Вопросы на 3 тур. Вопросы к iii туру олимпиады по гистологии для студентов лечебного и педиатрического факультетов
 Скачать 204.5 Kb. Скачать 204.5 Kb.
|

Эмбриональный и постэмбриональный гистогенез.Развитие коры больших полушарий (неокортекса) человека в эмбриогенезе происходит из вентрикулярной герминативной зоны конечного мозга, где расположены малоспециализированные пролиферирующие клетки. Из этих клеток дифференцируются нейроциты неокортекса. При этом клетки утрачивают способность к делению и мигрируют в формирующуюся корковую пластинку. Вначале в корковую пластинку поступают нейроциты будущих I и VI слоев, т.е. наиболее поверхностного и глубокого слоев коры. Затем в нее встраиваются в направлении изнутри и кнаружи последовательно нейроны V, IV, III и II слоев. Этот процесс осуществляется за счет образования клеток в небольших участках вентрикулярной зоны в различные периоды эмбриогенеза (гетерохрон-но). В каждом из этих участков образуются группы нейронов, последовательно выстраивающихся вдоль одного или нескольких волокон радиальной глии в виде колонки. Цитоархитектоника. В двигательной зоне коры различают шесть основных слоев: молекулярный, наружный зернистый, пирамидный, внутренний зернистый, ганглионарный, слой полиморфных клеток. В первом (наружном) молекулярном слое почти нет тел нейронов. Обнаруживаются единичные горизонтально ориентированные нейроны, тангенциальные ветвления нервных волокон нижележащих нейронов и глиальные клетки. Второй, или наружный зернистый, слой содержит мелкие звездчатые и пирамидные нейроны размером около 10 мкм. Аксоны этих нейронов оканчиваются в III-м, IV-м и VI-м слоях коры, а дендриты поднимаются в молекулярный слой. Третий слой — это слой средних и крупных пирамидных нейронов. Аксоны этих клеток образуют ассоциативные нервные волокна, идущие через белое вещество и соединяющие соседние участки коры. Четвертый, или внутренний зернистый, слой содержит в основном мелкие звездчатые нейроны. Аксоны этих клеток, разветвляются в пределах соседних как выше, так и ниже лежащих слоев коры. Этот слой сильно развит в зрительной и слуховой зонах коры. Он состоит из сенсорных звездчатых нейронов, имеющих многочисленные ассоциативные связи с нейронами других типов. Пятый — ганглионарный — слой образован большими пирамидными нейронами (клетками Беца). Апикальные дендриты нейронов направляются в молекулярный слой. Аксоны этих клеток идут в белое вещество, образуя комиссуральные и проекционные нервные волокна, и прежде всего пирамидные пути. Шестой слой — слой полиморфных нейронов — содержит также много эфферентных пирамидных нейронов. Кроме того, здесь есть веретенообразные нейроны. Дендриты нейронов шестого слоя пронизывают всю толщину коры, достигая молекулярного слоя. 1) чувствительные нейроны — обеспечивают восприятие импульсов непосредственно с рецепторов и от ядер таламуса, а через него от различных рецепторных полей; 2) моторные — посылают импульсы от коры к нижележащим структурам ЦНС и рабочим органам, являющиеся представителями нервных центров безусловных рефлексов в коре больших полушарий; 3) контактные — осуществляют связь между нейронами коры больших полушарий. Чувствительные нейроны. Расположены в III и IV слоях коры и образуют воспринимающие зоны: сенсорные и окружающие их ассоциативные. Сенсорные нейроны. Образуют сенсорные зоны. Каждая зона называется соответственно рецепции, в которой она участвует. Размеры каждой сенсорной зоны зависят от физиологической значимости для организма животного рецепции. Чем выше значимость, тем она больше. Выделяют следующие сенсорные проекционные зоны: 1) двигательная — расположена между лобной и теменной долями. Раздражение этой зоны вызывает сокращение мышц. Рядом с ней находится вторичная двигательная зона. Эти зоны имеют и сенсорные входы. Поэтому их называют первичная и вторичная мотосенсорные зоны; 2) соматосенсорная первичная и вторичная — расположена в лобной и теменной долях, вдоль центральной борозды; воспринимает импульсы с рецепторов кожи и двигательного аппарата через таламус; 3) слуховая — расположена в височной доле; воспринимает импульсы от слуховых рецепторов; 4) зрительная — лежит в затылочной области; воспринимает импульсы от рецепторов сетчатки глаз; 5) обонятельная и 6) вкусовая — лежат на внутренней поверхности коры; связаны, соответственно, с обонятельными рецепторами носовых раковин и вкусовыми рецепторами языка и ротовой полости. Имеют двусторонние связи с лимбической системой. Ассоциативные зоны располагаются рядом с проекционными зонами. Их нейроны участвуют в анализе информации, В осуществлении связи между сенсорными и двигательными нейронами. Без ассоциативных нейронов невозможен четкий анализ и синтез программы. Моторные нейроны. Располагаются в V слое коры больших полушарий, образуют в ней корковые отделы нервных центров безусловных рефлексов. Моторные нейроны объединяются группами и образуют моторные зоны. Каждая моторная зона обеспечивает связь коры с органами организма. Моторные зоны способны переводить органы из состояния покоя в деятельное. Контактные нейроны. Осуществляют связь между разными нейронами в коре больших полушарий. В коре имеется большое количество глиальных клеток, выполняющих опорную, обменную, секреторную роли, а также участвующих в хранении следов осуществленных реакций. 4. Кора полушарий большого мозга. Миелоархитектоника коры полушарий большого мозга. Представление о модульной организации коры полушарий: межнейрональные связи, тормозная система, афферентное и эфферентное звенья КОРА БОЛЬШИХ ПОЛУШАРИЙ: МОЛЕКУЛЯРНЫЙ – мелкие ассоциативные клетки, их аксоны идут параллельно поверхности коры и образуют тангенциальное сплетение. НАРУЖНЫЙ ЗЕРНИСТЫЙ – мелкие нейроны различной формы, их дендриты уходят в тангенциальное сплетение, а аксоны частично в белое вещество или в тангенциальное сплетение. ПИРАМДНЫЙ – представлен мелкими и средними пирамидами, аксоны уходят в белое вещество ВНУТРЕННИЙ ЗЕРНИСТЫЙ – мелкие звездчатые нейроны, строение как в наружном ГАНГЛИОНАРНЫЙ – представлен гигантскими клетками Беца – крупные пирамиды, их дендриты поднимаются в молекулярный слой, а аксоны образуют главную часть кортикоспинального и кортиконуклеарного путей. СЛОЙ ПОЛИМОРФНЫХ КЛЕТОК – дендриты уходят в молекулярный слой, а аксоны в белое вещество в составе эфферентных путей головного мозга Выделяют два типа коры: агранулярный – характерен для моторных зон, хорошо развиты 3, 5, 6 слои коры. Гранулярный – характерен для чувствительных корковых центров, хорошо развиты 2 и 4 слои. МиелоархитектоникаСреди нервных волокон коры полушарий большого мозга можно выделить ассоциативные волокна, связывающие отдельные участки коры одного полушария, комиссуральные, соединяющие кору различных полушарий, и проекционные волокна, как афферентные, так и эфферентные, которые связывают кору с ядрами низших отделов центральной нервной системы. Проекционные волокна в коре полушарий образуют радиальные лучи, заканчивающиеся в III - пирамидном слое. Кроме уже описанного тангенциального сплетения I - молекулярного слоя, на уровне IV - внутреннего зернистого и V - ганглионарного слоев расположены два тангенциальных слоя миелиновых нервных волокон - соответственно, наружная полоска Байярже и внутренняя полоска Байярже. Последние две системы являются сплетениями, образованными конечными отделами афферентных волокон. Модульный принцип организации корыВ коре полушарий большого мозга описаны повторяющиеся блоки, или модули, нейронов, которые рассматривают как ее морфофункциональные единицы, способные к относительно автономной деятельности. Они имеют форму цилиндров, или колонок, проходящих вертикально через всю толщу коры. Каждый модуль включает афферентные пути, систему локальных связей и эфферентные пути. К афферентным путям относятся кортико-кортикальные и таламо-кортикальные волокна. Модуль организован вокруг кортико-кортикальных волокон, представляющих собой аксоны пирамидных клеток либо этого же полушария, либо противоположного. Кортико-кортикальные волокна образуют окончания во всех слоях коры данного модуля. В модуль входят также таламо-кортикальные волокна, оканчивающиеся в IV слое коры на шипиковых звездчатых нейронах и базальных дендритах пирамидных нейронов. Эфферентные пути формируются аксонами крупных и гигантских пирамидных нейронов, а также аксонами веретеновидных и некоторых других клеток VI слоя коры. Система локальных связей формируется вставочными нейронами модуля, которые включают более десятка типов клеток. Большая часть из них является тормозными и регулирует активность преимущественно пирамидных нейронов. Из тормозных нейронов модуля наибольшее значение имеют: аксо-аксональные клетки; клетки-"канделябры"; корзинчатые клетки; клетки с двойным букетом дендритов; клетки с аксональной кисточкой. Система тормозных нейронов играет роль фильтра, тормозящего часть пирамидных нейронов коры. 5. Мозжечок. Общая морфофункциональная характеристика. Гистогенез. Нейронный состав и глия коры мозжечка; афферентное и эфферентное нервные волокна коры мозжечка. Тормозные нейроны. Межнейрональные связи. МОЗЖЕЧОК – центральный орган равновесия и координации движений. Порыт корой (серое вещество). КОРА (нейральные взаимоотношения): МОЛЕКУЛЯРНЫЙ слой – образован двумя основными типами клеток: ЗВЕЗДЧАТЫЕ – мелкие и крупные, лежат в верхней 1/3 слоя, по функции – вставочные, тормозные нейроны. Могут заканчиваться на телах грушевидных клеток, образуя корзинку. КОРЗИНЧАТЫЕ – лежат в нижней 1/3 слоя. Их аксоны образуют корзинки вокруг тел грушевидных клеток. ГАНГЛИОНАРНЫЙ СЛОЙ – представлен одним слоем глеток – клетки Пуркинье (грушевидные) – клетки лежат в один ряд, дендриты располагаются в одной плоскости перпендикулярно поверхности мозжечка, имеют 2-3 главных дендрита, от их основания уходит аксон в белое вещество, при этом он образует обратные связи. ЗЕРНИСТЫЙ ЗЕРНА – многочисленные дендриты контактируют дендритами с афферентным (моховидным) волокном, образуя птичью лапку. Аксоны поднимаются в молекулярный слой, где Т-образно ветвятся и образуют синапсы с дендритами грушевидных клеток ГОЛЬДЖИ С КОРОТКИМИ НЕЙРИТАМИ – их аксоны заканчиваются проксимальнее клубочка мозжечка, лазающие волокна к клеткам Пуркинье. По функции – тормозные, вставочные. ГОЛЬДЖИ С ДЛИННЫМИ НЕЙРИТАМИ – их нейриты уходят в белое вещество и обеспечивают связь между различными слоями коры мозжечка. ВЕРЕТЕНОВИДНЫЕ (горизонтальные) – располагаются между зернистым и ганглионарным слоем, аксоны образуют коллатерали в зернистый слой и далее уходят в белое вещество. Развитие. Мозжечок образуется за счет разрастания дорсолатеральной стенки нервной трубки в области заднего мозга. В первые недели развития человека миграция нейробластов матричной зоны приводит к закладке ядер и клеток Пуркинье. На 9-11-й нед матричные стволовые клетки отделяются от эпендимного слоя и мигрируют (первичная миграция) на поверхность зачатка мозжечка. Там они образуют наружный герминативный слой (к 21-й нед развития его толщина составляет 6-9 клеточных слоев). Отсюда клетки, дифференцируясь в нейробласты, мигрируют в обратном направлении. Продвижение клеток через слой клеток Пуркинье направляет радиальная (бергмановская) глия. Скапливаясь в глубине коры ниже слоя клеток Пуркинье, нейробласты формируют внутренний зернистый слой коры. Такая миграция нейробластов способствует по мере их созревания формированию послойной структуры коры мозжечка и образованию характерных для нее внутренних межнейрональных связей. Процессы дифференциации в коре продолжаются и после рождения. Так, поверхностный герминативный слой сохраняется в мозжечке человека в течение первого года жизни. До 5 лет индивидуальная вариабельность строения коры мозжечка выражена слабо. Однако продолжающиеся гистогенетические процессы, которые заключаются в росте и усложнении ветвлений отростков нейронов коры, процессы их специализации к 20 годам жизни приводят к отчетливой индивидуальной вариабельности строения гистологических элементов коры. ВОЛОКНА: моховидные – идут в составе оливо-мозжечкового и мосто-мозжечкового путей, оказывают на клетки Пуркинье возбуждающее действие, которое большей частью гасится вставочными тормозными нейронами. Лазящие – поступают по спино-мозжечковому и вестибуло-мозжечковому путям, информация при этом не гасится. Они пересекают зернистый слой, прилегают к грушевидным нейронам и стелются по их дендритам, заканчиваясь на их поверхности синапсами.Лазящие волокна передают возбуждение непосредственно грушевидным нейронам Грушевидные нейроциты - основная функциональная звено коры мозжечка, на обеспечение нормальной деятельности которой направлена на активность всех других клеточных элементов коры, афферентных мохообразных и лианоподобных волокон. Только аксоны грушевидных нейронов выходят из коры мозжечка и реализуют регуляторное влияние мозжечка на организм. В глубине белого вещества локализованы подкорковые ядра мозжечка - зубчатое, коркообразные, шаровидные, ядро шатра, к которым поступают импульсы по аксонам грушевидных клеток и которые, таким образом, выполняют функцию переключателей. Афферентные волокна, поступающие в кору мозжечка, представлены двумя видами - моховидными и лазящими волокнами. Моховидные волокна через клетки-зерна оказывают на грушевидные клетки возбуждающее действие. Они заканчиваются в клубочках зернистого слоя мозжечка в виде расширений-розеток, где вступают в контакт с дендритами клеток-зерен. Каждое моховидное волокно дает ветви ко многим клубочкам мозжечка, и каждый клубочек получает ветви от многих моховидных волокон. Аксоны клеток-зерен по параллельным волокнам молекулярного слоя передают импульс дендритам грушевидных, корзинчатых, звездчатых нейронов зернистого слоя. Лазящие, или лиановидные, волокна пересекают зернистый слой, прилегают к грушевидным нейронам и стелются по их дендритам, заканчиваясь на их поверхности возбуждающими синапсами. Лазящие волокна передают возбуждение непосредственно грушевидным нейронам. С каждой клеткой Пуркинье обычно контактирует одно такое волокно. Таким образом, возбуждающие импульсы, поступающие в кору мозжечка, достигают грушевидных нейронов или непосредственно по лазящим волокнам или по параллельным волокнам клеток-зерен. Торможение в мозжечке - функция звездчатых нейронов молекулярного слоя, корзинчатых нейронов, а также клеток Гольджи зернистого слоя. Аксоны двух первых, следуя поперек извилин и тормозя активность грушевидных клеток, ограничивают их возбуждение узкими дискретными зонами коры. Поступление в кору мозжечка возбуждающих сигналов по моховидным волокнам, через клетки-зерна и параллельные волокна может быть прервано тормозными синапсами больших звездчатых нейронов, локализованными на концевых ветвлениях дендритов клеток-зерен проксимальнее возбуждающих синапсов. Эфферентные волокна коры мозжечка представлены аксонами клеток Пуркинье, которые в виде миелиновых волокон направляются в белое вещество и достигают глубоких ядер мозжечка и вестибулярного ядра, на нейронах которых они образуют тормозные синапсы. Кора мозжечка содержит различные глиальные элементы. В зернистом слое имеются волокнистые и протоплазматические астроциты. Ножки отростков волокнистых астроцитов образуют периваскулярные мембраны, являющиеся компонентом гемато-энцефалического барьера, а также оболочки вокруг клубочков мозжечка. Во всех слоях в мозжечке имеются олигодендроциты. Особенно богаты этими клетками зернистый слой и белое вещество мозжечка. В ганглионарном слое между грушевидными нейронами лежат особые астроциты с темными ядрами - клетки Бергмана. Отростки этих клеток направляются к поверхности коры и образуют глиальные волокна молекулярного слоя мозжечка (волокна Бергмана), поддерживающие ветвления дендритов грушевидных клеток. Микроглия в большом количестве содержится в молекулярном и ганглионарном слоях. Межнейрональные связи. Афферентные волокна, поступающие в кору мозжечка, представлены двумя видами — моховиднымии так называемыми лазящимиволокнами. 6. Вегетативная нервная система. Эмбриогенез. Общая морфофункциональная характеристика. Центральный и периферический отделы. Вегетативная рефлекторная дуга. Строение интрамурального вегетативного нервного узла. АВТОНОМНАЯ НЕРВНАЯ СИСТЕМА – регулирует моторику внутренних органов, кровяное давление, температуру, обменные процессы. Делится на симпатическую и парасимпатическую. Обе регулируют деятельность каждого органа (кроме мочевого пузыря и матки). . 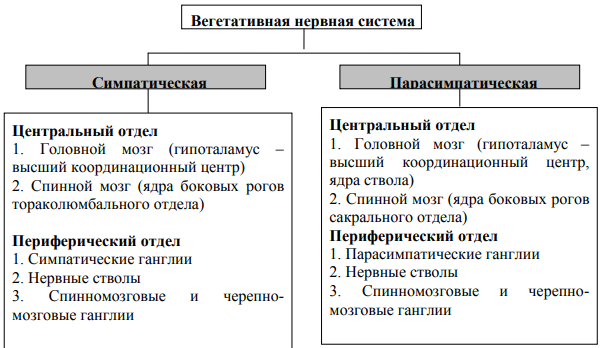 ЯДРА ВНС – находятся в среднем и продолговатом мозге, а так же в боковых рогах грудных, поясничных и крестцовых сегментов спинного мозга. К симпатической НС относятся вегнитативные ядра боковых рогов грудного и верхнепоясничного отдела спинного мозга. К парасимпатике: вегетативные ядра 3, 7, 9, 10 пары черепных нервов и вегитативные ядра крестцовых отделов спинного мозга. Мультиполярные нейроны – ассоциативные нейроны рефлекторных дуг ВНС. Идут к ганглиям. Периферические узлы автономной нервной системы лежат как вне органов (симпатические паравертебральные и превертебральные узлы, парасимпатические узлы головы), так и в стенке органов в составе интрамуральных нервных сплетений пищеварительного тракта, сердца, матки, мочевого пузыря и др. Паравертебральные узлы расположены по обе стороны позвоночника и со своими соединительными стволами образуют симпатические цепочки. Превертебральные узлы образуют кпереди от брюшной аорты и ее главных ветвей брюшное сплетение, в состав которого входят чревный, верхний брыжеечный и нижний брыжеечный ганглии. Автономные узлы снаружи покрыты соединительнотканной капсулой. Прослойки соединительной ткани проникают внутрь узла, образуя его остов. Узлы состоят из мультиполярных нервных клеток, весьма разнообразных по форме и величине. Дендриты нейронов многочисленны и сильно ветвятся. Аксоны в составе постганглионарных (обычно безмиелиновых) волокон поступают в соответствующие внутренние органы. Каждый нейрон и его отростки окружены глиальной оболочкой. Эмбриогенез На 3-й неделе эмбрионального развития человека в головном и спинном мозге в результате размножения и дифференцировки нейробластов возникают вегетативные центры. Источником нейронов периферического отдела вегетативной нервной системы является ганглиозная пластинка, которая возникает из нервных валиков (переходная область между нервной пластинкой и кожной эктодермой). Название “ганглиозной пластинки” обусловлено тем, что из нее образуются все нервные узлы. В основе образования вегетативных узлов лежит миграция клеток из гаглиозной пластинки на периферию (рис.5). Первыми закладываются узлы I порядка (околопозвоночные), несколько позже - узлы II (предпозвоночные) и III (органные) порядков. На первой стадии в результате периферической миграции клеток из ганглиозной пластинки образуются первичные сегментарные закладки вначале грудных и шейных, а затем, к концу I-го месяца, поясничных и крестцовых узлов симпатического ствола. На второй стадии происходит накопление клеток за счет прибывающих новых клеток, а также благодаря размножению уже прибывших. При этом происходит перемещение клеток вдоль симпатического ствола (продольная миграция) и первичные нервные узлы сливаются между собой, образуя сплошной несегментированный клеточный тяж (рис.6). На третьей стадии из клеточного тяжа формируются дефинитивные узлы симпатического ствола, которые, благодаря продольной миграции, содержат нейроны из многих сегментов ганглиозной пластинки. Предпозвоночные и органные вегетативные узлы формируются за счет продолжающейся в вентральном направлении миграции клеток и их размножения Нервные волокна различной функциональной принадлежности появляются неодновременно. Первыми обнаруживаются нервные волокна центрального происхождения - преганглионарные и афферентные, которые выходят из спинного и головного мозга в составе черепных и спинномозговых нервов. При этом преганглионарные волокна, вступающие в симпатический ствол, распространяются на значительные расстояния вверх и вниз и каждый узел получает их из многих сегментов. Такой характер распределения преганглионарных волокон, а также процесс продольной миграции клеток создают предпосылки к многосегментарной иннервации органов. Рефлекторная дуга строение В отличие от рефлекторных дуг соматической нервной системы моторный нейрон рефлекторных дуг вегетативного отдела идет не в составе серого вещества центральной нервной системы, а в одном из периферических ганглиев. Эфферентное звено вегетативных рефлекторных дуг двухчленно. Центральный нейрон всегда располагается в головном или спинном мозге. Второй нейрон (периферический) в симпатической нервной системе лежит в превертебральных или паравертебральных узлах, а в парасимпатической - в органе или вблизи него (рис. 180). В парасимпатической нервной системе оба нейрона холинергические. Пресинаптический полюс как первого, так и второго нейрона содержит мелкие, светлые синаптические пузырьки (40 - 60 нм) и одиночные крупные электроноплотные везикулы. В симпатической нервной системе первый нейрон холинергический, второй адренергический (медиатор - норадреналин). Морфологически адренергические синаптические пузырьки (30 - 50 нм в диаметре) характеризуются электроноплотной сердцевиной (гранулярные синаптические пузырьки). Кроме характерных для адренергических синапсов гранулярных везикул, в их составе, как и в холинергических, выявляют и большие гранулярные В симпатических нервных узлах, кроме свойственных для них мультиполярных нейронов, имеются группы мелких гранулосодержащпх интенсивно флуоресцирующих клеток (МИФ-клетки). Для них типичны короткие отростки и наличие в составе цитоплазмы гранулярных пузырьков. По флуоресценции и субмикроскопическому строению их пузырьки соответствуют пузырькам цитоплазмы клеток мозгового вещества надпочечников. Предполагается их участие в проведении нервного импульса от преганглионарных волокон на нейроны ганглия (роль интернейронов). МИФ-клетки рассматривают как внутриганглионарную тормозную систему. Возбуждаясь преганглионарными синапсами, они выделяют катехоламины, тормозящие передачу нервного импульса с преганглионарных волокон на симпатические нейроны узла |