Вопрос 1. Структурно-химический состав и молекулярная организация плазмолеммы.
Плазмолемма – это поверхностная периферическая структура, не только ограничивающая клетку снаружи, но и обеспечивающая ее непосредственную связь с внеклеточной средой.
Основу плазмолеммы составляет липопротеиновый комплекс. Она имеет толщину около 10 нм. Основными химическими компонентами клеточных мембран, в частности плазмолеммы, являются липиды и белки. Снаружи от плазмолеммы располагается надмембранный слой – гликокаликс – ассоциированный с плазмолеммой гликопротеиновый комплекс, в состав которого входят углеводы. Эти углеводы имеют различные функции, а также придают мембране отрицательный заряд. Изнутри располагается кортикальный (подмембранный) комплекс, содержащий элементы цитоскелета.
К липидам плазматической мембраны относится большое количество гидрофобных веществ. Основные представители – фосфолипиды, холестерин, сфингомиелин (в основнов в цнс). Фосфолипиды располагаются в 2 стоя и имеют 2 части – гидрофильные головки(фосфатная часть), обращенные кнаружи в обе стороны и гидрофобные хвосты(ацильная часть), обращенные внутрь.
Мембранные белки также состоят из 2 частей – участков, богатых полярными аминокислотами и обращенных кнаружи, и гидрофобных участков, находящихся в гидрофобной части мембраны. Выделяют белки:
Интегральные 1. Белки-ферменты
Полуинтегральные 2. Белки-переносчики
Периферические 3. Рецепторные белки
4. Структурные белки
При этом интегральные выполняют: белки ионных каналов, рецепторные белки, а периферические: наружные (рецепторные и адгезионные), внутренние (цитоскелет – спектрин и анкирин).
Углеводы мембран входят в их состав не в свободном состоянии, они связаны с молекулами липидов или белков. Такие вещества называются соответственно гликолипидами и гликопротеидами.
Функции плазмолеммы: 1) распознавание других клеток и межклеточного вещества 2) движение клетки 3) транспорт веществ 4) взаимодействие с сигнальными молекулами.
Вопрос 2. Надмембранный (гликокаликс) и подмембранный (кортикальный) компоненты плазмолеммы. Особенности строения и функции. Мембранные рецепторы.
Гликокаликс представляет собой ассоциированный с плазмолеммой комплекс толщиной 50 нм, в состав которого входят различные углеводы. Углеводы образуют длинные, ветвящиеся цепочки полисахаридов, связанные с белками и липидами, входящими в состав плазмолеммы. Углеводные участки гликолииидов и гликонротеинов придают поверхности клетки отрицательный заряд.
Эти углеводные участки играют роль рецепторов (веществ, чувствительных к отдельным веществам), обеспечивающих распознавание клеткой соседних клеток и межклеточного вещества, а также адгезивные взаимодействия с ними. В состав гликокаликса некоторые авторы включают, помимо углеводных компонентов, периферические мембранные белки и полуинтегральные белки, функциональные участки которых находятся в надмембранной зоне (например, иммуноглобулины). В гликокаликсе находятся рецепторы гистосовместимости, некоторые ферменты, рецепторы гормонов. Такие чувствительные к отдельным веществам участки могут быть разбросаны по всей поверхности клетки или собраны в небольшие зоны. Выделяют рецепторы:
Ионотропные, которые при взаимодействии с субстратом образуют специфический ионный канал. Эти рецепторы обычно представляют собой сложный белок.
Метаботропные – обычно представлены гликопротеинами
Каталитические рецепторы включают внеклеточную часть (собственно рецептор) и цитоплазматическую часть, которая функционирует как протеинкиназа.
Рецепторы, связанные с G-белками - трансмембранные белки, ассоциированные с ионным каналом или ферментом, - состоят из рецептора, взаимодействующего с сигнальной молекулой (первый посредник), и G-белка, включающего несколько компонентов, который передает сигнал на связанный с мембраной фермент (аденилатциклазу) или ионный канал, вследствие чего активируется второй внутриклеточный посредник - чаще всего цАМФ или Са2+.
Функции гликокаликса: 1) Рецепторы 2)Узнавание 3)Взаимодействие 4)Гистосовместимость 5)Пристеночное переваривание
Кортикальный компонент образован специализированной периферической частью цитоплазмы, прилежащей к плазмолемме и содержащей элементы цитоскелета, преимущественно актиновые микрофиламенты. Более глубоко располагаются промежуточные филаменты и микротрубочки. Благодаря сокращению сети микрофиламентов, связанных с белками плазмолеммы, происходят изменения формы клетки и ее отдельных участков, формирование псевдоподий, выростов, перемещение клетки в пространстве. Также благодаря этому комплексу происходит перемещение белковых частиц, входящих в состав плазмолеммы с одного места на другое.
Вопрос 3. Молекулярная организация плазмолеммы. Пассивный и активный транспорт. Эндоцитоз и его разновидности.
Молекулярная организация плазмолеммы (см вопрос 1)
Пассивный транспорт включает — процессы, которые не требуют затраты энергии. Механизмом простой диффузии осуществляется перенос мелких молекул (например, О2, Н2О, СО2); этот процесс малоспецифичен и протекает со скоростью, пропорциональной градиенту концентрации транспортируемых молекул по обеим сторонам мембраны. Облегченная диффузия осуществляется через каналы и (или) белки—переносчики, которые обладают специфичностью в отношении транспортируемых молекул. В качестве ионных каналов выступают трансмембранные белки, образующие мелкие водные поры, через которые по электрохимическому градиенту транспортируются мелкие водорастворимые молекулы и ионы. Белки-переносчики также являются трансмембранными белками, которые претерпевают обратимые изменения конформации, обеспечивающие транспорт специфических молекул через плазмолемму. Они функционируют в механизмах как пассивного, так и активного транспорта.
Активный транспорт – перемещение веществ через мембрану против градиента концентрации с затратой энергии. Осуществляется за счёт специальных белков переносчиков. Выделяют:
Первично-активный транспорт, осуществляемый непосредственно за счёт использования энергии макроэргической связи АТФ. Например, Na-K насос.
Вторично-активный транспорт, осуществляемый за счёт ранее созданного с помощью первично-активного транспорта градиента концентрации. Например, перенос аминокислот внутрь клетки за счёт котранспорта с ионом натрия.
Макромолекулы, их агрегаты и крупные частицы попадают в клетку путём эндоцитоза. Эндоцитоз начинается с сорбции на поверхности плазмолеммы поглощаемых веществ. Связывание их с плазмолеммой определяется наличием на ее поверхности рецепторных молекул. После сорбции веществ на поверхности плазмолемма начинает образовывать сначала небольшие впячивания внутрь клетки. Эти впячивания могут иметь вид еще не замкнутых округлых пузырьков или представлять собой глубокие инвагинации, впячивания внутрь клетки. Затем такие локальные впячивания отшнуровываются от плазмолеммы и в виде пузырьков свободно располагаются под ней.
Часто для экономии (образования меньшего количества пузырьков) в ходе процесса эндоцитоза рецепторы определенных макромолекул перемещаются латерально по плазмолемме и накапливаются в окаймленной ямке. Благодаря белку клатрину, который образует сетевидную структуру в области этой ямки, вытесняются все остальные рецепторы из этой зоны. Через 1 минуту окаймленная ямка превращается в окаймленный пузырёк и теряет клатриновую оболочку, после чего происходит слияние пузырька с лизосомой и начинается процессинг лигандов, а рецепторы вторично используются в плазмолемме.
Эндоцитоз условно разделяют на:
Пиноцитоз – захват и поглощение клеткой жидкости и/или растворимых веществ.
Фагоцитоз – захват и поглощение клеткой плотных, обычно крупных частиц; обычно сопровождается образованием выпячиваний цитоплазмы - псевдоподий, охватывающих объект фагоцитоза и смыкающихся над ним.
Вопрос 4. Специализированные структуры плазмолеммы: микроворсинки, реснички, базальный лабиринт (СМ и ЭМ). Функции.
Микроворсинки – пальцевидные выросты цитоплазмы клетки диаметром 0.1 мкм и длиной 1 мкм (СМ), основу которых составляют актиновые микрофиламенты, располагающихся в кортикальном слое цитоплазмы. Каркас каждой микроворсинки образован пучком, содержащим около 40 микрофиламентов, лежащих вдоль ее длинной оси. В апикальной части микроворсинки этот пучок закреплен в аморфном веществе. Его жесткость обусловлена поперечными сшивками из белков фимбрина и виллина, изнутри пучок прикреплен к плазмолемме микроворсинки особыми белковыми мостиками. У основания микроворсинки микрофиламенты пучка вплетается в терминальную сеть, среди элементов которой имеются миозиновые филаменты. (ЭМ)
Микроворсинки обеспечивают многократное увеличение площади поверхности клетки, на которой происходит расщепление и всасывание веществ.
Реснички – органеллы специального значения, участвующие в процессах движения, - представляют собой выросты цитоплазмы, основу которых составляет аксонема (каркас из микротрубочек). Длина ресничек равна 2-10 мкм (ЭМ). Аксонема образована 9 периферическими парами микротрубочек (тубулин) и одной центрально расположенной парой; такое строение описывается формулой (9 х 2) + 2. В парах микротрубочки связаны белком динеином, который обладает АТФазной активностью (за счёт него происходит движение реснички). В основании аксонемы лежит базальное тельце, которое является матрицей для построения аксонемы. Состоит оно из девяти триплетов микротрубочек на периферии, в центре микротрубочек нет (9х3+0) (ЭМ). Функции:
Для подвижных клеток – перемещение, например, для сперматозоида
Для неподвижных клеток – перемещение жидкости или частиц, например, ресничные клетки ресничного эпителия (дыхательного) перемещают частицы пыли.
Базальный лабиринт –инвагинации плазмолеммы базального полюса клетки с лежащими между ними митохондриями. Эти митохондрии вырабатывают энергию, которая потребляется ионными насосами в их плазмолемме. Функция эпителиоцитов, обладающих базальной исчерченностью, связана с изменением ионного состава жидкости (мочи, слюны) в просвете указанных канальцев и протоков. Форму базального лабиринта поддерживают структуры цитоскелета: промежуточные филаменты и тонкие микрофиламенты.
Вопрос 5. Структура и типы рибосом (ЭМ, химический состав, гистохимическая характеристика). Полисомы. Синтез цитоплазматических белков на свободных полисомах.
Рибосомы – мелкие (диаметр - 15-30 нм) немембранные органеллы, обеспечивающие синтез белка путем соединения аминокислот в попипептидные цепочки. Они представляют собой сложные рибонуклеопротеиды (белки и рРНК примерно одинаковы по массе). Рибосома состоит из большой и малой субъединиц и содержит 4 активных центра:
М-центр (на малой субъединице) – связывает мРНК и передвигает рибосому по ней
П-центр (на обеих субъединицах) – связывает метионин-тРНК (инициация) а затем пептидил-тРНК (синтезиуемый белок с одной тРНК, которая ещё не отщипилась)
А-центр (на обеих субъединицах) – связывает аа-тРНК
ПТФ-центр (на большой субъединице) – катализируюет перенос аминокислоты из А-центра в П-центр, а также отсоединение тРНК от пептидил-тРНК в П-центре
Рибосомы различают свободные, которые находятся в цитоплазме в разобранном состоянии и не способны участвовать в процессе трансляции, и функционирующие, которые связаны с мембраной грЭПС. Рибосомы могут располагаться единично или в комплексах (полисомы).
Синтез белка рибосомой начинается со связывания малой субъединицы с участком иРНК; далее рибосома передвигается вдоль цепи иРНК, причем на каждом этапе происходит специфическое присоединение к рибосоме молекулы аа-тРНК, антикодон которой комплементарен соответствующему кодону иРНК. В полипептид включается около 20 аминокислот в 1 секунду; белковая молекула среднего размера синтезируется за 20-60 с. Когда образование белковой цепочки завершается, субъединицы диссоциируют, освобождаясь от иРНК. Пока продолжается синтез белка данной рибосомой, новая рибосома занимает освобождающееся на иРНК место. По этой причине активно транслируемая иРНК находится в полисомах.
Из Могильной (так проще запомнить):
Синтез сигнального пептида на полисомах
Связывание сигнального пептида с сигналраспознающей частицей (СРЧ)
Прекращение синтеза белка
Взаимодействие СРЧ со спецефическим рецептором (якорный белок) на мембране ЭПС
Удаление СРЧ от полисом
Разблокирование синтеза белка
Присоединение большой субъединицы рибосом за счёт рибофорина
Поступление синтезируемого полипептида в полость ЭПС
Отщепление сигнального пептида от полипептидной цепи
Посттранялсляционная модификация.
Белки, которые после синтеза остаются в гиалоплазме и далее используются ею, обычно синтезируются на свободных полисомах. Полисомы, которые своими большими субъединицами прикреплены к мембранам ЭПС, синтезируют белки, накапливающиеся в просвете цистерн ЭПС и в дальнейшем либо секретируемые клеткой, либо запасаемые ею внутри гранул (например, лизосомальные ферменты). На полисомах, связанных с мембранами ЭПС, синтезируется также большая часть интегральных мембранных белков. Будет ли белок синтезироваться на ЭПС или на свободных полисомах, зависит от характера начально образуемого отдела полипептидной цепи (сигнальной последовательности или пептида).
Вопрос 6. Эндоплазматическая сеть. Строение, разновидности ЭПС. Структура гранулярной и агранулярной эндоплазматической сети (СМ,ЭМ) и их функции.
Эндоплазматическая сеть (ЭПС) - органелла, обеспечивающая синтез углеводов, липидов и белков, а также начальные посттрансляционные изменения последних. Она имеет мембранное строение и состоит из системы уплощенных и удлиненных цистерн, трубочек и везикул, образующих 3-мерную сеть.
Выделяют две разновидности ЭПС: гранулярную ЭПС (грЭПС) и агранулярную ЭПС (аЭПС), которые связаны друг с другом переходной ЭПС. В области переходной ЭПС трубочки распадаются на отдельные фрагменты, образующие окаймленные транспортные пузырьки, которые переносят материал из ЭПС в комплекс Гольджи. Развитие ЭПС и преобладание одного из видов зависит от типа клетки и её функции.
Гранулярная ЭПС обеспечивает (1) биосинтез всех мембранных белков и белков, предназначенных для экспорта из клетки, и (2) начальные этапы модификации белковых молекул. Гранулярная ЭПС образована уплощенными мембранными цистернами и трубочками, на наружной поверхности которых располагаются рибосомы и полисомы. Мембраны грЭПС содержат особые белки (рибофорины), которые обеспечивают связывание рибосом и уплощение цистерн. Полость грЭПС содержит продукты синтеза и сообщается с перинуклеарным пространством. Благодаря грЭПС происходит отделение (изоляция) вновь синтезированных белковых молекул от гиалоплазмы.
Агранулярная (гладкая) ЭПС представляет собой трехмерную замкнутую сеть мембранных анастомозирующих трубочек, канальцев, цистерн и пузырьков диаметром 20-100 нм. Функции:
Синтез липидов, в том числе мембранных
Синтез гликогена
Синтез холестерина
Детоксикация эндогенных и экзогенных веществ
Накопление ионов Са2+
Восстановление кариолеммы в телофазе митоза.
Вопрос 7. Комплекс Гольджи, (СМ и ЭМ). Полярность комплекса Гольджи. Особенности процессинга молекул и направленный транспорт веществ.
Комплекс Гольджи - сложно организованная мембранная органелла, образованная тремя основными элементами, комплекс которых называется диктиосома:
Цистерны – имеют вид изогнутых дисков 0.5 – 5 мкм и образуют стопку из 3-30 элементов. Выпуклой стороной стопка обычно обращена к ядру, вогнутой - к плазмолемме. Каждая группа цистерн внутри стопки отличается особым составом ферментов, определяющим характер реакций процессинга белков. Периферические отделы цистерн несколько расширены, от них отщепляются пузырьки и вакуоли.
Пузырьки - сферические окруженные мембраной элементы диаметром 40-80 нм с содержимым умеренной плотности; образуются путем отщепления от цистерн.
Вакуоли - крупные, окруженные мембраной сферические образования, отделяющиеся от цистерны на зрелой поверхности комплекса Гольджи. Они содержат секреторный продукт умеренной плотности.
В специализированных секреторных клетках комплекс Гольджи располагается надъядерно под апикальной частью клетки, через которую происходит выделение секрета механизмом экзоцитоза. Нередко он лежит у ядра вблизи центриолей, в некоторых клетках его компоненты рассеяны по всей цитоплазме.
Полярность комплекса Гольджи. Комплекс Гольджи представляет собой поляризованную структуру, в которой выделяют две поверхности, обладающие структурными и функциональными различиями:
Цис-поверхность – незрелую, обращенную к ЭПС и связанную с системой мелких пузырьков, отщепляющихся от ЭПС.
Транс-поверхность – зрелую, обращенную к плазмолемме и связанную с отделяющимися от цистерн вакуолями
Функции комплекса Гольджи:
Синтез полисахаридов и гликопротеинов
Процессинг молекул
Конденсация секреторного продукта и образование секреторных гранул
Обеспечение новообразованных гранул мембраной
Сортировка белков на транс-поверхности перед их окончательным транспортом
Особенности процессинга молекул. В ходе процессинга в КГ происходит включение углеводных компонентов в гликопротеины (терминальное гликозилирование), добавление фосфатных групп (фосфорилирование), жирных кислот (ацилирование), сульфатных остатков (сульфатирование), частичное расщепление белковых молекул (протеолитическая доработка).
Каждый их указанных этапов процессинга веществ внутри комплекса Гольджи осуществляется в топографически определенном его компоненте (цис-, медиальных или транс-цистернах).
Транспорт белков из КГ тесно связан с их сортировкой. Сортировка производится посредством специфических мембранных рецепторных белков, которые распознают сигнальные участки на макромолекулах и направляют их в соответствующие пузырьки. Транспорт белков из комплекса Гольджи осуществляется в составе трех важнейших потоков:
В гидролазные пузырьки (первичные лизосомы)
В плазмолемму
В секреторные гранулы
Вопрос 8. Структура и функции эндосом и лизосом. Типы эндосом и лизосом.
Лизосомы – это разнообразный класс вакуолей размером 0,2 – 0,4 мкм, ограниченных мембраной, являются производным КГ. Характерным признаком лизосом является наличие в них гидролитических ферментов – гидролаз, расщепляющих различные биополимеры при кислом рН. Основная функция – внутриклеточное переваривание.
Эндосомы - мембранные пузырьки с постепенно закисляющимся содержимым, которые обеспечивают перенос макромолекул с поверхности клетки в лизосомы и их частичный или полный гидролиз на стадиях, предшествующих лизосомальному уровню деградации. Основные функции – транспорт веществ и начальное внутриклеточное переваривание.
Лизосомы и эндосомы представляют собой компартменты (то есть обособленные от цитоплазмы участки с собственной средой для протекания определенных реакций). Создание кислой среды в этих пузырьках осуществляется за счёт большого количества АТФазных протонных насосов в их мембране. Также мембрана:
Содержит рецепторы, обуславливающие её связывание с мембраной гидролазных и транспортных пузырьков, а также фагосом
Обеспечивает свободную диффузию низкомолекулярных продуктов переваривания в гиалоплазму
В неповрежденном состоянии представляет собой барьер, резистентный к действию литических ферментов и препятствующий их утечке в гиалоплазму.
Типы эндосом:
Ранние (периферические) эндосомы – являются мембранными пузырьками на ранних этапах после их отделения от плазмолеммы (но уже после утраты первоначально имевшейся клатриновой оболочки). Они располагаются неподалеку от плазмолеммы в периферических отделах цитоплазмы. В них происходит:
В условиях слабокислой среды (pH 6.0) осуществляется ограниченное и регулируемое переваривание макромолекул протеазами
Отщепление лигандов от рецепторов с их сортировкой (сортировкой рецепторов) и возможным возвращением последних в специальных пузырьках в плазмолемму для повторного цикла их использования.
расщепление комплексов рецептор-гормон, рецептор-фактор роста, антиген-антитело, а также ограниченный протеолиз (процессинг) антигенов (например, в макрофагах)
Поздние (перинуклеарные) эндосомы (800 нм) – образуются из ранних и располагаются в глубоких отделах цитоплазмы вблизи ядра. Их отличает от ранних эндосом более плотный матрикс, более кислое содержимое (pH 5.5) и более глубокий уровень переваривания ферментами.
Типы лизосом:
Гидролазные пузырьки – округлые мембранные органеллы диаметром до 200-400 нм с мелкозернистым плотным матриксом, содержащие литические ферменты в неактивной форме. Их перемещение в цитоплазме контролируется микротрубочками. Гидролазные пузырьки участвуют в транспорте литических ферментов в эндоцитозный путь из КГ. В настоящее время известно около 60 литических ферментов; все они представляют собой кислые гидролазы с оптимумом рН 5.
Лизосомы – органеллы, активно участвующие в завершающих этапах процесса внутриклеточного переваривания захваченных клеткой макромолекул посредством широкого спектра литических ферментов при низких значениях pH. Их разделяют в зависимости от наличия в их просвете морфологически распознаваемого материала:
Фаголизосома формируется путем слияния поздней эндосомы или лизосомы с фагосомой.
Аутофаголизосома образуется при слиянии поздней эндосомы или лизосомы с аутофагосомой
Мулътивезикулярное тельце – крупная (800 нм) сферическая окруженная мембраной вакуоль, содержащая мелкие пузырьки (40 нм). причем мелкие пузырьки формируются, вероятно, путем отпочкования внутрь от мембраны вакуоли. Матрикс тельца содержит литические фермента и, очевидно, обеспечивает постепенное разрушение внутренних пузырьков.
Остаточные тельца - лизосомы, содержащие непереваренный материал, которые могут длительно находиться в цитоплазме или выделять свое содержимое за пределы клетки.
Вопрос 9. Митохондрии (СМ и ЭМ). Функции митохондрий.
Митохондрии представляют собой двумембранные полуавтономные органеллы, обеспечивающие клетку энергией, получаемой благодаря процессам окисления и запасаемой в виде фосфатных связей АТФ. Митохондрии также участвуют в биосинтезе стероидов, окислении жирных кислот и синтезе нуклеиновых кислот (функции)
Митохондрии могут иметь эллиптическую, сферическую, палочковидную, нитевидную формы. В цитоплазме митохондрии могут располагаться диффузно, однако обычно они сосредоточены в участках максимального потребления энергии, например, вблизи ионных насосов, сократимых элементов (миофибрилл), органелл движения (аксонем спермия, ресничек), компонентов синтетического аппарата (цистерн ЭПС).
Митохондрии состоят из наружной и внутренней мембран, разделенных межмембранным пространством, и содержат митохондриальный матрикс, в который обращены складки внутренней мембраны – кристы.
Наружная митохондриальная мембрана напоминает плазмолемму. Она содержит много молекул специализированных транспортных белков (например, порин), которые формируют широкие гидрофильные каналы и обеспечивают ее высокую проницаемость, а также небольшое количество ферментных систем. На ней находятся рецепторы, распознающие белки, которые переносятся через обе митохондриальные мембраны в особых точках их контакта – зонах слипания.
Внутренняя митохондриальная мембрана отделена от наружной межмембранным пространством. Низкая проницаемость внутренней мембраны для мелких ионов имеет большое значение для функции митохондрий, так как она обеспечивает возможность создания электрохимического градиента. В состав мембраны входят:
Транспортные белки
Ферменты дыхательной цепи и сукцинатдегидрогеназаназа
Комплекс АТФ-синтетазы
Кристы - складки внутренней мембраны толщиной 20 нм; располагаются чаще всего перпендикулярно длиннику митохондрии, но могут лежать и продольно. Их число и площадь пропорциональны активности митохондрии. На них происходит сопряжение процессов окисления и фосфорилирования. В области округлой головки F1-частицы осуществляется синтез АТФ из АДФ.
Митохондриальный матрикс - гомогенное мелкозернистое вещество умеренной плотности, заполняющее полость митохондрии. В нём содержится несколько сотен различных ферментов, участвующих в ЦТК, окислении ЖК, ферментов белкового синтеза. В матриксе находятся также митохондриальные рибосомы, митохондриальные гранулы и митохондриальная ДНК.
Функции митохондрий: 1) Обеспечение клетки энергии АТФ 2) Участие в синтезе стероидных гормонов 3) Участие в синтезе нуклеиновых кислот 4) Депонирование кальция
Вопрос 10. Цитоскелет. Компоненты цитоскелета. Строение, ЭМ, химичиский состав, функции.
Цитоскелет – сложная динамичная система микротрубочек, микрофиламентов, промежуточных филаментов и микротрабекул. Компоненты цитоскелета являются немембранными органеллами, и каждый из компонентов образует 3мерную сеть, которая взаимодействует с сетями из других компонентов. Они входят также в состав ряда других более сложно организованных органелл (ресничек, жгутиков, микроворсинок, клеточного центра) и клеточных соединений (десмосом, полудесмосом, опоясывающих десмосом).
1)Микротрубочки - наиболее крупные компоненты цитоскелета. Они представляют собой полые цилиндрические образования, имеющие форму трубочек, длиной до нескольких микрометров.
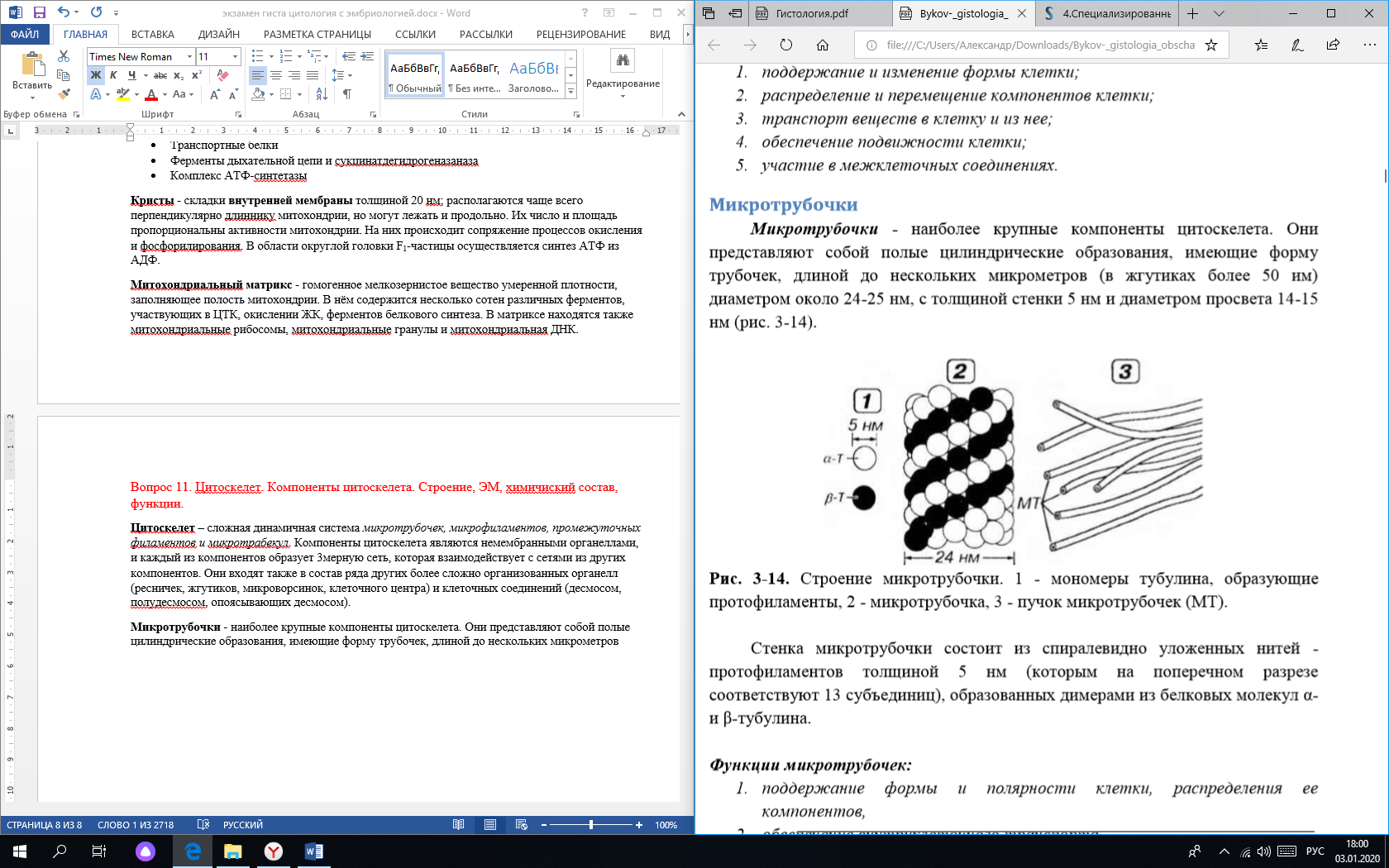
Стенка микротрубочки состоит из спиралевидно уложенных нитей - протофиламентов(1) толщиной 5 нм, образованных димерами из белковых молекул α- и β-тубулина.
Функции микротрубочек:
поддержание формы и полярности клетки, распределения ее компонентов
обеспечение внутриклеточного транспорта
обеспечение движения хромосом в митозе (образуют веретено деления)
образование основы других органелл (центриолей, ресничек, жгутиков)
Реснички (см вопрос 4)
Клеточный центр – немембранная органелла, образованная двумя полыми цилиндрическими структурами – центриолями, состоящими из 9 триплетов микротрубочек по периферии и полым центром. Они располагаются вблизи ядра во взаимно перпендикулярных плоскостях. При делении в клетке центриоли служат основой для формирования веретена деления, а их количество удваивается.
2)Микрофиламенты – тонкие белковые нити диаметром 5-7 нм, лежащие в цитоплазме поодиночке, в виде сетей ими пучками. Кортикальная (терминальная) сеть - зона сгущения микрофиламентов под плазмолеммой, характерная для большинства клеток. В этой сети микрофиламенты переплетены между собой и "сшиты" друг с другом с помощью особых белков, самым распространенным из которых является филамин. Кортикальная сеть препятствует резкой и внезапной деформации клетки при механических воздействиях и обеспечивает плавные изменения ее формы путем перестройки. Микрофиламенты более устойчивы к физическим и химическим воздействиям, чем микротрубочки.
Актин - основной белок микрофиламентов - встречается в моно-мерной форме (G-, или глобулярный актин), которая способна полимеризоваться в длинные цепи (F-, или фибриллярный актин)
Функции мирофиламентов:
Обеспечение функций, связанных с кортикальным слоем
Перемещение внутри цитоплазмы органелл
Обеспечение определенной жесткости клетки за счет наличия кортикальной сети, которая препятствует действию деформаций, но сама, перестраиваясь, способствует изменениям клеточной формы
Формирование сократимой перетяжки при цитотомии, завершающей клеточное деление
Образование основы микроворсинок
Участие в организации структуры межклеточных соединений
Обеспечение сократимости мышечных клеток (при взаимодействии с миозином)
3)Промежуточные филаменты - прочные и устойчивые в химическом отношении белковые нити толщиной около 10 нм (что является промежуточным значением между толщиной микротрубочек и микрофиламентов). Они встречаются в клетках разных тканей и располагаются в виде трехмерных сетей в различных участках цитоплазмы, окружают ядро, входят в состав десмосом и полудесмосом эпителиальных клеток, лежат по всей длине отростков нейронов. Промежуточные филаменты образованы нитевидными белковыми молекулами, сплетенными друг с другом наподобие каната. Функции:
Структурная
Обеспечение равномерного распределения сил деформации
Участие в образовании рогового вещества в эпителии кожи
Поддержание формы отростков нервных клеток и фиксация трансмембраниых белков
Удержание миофибрилл в мышечной ткани и прикрепление их к плазмолемме
4)Микротрабекулы - наименее изученная система цитоскелета, само существование которой оспаривается многими исследователями. Предполагают, что гри описанные выше системы филаментов пронизываются и объединяются некоей четвертой системой, названной микротрабекулярной сетью.
Вопрос 11. Ядро. Понятие об интерфазном ядре. Структурные компоненты ядра (СМ,ЭМ). Значение и функции ядра в жизнедеятельности клетки.
Ядро является важнейшим компонентом клетки, содержащим ее генетический аппарат. Функциями ядра являются хранение, реализация и передача генетического материала при делении клетки.
Обычно в клетке имеется только одно ядро, однако встречаются многоядерные клетки, которые образуются вследствие деления клеток, не сопровождающегося цитотомией, или слияния нескольких одноядерных клеток
Интерфазное ядро— ядро клетки в период интерфазы, когда оно не претерпевает митотических делений, а выполняет лишь свои специфические рабочие функции по синтезу полипептидов и белка.
Структурные компоненты ядра. В ядре неделящейся (интерфазной) клетки выявляются кариолемма (ядерная оболочка), хроматин, ядрышко и кариоплазма (ядерный сок).
1) Ядерная оболочка (кариолемма) на светооптическом уровне практически не определяется; под электронным микроскопом обнаруживается, что она состоит из двух мембран - наружной и внутренней, - разделенных полостью шириной 15-40 нм (перинуклеарным пространством) и смыкающихся в области ядерных пор.
Наружная мембрана составляет единое целое с мембранами грЭПС - на ее поверхности имеются рибосомы, а перинуклеарное пространство соответствует полости цистерн грЭПС и может содержать синтезированный материал. Со стороны цитоплазмы окружена виментиновыми филаментами
Внутренняя мембрана - гладкая, ее интегральные белки связаны с ядерной пластинкой - ламиной - слоем толщиной 80-300 нм, состоящим из переплетенных промежуточных филаментов (ламинов), образующих кариоскелет. Ламина играет очень важную роль в:
поддержании формы ядра
упорядоченной укладке хроматина
структурной организации поровых комплексов
Ядерные поры — это специализированные коммуникации (от 3-4 тыс.), осуществляющие транспорт веществ между ядром и цитозолем. Ядерная пора диаметром 80 нм имеет следующее:
канал поры – 9 нм
комплекс ядерной поры содержит белок-рецептор, реагирующий на сигналы ядерного импорта (входной билет в ядро). Диаметр ядерной поры может увеличивать диаметр канала поры и обеспечивать перенос в ядро больших макромолекул (ДНК-РНК-полимераза)
Ядерная пора состоит из 2-х параллельных колец по одному с каждой поверхности кариолеммы. Кольцо диаметром 80 нм, образовано 8 гранулами, от каждой гранулы к центру тянется нить (5 нм), которая формирует перегородку (диафрагму). В центре расположена центральная гранула. Совокупность этих структур называется комплекс ядерной поры. Здесь формируется канал диаметром 9 нм, такой канал называют водным, поскольку по нему движутся мелкие водорастворимые молекулы и ионы.
Функции ядерной поры:
Избирательный транспорт
Активный перенос в ядро белков с последовательностью, характерной для белков ядерной локализации
Перенос в цитоплазму субъединиц рибосом с изменением конформации порового комплекса
2) Хроматин – мелкие зернышки и глыбки материала, который обнаруживается в ядре клеток и окрашивается основными красителями. Хроматин состоит из комплекса ДНК и белка и соответствует хромосомам, которые в интерфазном ядре представлены длинными, тонкими перекрученными нитями и неразличимы как индивидуальные структуры. Различают два вида хроматина:
Эухроматин соответствует сегментам хромосом, которые деспирализованы и открыты для транскрипции. Эти сегменты не окрашиваются и не видны в световой микроскоп.
Гетерохроматин соответствует конденсированным, плотно скрученным сегментам хромосом (что делает их недоступными для транскрипции). Он интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид гранул.
3) Ядрышко – образовано специализированными участками (петлями) хромосом, которые называются ядрышковыми организаторами. У человека такие участки имеются в пяти хромосомах - 13-й, 14-й, 15-й, 21-й и 22-й, где располагаются многочисленные копии генов, кодирующих рРНК. Функции ядрышка заключаются в синтезе рРНК и ее сборке в предшественники рибосомальных субъединиц.
Ядрышко выявляется в интерфазном ядре на светооптическом уровне как мелкая плотная гранула диаметром 1-3 мкм, интенсивно окрашивающаяся основными красителями. Оно располагается в центре ядра или эксцентрично. Размеры и число ядрышек увеличиваются при повышении функциональной активности клетки.
Под электронным микроскопом в ядрышке обнаруживают три компонента - фибриллярный, гранулярный и аморфный.
4) Кариоплазма (ядерный сок) – жидкий компонент ядра, в котором располагаются хроматин и ядрышко. Содержит воду и ряд растворенных и взвешенных в ней веществ: РНК, гликопротеинов, ионов, ферментов, метаболитов.
Вопрос 12. Структура ядерной оболочки и ее молекулярная организация.
См вопрос 11
Вопрос 13. Хроматин интерфазного ядра. Эухроматин и гетерохроматин. Уровни укладки хроматина. Роль гистоновых белков в обеспечении структуры хроматина.
Хроматин – мелкие зернышки и глыбки материала, который обнаруживается в ядре клеток и окрашивается основными красителями. Хроматин состоит из комплекса ДНК и белка и соответствует хромосомам, которые в интерфазном ядре представлены длинными, тонкими перекрученными нитями и неразличимы как индивидуальные структуры. Различают два вида хроматина:
Эухроматин соответствует сегментам хромосом, которые деспирализованы и открыты для транскрипции. Эти сегменты не окрашиваются и не видны в световой микроскоп.
Гетерохроматин соответствует конденсированным, плотно скрученным сегментам хромосом (что делает их недоступными для транскрипции). Он интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид гранул. Его скопления располагаются под кариолеммой, прерываясь в области пор (что обусловлено его связью с ламиной) и вокруг ядрышка (перинуклеолярный гетерохроматин), более мелкие глыбки разбросаны по всему ядру.
Тельце Барра - скопление гетерохроматина, соответствующее одной Х- хромосоме у особей женского пола, которая в интерфазе плотно скручена и неактивна.
Очевидна необходимость компактной упаковки молекул ДНК, ведь в деконденсированном состоянии длина одной молекулы ДНК, образующей каждую хромосому, равна в среднем, около 5 см, а общая длина молекул ДНК всех хромосом в ядре (диаметром около 10 мкм) составляет более 2 м. В клеточном ядре это осуществляется благодаря их связи со специальными основными (гистоновыми) белками. Компактная упаковка ДНК в ядре обеспечивает:
Упорядоченное расположение очень длинных молекул ДНК в небольшом объеме ядра
Функциональный контроль активности генов
Уровни упаковки хроматина:
Нуклеосома, d=11нм. Нуклеосомная нить и 8 молекул гистонов образуют сердцевину, имеющую дисковидную форму, на которую накручивается фрагмент ДНК. Нуклеосомы разделены короткими участками свободной ДНК.
Хроматиновая фибрилла, d=30нм. Представляет собой скрученную нуклеосомную нить.
Петельная структура, d=300нм. Хроматиновые фибриллы образуют петли.
Хромонема, d=700нм. Происходит укладка нуклеомерных петель в области хромомерных учасков.
Хроматида, d=1400нм. Хромонемы укладываются спирально или петлеобразно, образуя хроматиду.
Роль гистоновых белков в обеспечении структуры хроматина. Гистоны в октамере имеют подвижный N-концевой фрагмент («хвост») из 20 аминокислот, который выступает из нуклеосом и важен для поддержания структуры хроматина и контроля над генной экспрессией.
Вопрос 15. Ядрышко. Структура ядрышка (СМ и ЭМ). Основные компоненты ядрышка. Роль ядрышка в синтезе рРНК и образовании рибосом.
Ядрышко – образовано специализированными участками (петлями) хромосом, которые называются ядрышковыми организаторами. У человека такие участки имеются в пяти хромосомах - 13-й, 14-й, 15-й, 21-й и 22-й, где располагаются многочисленные копии генов, кодирующих рРНК. Функции ядрышка заключаются в синтезе рРНК и ее сборке в предшественники рибосомальных субъединиц.
Ядрышко выявляется в интерфазном ядре на светооптическом уровне как мелкая плотная гранула диаметром 1-3 мкм, интенсивно окрашивающаяся основными красителями. Оно располагается в центре ядра или эксцентрично. Размеры и число ядрышек увеличиваются при повышении функциональной активности клетки.
Под электронным микроскопом в ядрышке обнаруживают три компонента:
Фибриллярный компонент – состоит из множества тонких (диаметром 5-8 нм) нитей и располагается преимущественно во внутренней части ядрышка. Он представлен преимущественно совокупностью первичных транскриптов рРНК.
Гранулярный компонент – образован скоплением плотных частиц диаметром 10-20 нм, которые соответствуют наиболее зрелым предшественникам субъединиц рибосом.
Аморфный компонент, в отличие от первых двух, окрашивается бледно. Он содержит участки расположения ядрышковых организаторов со специфическими РНК-связывающими белками и крупными петлями ДНК, активно участвующими в транскрипции рРНК.
Функции ядрышка заключаются в синтезе рРНК и ее сборке в предшественники рибосомальных субъединиц. При транскрипции генов ядрышковых организаторов начально формируется очень крупная молекула предшественника рРНК, которая связывается с белками, синтезированными в цитоплазме и импортированными в ядро с образованием РНП (рибонуклеопротеинов). Далее предшественник расщепляется на 3 вида РНК, которые выявляются в рибосомах. Два из них соединяются с добавочными белковыми молекулами, образуя предшественники большой субъединицы рибосомы, третий формирует предшественник малой субъединицы. Предшественники рибосомальных субъединиц далее по-отдельности транспортируются через ядерные поры в цитоплазму, где окончательно созревают.
Вопрос 16. Клеточный конвейер при синтезе белка. Морфологическая характеристика клетки, синтезирующей белки.
Клеточный конвейер – это сборка секреторного продукта на живой конвейерной ленте при участии различных клеточных органелл, она слагается из ряда этапов, происходящих в определённой последовательности.
Синтез сигнального пептида на полисомах.
Связывание сигнального пептида с СРЧ (сигнал распознающей частицей)
Прекращение синтеза белка (результат связывания с СРЧ)
Взаимодействие СРЧ со специфическим рецептором (причальный белок) на мембране ЭПС.
Удаление СРЧ от полисом
Разблокирование синтеза белка.
Присоединение большой субъединицы рибосом к мембране ЭПС
Отщепление сигнального пептида от полипептидной цепи
Поступление синтезируемого полипептида в полость ЭПС
Посттрансляционная модификация и поступление в КГ, секреция
Морфологическая характеристика клетки, синтезирующей белки:
Базофилия цитоплазмы
Крупные ядра
Большие – одно-два (иногда больше) ядрышка
Кариолемма нередко формирует инвагинации, увеличивающие ее поверхность
Множество ядерных пор, обеспечивающих интенсивный обмен ядра с цитоплазмой
Ядро с дисперсным хроматином
Много цистерн грЭПС и КГ, нередко обилие секреторных белковых включений
Умеренно либо сильно развиты митохондрии
Вопрос 17. Клеточный конвейер при синтезе углеводов и липидов. Морфологические особенности клеток, синтезирующих углеводы и липиды.
Клеточный конвейер – это сборка секреторного продукта на живой конвейерной ленте при участии различных клеточных органелл, она слагается из ряда этапов, происходящих в определённой последовательности.
Транскрипция ДНК с образованием мРНК
Процессинг мРНК с образованием иРНК
Выход из ядра иРНК
Синтез на свободных рибосомах ферментов для биосинтеза углеводов и липидов
Поступление ферментов в гиалоплазму или гладкую ЭПС, где происходит синтез углеводов или липидов
Поступление синтезированных веществ в КГ и формирование секреторной гранулы с выделением из клетки или сохранением в-в внутри клетки
Морфологическая характеристика клетки, синтезирующей липиды и углеводы:
При световой микроскопии цитоплазма оксифильна
Хорошо развита аЭПС
Ядро с частично конденсированных хроматином
Ядрышко отсутствует
Вопрос 18. Экстрагонадное происхождение половых клеток. Морфофункциональная характеристика мужской половой клетки и место ее образования.
Все половые клетки млекопитающих и человека берут начало от первичных половых клеток (ППК) – гоноцитов. Происхождение ППК до настоящего времени окончательно не выяснено. Не вызывает, однако, сомнения, что эти клетки возникают значительно раньше, чем появляются зачатки гонад, то есть они имеют экстрагонадное происхождение. Согласно существующим представлениям ППК могут быть обнаружены в первичной полоске уже на 16-18-й день развития, затем они перемещаются в желточную (внезародышевую) энтодерму у основания аллантоиса, мигрируют в энтодерму средней кишки, откуда и попадают в половые валики -- зачатки гонад. В последнее время получены данные о том, что ППК выделяются в самостоятельный эмбриональный зачаток значительно раньше, еще во время дробления и формирования бластоцисты. Попав в зачатки гонад, гоноциты впервые обнаруживают признаки полового диморфизма.
Сперматозоиды (спермии) человека образуются в течение всего активного полового периода. Продолжительность развития зрелых сперматозоидов из сперматогоний составляет около 72 дн. Подвижность обусловлена наличием жгутиков. Целенаправленному движению способствует хемотаксис (движение к химическому раздражителю/от него) и реотаксис (движение против тока жидкости).
Сперматозоиды имеют головку и хвост. Сперматозоид покрыт цитолеммой, которая в переднем отделе содержит рецептор, обеспечивающий узнавание рецепторов яйцеклетки.
Головка сперматозоида включает небольшое плотное ядро с гаплоидным набором хромосом. Передняя половина ядра покрыта плоским мешочком, составляющим чехлик сперматозоида. В нем располагается акросома. Она содержит набор ферментов, среди которых важное место принадлежит гиалуронидазе и протеазам, способным при оплодотворении растворять оболочки, покрывающие яйцеклетку. Чехлик и акросома являются производными комплекса Гольджи.
Хвостовой отдел сперматозоида состоит из шейки (связующей части), промежуточной, главной и терминальной частей. В связующей части, или шейке, располагаются центриоли – проксимальная, прилежащая к ядру, и дистальная, от которой начинается осевая нить (аксонема), продолжающаяся в промежуточной, главной и терминальной частях.
В промежуточной части располагается аксонема с митохондриями, расположенными по спирали. От микротрубочек отходят парные выступы, или «ручки», состоящие из другого белка – динеина, обладающего АТФазной активностью. Динеин расщепляет АТФ, вырабатываемую митохондриями, окружающими аксонему, и преобразует химическую энергию в механическую, за счет которой осуществляется движение спермия.
Главная |
 Скачать 483.12 Kb.
Скачать 483.12 Kb.