коллоквиум. Вопросы коллоквиума по молекулярной биологии и медицинской генетики 1
 Скачать 0.8 Mb. Скачать 0.8 Mb.
|

|
Вопросы коллоквиума по молекулярной биологии и медицинской генетики № 1. 1. Предмет и задачи молекулярной биологии и медицинской генетики. Молекул. биология молодая наука. Она началась, когда Уотсон и Крик предложили структуру ДНК. Ее развитие началось, когда в биологии внедрились методы физики и химии, микроскопа, когда уже можно было видеть клетку и ее структуры. Молекулярная генетика позволила углубленно изучать процессы, происходящие в организме человека, послужила основой для изучения о наслед. заболеваниях и развитие микробиологии, вирусологии, эндокринология, иммунология, фармакологии. На ее основе создаются высокоэффективны лекар. Препараты. Также изучение генетической рекомбинации, где происходит обмен генами и участками хромосом. Это важно для изучениивиросологии и борьбы с вирусами. Создание генетической карты человека. Молекулярная био изучает строение и функции нуклеиновых кислот, принцип реализации передачи наслед инф. И фунции клеток деление рост итд. Задачи молекбио: Структурно-функц организации генетического аппарата Механизм реализации наследственной информации Молекулярных механизмов взаимодействия вырусов с клетками Закономерностей иммунных реакций организмов Появление злокачественности клеток в ходе развитии организмов специализации клеток. 2. Основные этапы развития и достижения молекулярной биологии и медицинской генетики. Дата рождения молекулярной биологии и генетики апрель 1953 года, когда в научном журнале “Nature” появилась статья с предложением прос-транственной модели ДНК, предложенная Френсис Крик и Джеймс Д. Уотсон (Нобелевская премия в области физиологии и медицины, 1962). 1957 Структура нуклеотидов и нуклеотидсодержащие коферменты; 1958 Полуконсервативная репликация ДНК; 1961-1966 Расшифровка генетического кода; 1972 Синтезирован ген тРНК (Корана и др.); 1977 Прерывистость генов у эукариот; 1980 Разработка методов экспериментальных манипуляций с ДНК; 1983 Открытие мобильных элементов; 1989 РНК как катализатор (рибозимы); 1993 Полимеразная цепная реакция (ПЦР) и олигонуклеотидный сайт- направленный мутагенез; 1994 Опубликованы генетические и физические карты хромосом человека; 1995Секвенирование геномов бактерий; 1997 Клонирование млекопитающих из дифференцированной соматической клетки; 2001 Секвенирование генома человека. 3. Клетка – элементарная единица живого. Клеточная теория. 1.Клетка — это элементарная, функциональная единица строения всего живого. 2. Клетка — единая система, она включает множество закономерно связанных между собой элементов, представляющих целостное образование, состоящее из сопряжённых функциональных единиц — органоидов. 3. Клетки всех организмов гомологичны. 4. Клетка происходит только путём деления материнской клетки. Клетка составляет основу строения, жизнедеятельности и развития всех живых форм – одноклеточных, многоклеточных и даже неклеточных. Благодаря заложенным в ней механизмам клетка обеспечивает обмен веществ, использование биологической информации, размножение, свойства наследственности и изменчивости, обуславливая тем самым присущие органическому миру качества единства и разнообразия Дополнительные положения клеточной теории: Для приведения клеточной теории в более полное соответствие с данными современной клеточной биологии список её положения часто дополняют и расширяют. Во многих источниках эти дополнительные положения различаются, их набор достаточно произволен. 1. Клетки прокариот и эукариот являются системами разного уровня сложности и не полностью гомологичные друг другу. 2. В основе деления клетки и размножения организмов лежит копирование наследственной информации - молекул нуклеиновых кислот («каждая молекула из молекулы»). Положения о генетической непрерывности относится не только к клетке в целом, но и к некоторым из её более мелких компонентов - к митохондриям, хлоропластам, генам и хромосомам. 3. Клетки многоклеточные обладают генетическими потенциями всех клеток данного организма, равнозначны по генетической информации, но отличаются друг от друга разной экспрессией (работой) различных генов, что приводит к их морфологическому и функциональному разнообразию - к дифференцировке. 4. Особенности клеточной организации прокариот. Прокариотический тип клеточной организации • Современные прокариоты представлены бактериями и сине-зелеными водорослями. У прокариот можно обнаружить все основные метаболические пути и три главных способа получения энергии – гликолиз, дыхание и фотосинтез. Прокариоты имеют оболочку, состоящую из наружного слоя и плазматической мембраны, цитоплазму. В цитоплазме содержатся рибосомы и полисомы, которые образуют белоксинтезирующий аппарат клетки. Отличительной особенностью прокариот является своеобразное строение генетического аппарата. 1.Бактерии и все прокариоты не имеют морфологически обособленного ядра. Эквивалентом ядра является нуклеотид. 2.Генетический аппарат представлен главной кольцевой хромосомой. Хромосома состоит из одной молекулы ДНК и не содержит типичных хромосомных белков –гистонов 3.Внехромосомная ДНК предоставлена плазмидами. Плазмиды-это кольцевые фрагменты ДНК,каждая из которых состоит из нескольких тысяч нуклеотидных пар. Размножается прямым делением. 5. Особенности клеточной организации эукариот. Эукариотический тип клеточной организации. Эукариотические клетки больше по размеру и имеют более сложную организацию. 1. Эукариотические клетки имеют морфологическое обособленное ядро, вещество которого отделено от цитоплазмы ядерной оболочкой. 2. Основное вещество ядра представлено хроматином. 3. Ядро расположено в центре клетки, с хорошо выраженной ядерной оболочкой. 4. Жизненный цикл эукариотических клеток сопровождается сложной реорганизацией наследственного аппарата. Эукариотичекие клетки включают следующие основные компоненты: плазмалемму, цитоплазму, ядро. Эукариотические клетки имеют цитоплазму, сложную в химическом и структурном отношении. К основным компонентам цитоплазмы относятся: гиалоплазма, цитоскелет, органоиды и включения. Включения – непостоянные структурные компоненты цитоплазмы. Классификация включений: Трофические включения – лецитин в яйцеклетках, гликоген, липиды, имеются почти во всех клетках. Секреторные включения – секреторные гранулы в секретирующих клетках (зимогенные гранулы в ацинозных клетках поджелудочной железы, секреторные гранулы в эндокринных железах и другие). Экскреторные включения – вещества, подлежащие удалению из организма (например, гранулы мочевой кислоты в эпителии почечных канальцев). Пигментные включения – меланин, гемоглобин, липофусцин, билирубин и другие. Эти включения имеют определенный цвет и придают окраску всей клетке (меланин – черный или коричневый, гемоглобин – желто-красный и так далее). Пигментные включения характерны только для определенных типов клеток (меланин содержится в меланоцитах, гемоглобин – в эритроцитах). Однако, липофусцин может накапливаться во многих типах клеток обычно при их старении. Его наличие в клетках свидетельствует о их старении и функциональной неполноценности. •Органеллы - это постоянные структуры цитоплазмы, выполняющие в клетке жизненно важные функции. •Выделяют органеллы общего значения, к ним относят элементы канальцевой и вакуолярной системы в виде шероховатой и гладкой цитоплазматической сети, пластинчатый комплекс, митохондрии, рибосомы и полисомы, лизосомы, пероксисомы, микрофибриллы и микротрубочки, центриоли клеточного центра. •К органеллам специального значения относят, например, микроворсинки всасывающей поверхности эпителиальной клетки кишечника, реснички эпителия трахеи и бронхов, синаптические пузырьки, транспортирующие вещества – переносчики нервного возбуждения с одной нервной клетки на другую или клетку рабочего органа, миофибриллы, от которых зависит сокращение мыщцы. •По строению оргоноиды общего значения подразделяются на две группы: мембранные и немембранные. •К главным мембранным органоидам относятся: эндопазматическая сеть, пластинчатый комплекс Гольджи, лизосомы, митохондрии, секреторные пузырьки, микротельца. •К немембранным органоидам – свободные рибосомы и полисомы, микротрубочки, центриоли, микрофиламенты, микрофибриллы. 6. Вирусы- неклеточные формы жизни. Открыты рус ученым Ивановским в 1892г. Паразитирующие на генетическим уровне. Они проникают внутрь клетки и размножаются там. Вирусы имеют собств. ген аппарат, кодируют синтез новых вирусных частиц, используя ген аппарат хозяина. Есть днк и рнк содержащие вирусы. Белковая оболочка – капсид. Геномика большинства вирусов уже известна. развивается технология получения днк-вакцин от спида, гепатита с и др. внеклоточные вирусы называются вирионы, не способные размножаться. Виды вирусов: спиральная (табачная мозаика), иксоэдрическая(полиовирус, риновирус), сложная (комбинация иксоэд и спиральной формы, голова и хвост, бактериофаги. Они с помощью хвоста прикрепляются к бактерии, делают отверстия и впрыскивают свое днк.). Рнк полимераза чаще делает ошибки при копировании, чем ДНК полимеразы. 7. Строение, функции и классификация нуклеиновых кислот. Строение ДНК. Нуклеиновые кислоты – материальный субстрат наследственности и изменчивости, это макромолекулы, биополимеры мономерами которых являются нуклеотиды. Непрерывность существования живой клетки обусловлена двумя фундаментальными свойствами жизни: наследственностью и изменчивостью, которые обеспечиваются нуклеиновыми кислотами. Впервые нуклеиновые кислоты были открыты в 1868 г. (Ф. Мишер). Нуклеиновые кислоты – это высокомолекулярные соединения с определенной последовательностью мономеров - нуклеотидов. Информация о структуре белка, хранится и воспроизводится при помощи нуклеиновых кислот. Типы нуклеиновых кислот: 1. Дезоксирибонуклеиновая кислота (ДНК) 2. Рибонуклеиновая кислота (РНК) Дезоксирибонуклеиновая кислота(ДНК) Важнейшим процессом, происходящим во всех клетках, является синтез белка. Информация о последовательности аминокислот, составляющих первичную структуру белка, заключена в ДНК. Молекула дезоксирибонуклеиновой кислоты (ДНК) состоит из двух спирально закрученных цепей. Общепринятая модель структуры ДНК была впервые предложена Дж. Уотсон и Ф. Криком в 1953г. Модель строение ДНК, предложенная Дж. Уотсоном и Ф. Криком, основывались на следующих данных: 1. ДНК представляет собой полимер, состоящий из нуклеотидов, соединенных 3/ - 5/ фосфодиэфирными связями. 2. Состав нуклеотидов в ДНК подчиняется правилам Чаргаффа(правило комплементарности). В молекуле ДНК количество аденина равно количеству тимина, а количество гуанина — количеству цитозина: А = Т, Г = Ц. 3. Рентгенограммы волокон ДНК, впервые полученные М.Уилкинсом и Р.Франклин указывают на то, что молекулы обладают спиральной структурой. 4. Структура нативной ДНК стабилизируется водородными связями. 8. Особенности пространственной организации ДНК. Свойства и функции ДНК. Первичная структура. Мономером ДНК является нуклеотид ДНК. Нуклеотиды соединяются между собой фосфодиэфирной связью, образуя полинуклеотидную цепь. На полинуклеотидной цепи ДНК различают два конца 5’ и 3’, начало и конец. Образование фосфодиэфирной связи обеспечивает фермент ДНК-полимераза. Благодаря ДНК-полимеразе происходит наращивание полинуклеотидной цепи только на одном конце (3’). Начало любой полинуклеотидной цепи всегда несет фосфатную группу в положении 5’. • Полинуклеотидная цепь - это первичная структура ДНК. Вторичная структура ДНК • Вторичная структура - это две антипараллельные комплементраные полинуклеотидные цепи, соединенные между собой водородными связями, и образующие спираль. Диаметр спирали составляет 2 нм., длина шага- 3.4 нм., в каждый виток входит 10 пар нуклеотидов. Комплементарность в молекуле ДНК: Аденин – Тимин, Гуанин – Цитозин • Аденин одной цепи соединяется двумя водородными связями с тимином другой цепи. Между гуанином и цитозином – три водородные связи. Такое соединение азотистых оснований обеспечивает прочную связь двух цепей. • Антипараллельность в молекулеДНК: 5 конец одной цепи соединяется с 3-концом другой Третичная структура – упакованная молекула ДНК. Двойная спираль ДНК на некоторых участках может подвергаться дальнейшей спирализации. Суперспиральная структура обеспечивает экономную упаковку огромной молекулы ДНК. Упаковка происходит при помощи гистоновых и негистоновых белков. Одна молекула ДНК в комплексе с белками в ядрах образует нить хроматина. При делении хроматин конденсируется в хромосому 9. Особенности строения митохондриальной ДНК. Митохондриальная ДНК — ДНК, находящаяся (в отличие от ядерной ДНК) в митохондриях, органоидах эукариотических клеток. Гены, закодированные в митохондриальной ДНК, относятся к группе плазмагенов, расположенных вне ядра (вне хромосомы). У большинства многоклеточных организмов митохондриальная ДНК наследуется по материнской линии. Яйцеклетка содержит на несколько порядков больше копий митохондриальной ДНК, чем сперматозоид. В сперматозоиде обычно не больше десятка митохондрий (у человека — одна спирально закрученная митохондрия). Обычно происходит деградация митохондрий сперматозоида после оплодотворения, т.к большая часть митохондрий сперматозоида находятся в основании жгутика, которое при оплодотворении иногда теряется. 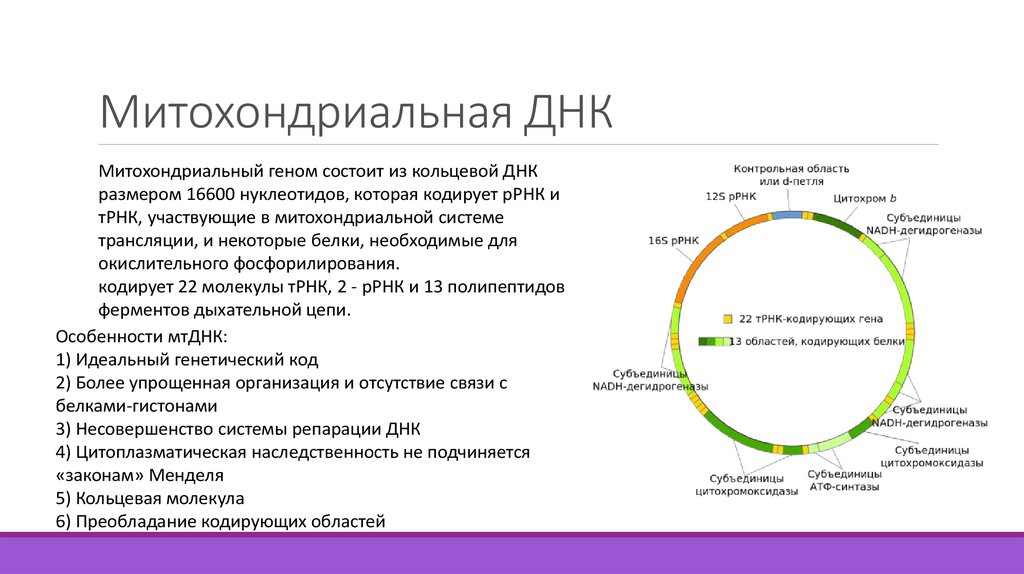 10.Репликация ДНК. Образование репликативного комплекса. Фазы репликации. Репликация – способность к самокопированию, это одно из основных свойств наследственного материала. Репликация ДНК обеспечивает воспроизведение наследственной информации при образовании новых клеток. Клеточное деление бывает двух типов: митоз (количество хромосом не изменяется) и мейоз (количество хромосом уменьшается в два раза). • Репликация ДНК происходит полуконсервативным способом, когда каждая дочерняя молекула ДНК содержит одну материнскую и одну синтезированную цепь. • Процесс репликации осуществляется сложным ферментным комплексом (15-20 различных белков). На каждой хромосоме работают несколько репликативных комплексов. На хромосоме имеется много точек начала репликации ДНК – в среднем около 40. Удвоение происходит не последовательно, а одновременно во многих местах сразу. В каждой точке репликации начинают работать два ферментативных комплекса, двигаясь в противоположные стороны – образуются две репликативные вилки. Между вилками образуется постепенно расширяющееся «вздутие» или «глазок». Каждый комплекс реплицирует обе цепи. В конечном счете соседние зоны репликации (глазки) сливаются и вся молекула ДНК оказывается удвоенной. 11. Этапы репликации ДНК. Особенности синтеза лидирующей и отстающей цепи Ферментативный комплекс функционирует так, что одна из двух синтезируемых им цепей растет с некоторым опережением по сравнению с другой цепью. Первая цепь называется лидирующей вторая запаздывающей. Запаздывающая же цепь образуется в виде серии относительно коротких фрагментов – примерно по 1500 нуклеотидов (фрагменты Оказаки). Этапы репликации • 1. Инициация репликации осуществляется в особых участках ДНК, обозначаемых ori (от англ. оrigin - начало). Точки начала репликации на молекуле ДНК имеют специфическую последовательность основании, богатую парами А-Т. Процесс начинается с того, что с каждой такой последовательностью связывается несколько молекул узнающих белков. Двойная спираль ДНК в этих локусах, под действием фермента геликазы, разделяется на две цепи, при этом, как правило, по обе стороны от точки начала репликации образуются области расхождения полинуклеотидных цепей – репликационные вилки, которые движутся в противоположных от локуса ori направлениях. • Фермент геликаза разделяет нити двойной цепи ДНК. Образующиеся при этом одинарные цепи ДНК связываются специальными дестабилизирующими белками (SSB-белки), которые растягивают остовы цепей, делая их азотистые основания доступными для связывания с комплементарными нуклеотидами. Разделение спирально закрученных цепей родительской ДНК ферментом геликазы вызывает появление супервитков перед репликационной вилкой. Фермент топоизомераза I разрывает одну цепь ДНК и дает возможность вращаться другой цепи, тем самым ослабляет напряжение в двойной спирали ДНК. 2. Полимеризация цепи ДНК • Синтез цепи ДНК (полимеризацию нуклеотидной цепи) осуществляет ДНК-полимераза. ДНК-полимераза присоединяет очередной нуклеотид в 3’ положении предшествующего нуклеотида, цепь постоянно удлиняется на ее 3’ конце. Особенностью ДНК-полимеразы является ее не способность начать синтез новой поленуклеотидной цепи путем простого связывания двух нуклеозиттрифосфатов необходим 3’-ОН-конец какой-либо полинуклеотидной цепи, спаренной с матричной цепью ДНК, к которой ДНК-полимераза может лишь добавлять новые нуклеотиды. Такую полинуклеотидную цепь называют затравкой или праймером. Роль затравки для синтеза полинуклеотидных цепей ДНК в ходе репликации выполняют короткие последовательности РНК, образуемые при участии фермента РНК-праймазы. • Специальный белок выполняет функции активатора праймазы. После чего праймаза, используя в качестве матрицы соответствующий участок однуцепочечную ДНК, синтезирует короткую РНК-затравку, или праймер. Указанная особенность ДНК-полимеразы означает, что матрицей при репликации может служить лишь цепь ДНК, несущая спаренную с ней затравку, которая имеет свободный 3’-ОН-конец. Этот фермент образует межнуклеотидную (фосфодиэфирную) связь. Для начала синтеза новой цепи для ДНК-полимеразы необходимо наличие свободного 3’-OH конца, к которому будет присоединен 5’-конец следующего нуклеотида, т.е. необходимо наличие праймера (затравки) комплементарного одной из материнских цепей. Для этого в точке начала репликации фермент РНК-праймаза синтезирует короткую цепь РНК (РНК-затравка, или праймер). • У эукариот известно 5 разных ДНК-полимераз. Из них β и ε полимеразы участвую в репарации ДНК, γ-полимераза - в репликации митохондриальной ДНК, а α- и δ-полимеразы в репликации ядерной РНК. Аналогом бактериальной ДНКполимеразы III у эукариот является комплекс α- и δ-ДНКполимераз (синтез цепи ДНК). Функцию ДНК-полимеразы I выполняют: специальная нуклеаза (удаление РНК-затравки) и β-ДНКполимераза (достраивание брешей). Особенности синтеза лидирующей и отстающей цепей. • Поскольку синтез цепи ДНК возможен только в направлении 5’- 3’, дочерние цепи ДНК в репликативной вилке синтезируются не с одинаковой скоростью. Ферментный комплекс периодически прекращает образование лидирующей цепи и переходит на вторую родительскую цепь и синтезирует очередной фрагмент Оказаки запаздывающей цепи. • Фрагменты Оказаки – короткие фрагменты отстающей цепи ДНК (с РНК-праймером на 5’-конце – 100-200 нуклотидов). 3. Терминация - конечный этап – завершение репликации. • Сшивание соседних фрагментов ДНК осуществляет ДНКлигаза. Этот фермент образует межнуклеотидную фосфодиэфирную связь. Молекула ДНК окажется реплицированной не полностью, если не произойдет специальный процесс репликации ее концов, или теломерных участков. В этом процессе ключевую роль играет фермент теломераза. • Репликация заканчивается в участке гена (молекулы ДНК), содержащим несколько специфических последовательностей нуклеотидов, называемых терминационными сайтами. Это короткие (около 23 пар нуклеотидов) последовательности. Терминация - конечный этап – завершение репликации. • У эукариот проблема недорепликации – концов линейных молекул ДНК мешается с помошью специального фермента – теломеразы, которая перед концом репликации наращивает недорепликированные 5‘ – концы ДНК дочерних рибосом короткими, повторяющимися последовательностями нуклеотидов или теломерами. • Теломеры располагаются на концах хромосом. В состав теломер входят специфические белки, не образующие нуклеосомные глобулы в коротких теломерах. • Плотность упаковки теломер позволяет отнести их к гетерохромативновой структуре хромосом. • 12.Особенности репликации теломерной ДНК. Роль теломер и теломераз в репликации концевых отделов ДНК. Теломеры – концевые участки хромосом, которые не несут генетической информации и защищают ДНК от расщепления нуклеазами, и предотвращают от слияния. В результате репликации молекула ДНК окажется реплицированной не полностью, если не произойдет репликации теломер. Существует фермент, восстанавливающий длину теломер – теломераза (ДНКнуклеотидилтрансфераза). Теломераза есть только в зародышевых клетках и клетках опухолей, в соматических клетках теломераза отсутствует. Функции теломер: • Участвует в фиксации хромосом к ядерному матриксу, обеспечивая правильную ориентацию хромосом в ядре. • Соединяют друг с другом концы сестринских хроматид, образующихся в хромосоме после S – фазы. Структура теломер однако допускает расхождение хроматид в анафазе. Мутация гена теломеразной РНК с изменением нуклеотидной последовательности теломер приводит к не расхождению хроматид. • Предохраняют от недорепликации генетические значимые отделы ДНК в отсутствие теломераз. • Стабилизируют в присутствии теломераз концы разорванных хромосом путем добавления к ним теломер с возможностью их функционирования. Примером является восстановление функции гена α глобина у больных α – талассемией путем добавления теломер к точкам разрыва длинного плеча 16 хромосомы. • Влияют на активность генов. Гены, расположенные рядом с теломерами, функционально менее активны (репрессированны). Данный эффект носит название транскрипционного молчания или сайленсинга. Укорочение теломер приводит к отмене эффекта положения генов с активацией прителомерных генов. В основе сайленсинга может лежать действие белков (Rap 1, TRF 1), взаимодействующих с теломерами. • Выступают в качестве регулятора количества клеточных делений. Каждое деление клетки сопровождается укорочением теломеры на 50-65 пар нуклеотидов. В отсутсвиетеломеразной активности количество делений клетки будет определяться протяженностью оставшихся теломер 13. Молекулярная структура и функции основных компонентов клетки: оболочка, ядро, цитоплазма. Плазмолемма Плазмолемма – окружает клетку. Основа плазмолеммы - биологическая мембрана, которая покрыта снаружи слоем гликокаликса толщиной 10 – 20 нм. Строение плазмолеммы: 1.Липидной бислой (гидрофильная «головка» и гидрофобные «хвосты»); 2.Белки: интегральные, периферические; 3.Гликокаликс- основными составляющими гликокаликса служат комплексы полисахаридов с белками (гликопротеины) и жирами (гликолипиды). Функции плазмолеммы: •ограничивающая (барьерная), регуляция и обеспечение избирательной проницаемости веществ, раздел между гидрофильной и гидрофобной фазами, наличие ферментных комплексов наличие рецепторов. Свойства плазмолеммы: замкнутость, латеральная подвижность, асимметрия. Ядро Компоненты ядра: ядерная оболочка; хроматин; ядрышко; ядерной сок. Ядерная оболочка: обособляет генетический материал состоит из двух мембран и перинуклеарного пространства, пронизана порами диаметром 80-90 нм. С внутренней стороны ядерной мембраны Генетическая прикреплена ядерная информация заключена в ламина. нитях хроматина, которые находятся в ядерном матриксе. Ядрышко – плотное круглое тельце, размеры которого могут изменятся от 1 до 10 мкм и больше. Количество ядрышек может меняться в разные периоды жизнедеятельности клетки и организма. Ядерный сок – полужидкое вещество под ядерной оболочкой и представляет собой внутреннюю среду ядра. В состав ядерного входят различные белки, ферменты, рибосомальные белки, хромосомы, нуклеотиды, аминокислоты и другие вещества. Цитоплазма В цитоплазме различают: основное вещество (матрикс, гиалоплазма), цитоскелет, включения и органеллы. Основное вещество цитоплазмы заполняет пространство между плазмалеммой, ядерной оболочкой и другими внутриклеточными структурами. •Белковый состав гиалоплазмы разнообразен. Важнейшие из белков представлены ферментами гликолиза, обмена сахаров, азотистых оснований, аминокислот и липидов. •Ряд белков гиалоплазмы служат субъединицами из которых происходит сборка таких структур, как микротрубочки. Основное вещество цитоплазмы образует истинную внутреннею среду клетки, которая объединяет все внутриклеточные структуры и обеспечивает взаимодействие их с друг другом. Выполнение матриксом объединяющей, каркасной функции может быть связано с наличием микротрабикулярной сети образованной фибриллами 2–3 нм и пронизывающей всю цитоплазму. Включения - непостоянные компоненты цитоплазмы, которые служат запасными питательными веществами (жир, гликоген) продуктами, подлежащим выведению из клетки (гранулы секрета) балластными веществами (некоторыми пигментами). 14. Строение и функции гликокаликса. .Гликокаликс- основными составляющими гликокаликса служат комплексы полисахаридов с белками (гликопротеины) и жирами (гликолипиды). Гликокаликс (Рис. 5) выполняет рецепторную и маркерную функции, а также участвует в обеспечении избирательности транспорта веществ и пристеночном (примембранном) пищеварении. Наличие гликокаликса характерно для клеток животных (к отличие от прокариотов, растений и грибов, где его нет). 15.Цитоскелет клетки, строение и функции . Цитоскелет представляет собой сложную трехмерную сеть белковых нитей, которая обеспечивает способность эукариотических клеток сохранять определенную форму., а также осуществлять направленные и координированные движения самих клеток, так и отдельных органелл. • Цитоскелет состоит из микрофиламентов – длинные, тонкие образования, образующие пучки и обнаруживаются по всей цитоплазме. Актиновые микрофиламенты- диаметр 7 нм, обеспечивают мышечное сокращение и клеточное движение. Им приписывают каркасную роль и участие в организации внутриклеточных перемещений органелл и участков гиалоплазмы. Промежуточные филаменты- диаметр 10 нм, располагаются по периферии и в околоядерной зоне, выполняют механическую и каркасную функции. В эпителиальных, нервных, глиальных, мыщечных клетках, фибробластах они построены из разных белков. 16. Структура и функции внутриклеточных органелл общего значения: эндоплазматическая сеть, комплекс Гольджи, лизосомы. Эндоплазматическая сеть Эндоплазматическая сеть -расположена вокруг ядра, образованна мембранами, разветвленная сеть полостей и каналов: гладкая ЭПС участвует в углеродном и жировом обмене; шероховатая ЭПС обеспечивает синтез белков с помощью рибосом. Особенность строения шероховатой сети состоит в прикреплении к ее мембранам полисом. В силу этого она выполняет функцию синтеза определенной категории преимущественно удаляемых из белков, клетки, например секретируемых клетками желез. Пластинчатый комплекс Гольджи Комплекс сложной структуры, состоящий из мембран, гранул и вакуолей. Здесь осуществляется биохимическая модификация белков и липидов, сборка протеингликанов, накапливание и выведение продуктов. В комплексе Гольджи осуществляется биохимическаямодификациявеществ: гликозилирование белков и липидов; гликозилирование и сборка протеогликанов; добавление маннозо-6-фосфата; сортировка веществ для дальнейшего транспорта. В комплексе Гольджи происходит упаковка, хранение и выведение веществ синтезированных в ЭПС 17. Структура и функции внутриклеточных органелл общего значения: митохондрии, рибосомы, клеточный центр, микротрубочки, микрофиламенты. Митохондрии - двухмембранный органоид, внутренняя мембрана имеет выросты - кристы, внутри находится собственный генетический аппарат биосинтеза белков. Главная функция митохондрии - ферментативное извлечение из определенных химических веществ энергии (синтез АТФ), путем окисления. Энергия идет на разнообразную работу: механическую, химическую, осмотическую. Число митохондрий в животной клетке колеблется 150-1500. Митохондрии является энергетической базой клетки (или силовыми станциями клетки), в них вырабатывается аденозинтрифосфат (АТФ), являющийся источником энергии. Митохондрии способны к перемещению в клетке. Также способны к самоудвоению, т.е. размножаются делением. В клетках печени они живут 10-20 дней Рибосомы - сферические частицы (диаметр 20-30 нм), располагаются в цитоплазме свободно или прикреплены к мембранам ЭПС; осуществляют синтез белка. •Рибосома состоит из малой и большой субъедениц, объединение которых происходит в присутствии матричной (информационной) РНК (мРНК). Одна молекула мРНК обычно объединяет несколько рибосом наподобие нитки бус. Такую структуру называют полисомой. На полисомах гиалоплазмы образуются белки для собственных нужд (для «домашнего» пользования), а на полисомах гранулярной сети сентезируются белки, выводимые из клетки и используемые на нужды организма (например, пищевые ферменты, белки грудного молока). Встречается во всех клетках, способных к делению. Клеточный центр состоит из двух центриолей: дочернего и материнского, расположенных перпендикулярно друг к другу и создающими диплосому. Только одна из центриолей, а именно материнская, имеет множество дополнительных образований. Одни из них это сатиллиты, их численность непостоянна, и они располагаются по всей длине центриоля. Материнский участок диплосомы является источником создания микротрубочек. Центриоли имеют форму цилиндра длиной 0,3мкм и диаметром 0,1мкм. Стенки центриолей состоят из девяти групп протеиновых микротрубочек. Окружены центриоли областью, более светлой цитоплазмы, (Эту светлую область и называют клеточным центром) от которой отходят микротрубочки, и образовывают центросферу, состоящую из углеводов, белков, и липидов. микрофиламентов – длинные, тонкие образования, образующие пучки и обнаруживаются по всей цитоплазме. Актиновые микрофиламенты- диаметр 7 нм, обеспечивают мышечное сокращение и клеточное движение. Им приписывают каркасную роль и участие в организации внутриклеточных перемещений органелл и участков гиалоплазмы. Промежуточные филаменты- диаметр 10 нм, располагаются по периферии и в околоядерной зоне, выполняют механическую и каркасную функции. В эпителиальных, нервных, глиальных, мыщечных клетках, фибробластах они построены из разных белков. Микротрубочки - диаметр 25 нм, встречаются в цитоплазме клеток, структурных элементах (жгутиках и ресничках), митотическом веретене и центриолях. 18. Структура и функции внутриклеточных органелл специального значения. Включения клетки. •К органеллам специального значения относят, например, микроворсинки всасывающей поверхности эпителиальной клетки кишечника, реснички эпителия трахеи и бронхов, синаптические пузырьки, транспортирующие вещества – переносчики нервного возбуждения с одной нервной клетки на другую или клетку рабочего органа, миофибриллы, от которых зависит сокращение мыщцы. Включения •Включения – непостоянные структурные компоненты цитоплазмы. Классификация включений: Трофические включения – лецитин в яйцеклетках, гликоген, липиды, имеются почти во всех клетках. Секреторные включения – секреторные гранулы в секретирующих клетках (зимогенные гранулы в ацинозных клетках поджелудочной железы, секреторные гранулы в эндокринных железах и другие). Экскреторные включения – вещества, подлежащие удалению из организма (например, гранулы мочевой кислоты в эпителии почечных канальцев). Пигментные включения – меланин, гемоглобин, липофусцин, билирубин и другие. Эти включения имеют определенный цвет и придают окраску всей клетке (меланин – черный или коричневый, гемоглобин – желто-красный и так далее). Пигментные включения характерны только для определенных типов клеток (меланин содержится в меланоцитах, гемоглобин – в эритроцитах). Однако, липофусцин может накапливаться во многих типах клеток обычно при их старении. Его наличие в клетках свидетельствует о их старении и функциональной неполноценности. 19. Молекулярная структура, свойства, функции биологических мембран. Значение биологических мембран. Основные виды мембранных белков.   20. Основные классы мембранных липидов и их значение Фосфолипиды – поддерживают в растворимом состоянии неполярные липиды, транспорт жиров, жирных кислот и холестерина. Сфинголипиды – передача клеточного сигнала и в клеточном распознавании. Гликолипиды - они обладают рецепторной функцией, участвуют в межклеточных взаимодействиях и контактах Стероиды, холестерин - обеспечивает жёсткость и стабильность мембран 21.Транспорт через мембраны: активный, пассивный. Активный транспорт - это транспорт против градиента концентраций и происходит при затрате энергии. Он локализован в плазматической мембране практически всех клеток и переносит ионы калия и натрия против градиента концентраций с использованием энергии АТФ. Собственно насос это фермент Na+-K+-АТФ- аза, интегральный белок пронизывающий липидный бислой мембраны насквозь. Внутри клетки к активному центру этого фермента присоединяется 3 иона Nа+, при этом фермент активируется и расщепляет АТФ на АДФ и остаток фосфорной кислоты. Этот остаток присоединяется к самому ферменту и изменяет его пространственную конформацию. При этом с внутренней стороны мембраны закрывается ионный канал, но открывается на наружной поверхности. Ионы Na+ отсоединяются от фермента, но в это же время к другому активному центру фермента присоединяются 2 иона К+. Это вновь изменяет его пространственную конформацию, отщепляется остаток фосфорной кислоты и открывается канал для проникновения К+ внутрь клетки. Далее цикл повторяется. Т.о. при выносе из клетки 3 ионов Na+ в нее проникает 2 иона К+. Это ведет к появлению электрического потенциала, который называется трансмембранным электрохимическим потенциалом. Если насос прекратит работу, то ионы Na+ и К+ начнут перемещаться в обратном направлении. По такому же механизму транспротируются ионы Са2+ (фермент Са2+- АТФ-аз Пассивный транспорт Простая диффузия. Вещества непосредственно, без чьей- либо помощи, диффундирует через липидный бислой из компартмента с большей концентрацией в компартмент с меньшей концентрацией (по градиенту концентрации) без затраты энергии АТФ. Путем простой диффузии через мембраны проникают главным образом неполярные гидрофобные вещества (в частности, кислород), легко растворимые в липидах. Облегченная диффузия. Сравнительно небольшие полярные молекулы, например, моносахариды и аминокислоты практически не проникают через мембрану большинства клеток за счет простой диффузии. Их перенос осуществляется путем облегченной диффузии. Облегченной называется диффузия вещества по градиенту его концентрации, которая осуществляется при участии специального транспортного интегрального белка мембранытранслоказы. 22.Транспорт через мембраны: эндоцитоз, экзоцитоз. Понятие о везикулярном транспорте. Перенос через мембрану частиц и высокомолекулярных соединений (эндоцитоз) • Эндоцитоз - перенос частиц в клетку. Его разновидности: пиноцитоз - захват и поглощение клеткой растворимых макромолекулярных соединений; фагоцитоз - то же самое, но в отношении твердых частиц; эндоцитоз, опосредованный рецепторами, - здесь поглощаемый субстрат предварительно специфически связывается с поверхностными рецепторами плазмолеммы. Это очень частый вариант фаго- и пиноцитоза, особенно в иммунных процессах. Во всех перечисленных случаях в месте проникновения субстрата вначале происходит впячивание плазмолеммы в цитоплазму. Затем оно все углубляется, пока не превращается в меммбранный пузырек, содержащий субстрат и полностью находящийся в цитоплазме. Перенос через мембрану частиц и высокомолекулярных соединений (экзоцитоз) • Экзоцитоз - перенос частиц и крупных соединений из клетки. Наиболее распространенный способ экзоцитоза - секреция. Это такое выведение из клетки растворимых соединений, которое является одной из функций данной клетки. Если из клетки удаляются твердые частицы, то такую разновидность экзоцитоза называют экскрецией. Примером экскреции может служить происходящее в конце эритропоэза удаление из ретикулоцитов сетчатой субстанции (агрегированных остатков органелл). Видимо, механизм экскреции вновь состоит в том, что в начале выделяемые частицы оказываются в цитоплазматическом пузырьке, который затем сливается с плазмолеммой. Существует еще одно понятие - рекреция. Это веществ через клетку; сочетаются фагоцитоз и перенос твердых фактически здесь экскреция. 23.Типы РНК в клетках. Функции РНК разных типов. Особенности пространственной организации тРНК.   24.Типы РНК в клетках. Функции РНК разных типов. Особенности пространственной организации тРНК, мРНК   25.Типы РНК в клетках. Функции РНК разных типов. Особенности пространственной организации рРНК. (рРНК) — несколько молекул РНК, составляющих основу рибосомы. Во вторичной структуре рРНК много двухцепочечных участков и петель. Основным назначением рРНК является осуществление трансляции — считывания информации с мРНК при помощи адапторных молекул тРНК и катализ образования пептидных связей между присоединёнными к тРНК аминокислотами.  |