессабельные – ферменты, синтез которых в клетке прекращается при добавлении в среду конечных продуктов тех метаболических путей, где функционируют эти ферменты. На основании этих фактов и была сформулирована теория оперона. Оперон – это комплекс генетических элементов, отвечающих за координированный синтез ферментов, которые катализируют ряд последовательных реакций. В состав лактозного оперона E.сoli (Lac-оперон) входят: 1 структурные гены, которые кодируют первичную структуру ферментов, катализирующих последовательные реакции метаболизма лактозы 2. контрольные (регуляторные) сайты оперона, которые содержат промотор, оператор и терминатор. Промотор - это участок ДНК, с которым взаимодействует РНК-полимераза, а промотор Lac-оперона дополнительно содержит участок, с которым взаимодействует белок-активатор, контролирующий связывание РНК- полимеразы с промотором. Оператор - это участок ДНК, с которым специфически связывается белок-репрессор. Оператор непосредственно прилегает к структурным генам и связывание белка-репрессора с оператором противодействует считыванию РНК-полимеразой информации со структурных генов. Терминатор – это последовательность ДНК, которая распознается РНК- полмеразой как сигнал к прекращению транскрипции.Для функционирования оперона еще необходимо существование регуляторного гена, который экспрессирует регуляторный белок-репрессор. Этот ген структурно не связан с опероном и может находиться на значительном расстоянии от него. 1.Репрессия Lac- оперона. Синтез ферментов метаболиз- ма лактозы в клетках E.сoli заблокирован, поскольку связанный с оператором белок- репрессор противо- действует транскрип- ции генов этих ферментов. Среда содержит достаточ- ные количества глюкозы. 2.Индукция Lac- оперона. При снижении в среде уровня глюкозы и появлении лактозы гены, кодирующие ферменты обмена лактозы, разблокируются. Лактоза связывается с белком-репрессором и изменяет его конформацию. Репрессор теряет сродство к оператору и покидает его. Начинается транскрипцию генов метаболизма лактозы и синтез молекул этих ферментов. То есть лактоза выполняет функцию индуктора синтеза ферментов, которые катализируют ее собственный обмен. 3.Существует еще один вариант индукции Lac-оперона, благодаря существованию белка активатора катаболических генов. Этот белок после присоединения цАМФ стимулирует транскрипцию структурных генов, а уровень цАМФ возрастает в ответ на дефицит глюкозы. Метаболический принцип регуляции активности оперона; сродство белка- репрессора к оператору контролируется метаболитами цепи реакций, ферменты которой кодируются этим опероном. Различают индуцибельные опероны, активатор которых - исходный субстрат метаболического пути. При отсутствии субстрата белок-супрессор блокирует оператор и не дает РНК-полимеразе транскрибировать структурные гены. При появлении субстрата определенное его количество связывается с белком- репрессором, тот теряет сродство к оператору и покидает его. Это приводит к разблокированию транскрипции структурных генов. Репресабельные опероны – для них регулятором служит конечный метаболит. В его отсутствии белок- репрессор имеет низкое сродство к оператору и не мешает считыванию структурных генов (ген включен). При накоплении конечного метаболита, определенное его количество связывается с белком-репрессором, который приобретает повышенное сродство к оператору и блокирует транскрипцию генов. |