Анатомия вопросы. Вопросы по Анатомии Строение животной клетки, основные органоиды и их функции. Строение клетки
 Скачать 2.52 Mb. Скачать 2.52 Mb.
|

|
По проводящим путям проходят импульсы в восходящем направлении от чувствительных и вставочных нейронов и в нисходящем — от клеток вышележащих нервных центров к двигательным нейронам. Задние канатики содержат восходящие пути, представленные тонким и клиновидным пучками. Они проводят к коре головного мозга сознательную проприоцептивную (мышечно-суставное чувство), кожную чувствительность (чувство стереогноза — узнавание предметов, имеющую отношение к определению положения тела в пространстве, и тактильную чувствительность. Боковые канатики содержат восходящие и нисходящие пути. Восходящие пути представлены задним и передним спиномозжечковыми путями, проводящими бессознательные проприоцептивные импульсы к мозжечку (бессознательная координация движения); спинопокрышечным и боковым спинобугорным путем (болевая и температурная чувствительность). К нисходящим путям относятся латерально-спинномозговой (пирамидный) путь, проводящий сознательные двигательные импульсы, и красноядерно-спинномозговой путь, проводящий непроизвольные двигательные импульсы. Передние канатики содержат нисходящие пути: - передний корковоспинномозговой (пирамидный), проводящий двигательные импульсы; - текто-спинномозговой, осуществляющий защитные движения при зрительных и слуховых раздражениях; - предцверно-спинномозговой, проводящий импульсы, обеспечивающие равновесие тела; - ретикулоспинномозговой. В спинном мозге замыкается большое количество рефлексов, регулирующих как соматические, так и вегетативные функции организма. Наиболее простые — это сухожильные рефлексы и рефлексы растяжения, имеющие моносинаптический характер. Более сложный характер имеют сгибательные рефлексы и рефлексы положения. Позные рефлексы направлены на поддержание определенной позы, что возможно лишь при наличии определенного мышечного тонуса. Кроме замыкания соматических рефлексов спинной мозг обеспечивает рефлекторную регуляцию внутренних органов, являясь центром висцеральных рефлексов. Спинной мозг имеет твердую, паутинную и мягкую соединительнотканные оболочки, продолжающиеся в такие же оболочки головного мозга. Твердая (наружная) мозговая оболочка обтекает его снаружи в виде мешка. Она не прилегает вплотную к стенкам позвоночного канала, которые покрыты надкостницей. Между надкостницей и твердой оболочкой находится эпидуральное пространство. В нем залегают жировая клетчатка и венозные сплетения. Вверху твердая оболочка срастается с краями большого отверстия затылочной кости, внизу на уровне II—III крестцовых позвонков суживается в виде нити и прикрепляется к копчику. Паутинная (средняя) мозговая оболочка в виде тонкого прозрачного бессосудистого листка прилегает изнутри к твердой оболочке. Между твердой и паутинной оболочками находится субдуральное пространство. Между паутинной и внутренней оболочкой находится подпаутинное пространство, в котором мозг и корешки лежат свободно и окружены большим количеством спинномозговой жидкости. Жидкость подпаутинного пространства спинного мозга непрерывно сообщается с жидкостью подпаутинных пространств головного мозга и мозговых желудочков. Мягкая (внутренняя) мозговая оболочка непосредственно обтекает спинной мозг. Между двумя своими листками она содержит сосуды, вместе с которыми входит в борозды и мозговое вещество спинного мозга. 39. Развитие головного мозга. Краниальный отдел нервной трубки в эмбриогенезе человека подразделяется на три мозговых пузыря (передний, средний и ромбовидный). В дальнейшем за счет складок и изгибов формируются пять отделов мозга: конечный и промежуточный мозг, средний сохраняется неделимым, ромбовидный подразделяется на продолговатый и задний мозг. Конечный мозг вначале непарный в дальнейшем формирует два полушария большого мозга. Производные среднего и ромбовидного мозговых пузырей образуют ствол мозга и являются древними образованиями, в которых сохраняется принцип сегментарности. Последний исчезает в производных промежуточного и конечного мозга, как образованиях филогенетически более молодых, в которых концентрируются интегративные функции. Стволовыми клетками для развития нервной ткани и нейроглии головного мозга служат матричные клетки, расположенные в эпендимном слое желудочков мозга. Матричные клетки интенсивно делятся митозом, мигрируют за пределы эпендимного слоя, дифференцируются на нейробласты и глиобласты (спонгиобласты), дающие позднее соответственно нейроны и клетки глии. Часть клеток остается на месте и формирует эпендимную выстилку желудочков мозга. Пролиферация и дифференцировка матричных клеток в разных отделах головного мозга происходят гетерохронно. При формировании коры важную роль в миграции нейробластов играют радиальные глиоциты (мюллеровы волокна). Тела клеток располагаются в эпендимном слое, а отроски простираются до наружной поверхности нейроэпителиальной выстилки нервной трубки. Именно по отросткам радиальных глиоцитов происходит направленная миграция нервных клеток из глубоких в поверхностные слои формирующихся корковых структур мозга. Из мезенхимы развиваются сосуды, а из стволовых кроветворных клеток — клетки микроглии. Ствол головного мозга К стволу головного мозга относят продолговатый мозг, мост, мозжечок и образования среднего и промежуточного мозга. По мере перехода спинного мозга в продолговатый теряется характерная форма серого вещества спинного мозга, но принцип локализации ядер по функции сохраняется: чувствительные ядра занимают дорсальную, а двигательные — вентральную части ствола. Между ними располагается ретикулярная формация. В чувствительных ядрах находятся нейроны, которые аналогичны не афферентным псевдоуниполярным нейронам спинального ганглия, а нейронам ядер задних рогов спинного мозга. Эти пучковые нейроны получают импульсы от нейронов сшшальных ганглиев по отросткам, идущим в составе клиновидного и тонкого пучков, а также от чувствительных нейронов таких ганглиев, как полулунный, коленный, каменистый, яремный, верхний пучковидный. В двигательных ядрах ствола мозга находятся мультиполярные нейроны, осуществляющие моторную иннервацию скелетных мышц головы и шеи. Нейроны вегетативных ядер продолговатого и среднего мозга направляют свои аксоны в вегетативные ганглии. В ассоциативных (переключательных) ядрах много нейронов, которые обеспечивают переключение импульсов из спинного мозга и ствола мозга на нейроны коры и в обратном направлении. Ствол головного мозга связан с периферическими органами чувствительными и двигательными волокнами черепных нервов. Аппарат черепных нервов вполне аналогичен собственному аппарату спинного мозга. Собственный внутренний аппарат ствола головного мозга обеспечивает связи между его частями. Рефлекторные дуги построены не из простой цепочки нейронов, а включают группы нейронов, в которых возможно осуществление пре- и постсинаптического торможения и модуляция с помощью нейропептидов интернейронов потока афферентных и эфферентных сигналов. Группировки нейронов являются общим принципом внутренней организации корковых и ядерных структур мозга. Объединять нейроны в единую функциональную систему могут коллатерали афферентного отростка и интернейроны (продуцирующие нейропептиды), модифицирующие проведение нервного импульса. Различные части ствола головного мозга тесно взаимосвязаны благодаря наличию внутреннего собственного рефлекторного аппарата. Важная роль в установлении этих взаимосвязей принадлежит также аппарату двусторонних связей спинного мозга и стволовой части головного мозга, который включает восходящие и нисходящие пути. В состав ствола мозга входит ретикулярная формация — восходящая диффузно активирующая система головного мозга. В ее сети располагаются мультиполярные нейроны (размером от 5 до 120 мкм) с маловетвящимися отростками. Ретикулярная формация получает импульсы от афферентных путей, но сами импульсы проходят через нее в 4-5 раз медленнее, чем через прямые пути. Отростки нейронов ретикулярной формации направляются в кору большого мозга, мозжечка, в ядра ствола мозга, где формируют синапсы (холин-, адрен-, дофаминэргические и др.) с нейронами. Так осуществляется интегративная функция ретикулярной формации. Нисходящие волокна нейронов ретикулярной формации взаимодействуют с моторными нейронами спинного мозга. При этом они тормозят их активность. Считается, что ретикулярная формация принимает участие в формировании эмоций, восприятии боли, осуществляет контроль стереотипных движений, тонуса мышц. 40. Ромбовидный мозг: продолговатый мозг, мост, мозжечок. Их строение и функции Продолговатый мозг – является непосредственным продолжением спинного мозга. Он сочетает в себе черты строения спинного мозга и начального отдела головного мозга. На его передней поверхности по средней линии проходит передняя срединная щель, являющаяся продолжением одноименной борозды спинного мозга. По бокам от щели находятся пирамиды, которые продолжаются в передние канатики спинного мозга. Пирамиды состоят из пучков нервных волокон, которые в борозде перекрещиваются с такими же волокнами противоположной стороны. Латеральнее пирамид с двух сторон находятся возвышения – оливы. На задней поверхности продолговатого мозга проходит задняя (дорсальная) срединная борозда, являющаяся продолжением одноименной борозды спинного мозга. По бокам от борозды лежат задние канатики. В них проходят восходящие пути спинного мозга. По направлению кверху задние канатики расходятся в стороны и идут к мозжечку. Внутреннее строение продолговатого мозга. Продолговатый мозг состоит из серого и белого вещества. Серое вещество представлено скоплениями нейронов, расположено оно внутри в виде отдельных скоплений ядер. Различают: 1) собственные ядра – это ядро оливы, имеющее отношение к равновесию, координации движений. 2) ядра ЧМН с IX по XII пару. Также в продолговатом мозге находится ретикулярная формация, образующаяся из переплетений нервных волокон и лежащих между ними нервных клеток. Белое вещество продолговатого мозга находится снаружи, содержит длинные и короткие волокна. Короткие волокна осуществляют связь между ядрами самого продолговатого мозга и между ядрами ближайших отделов головного мозга. Длинные волокна образуют проводящие пути – это восходящие чувствительные пути идущие от продолговатого мозга к таламусу и нисходящие пирамидные пути, проходящие в передние канатики спинного мозга. Функции продолговатого мозга. 1. Рефлекторная функция связана с центрами находящимися в продолговатом мозге. В продолговатом мозге расположены следующие центры: 1) Дыхательный центр, обеспечивающий вентиляцию легких; 2) Пищевой центр, регулирующий сосание, глотание, отделение пищеварительного сока (слюноотделения, желудочного и поджелудочного соков); 3) Сердечно-сосудистый центр – регулирующий деятельность сердца и кровеносных сосудов. 4) Центр защитных рефлексов – это мигание, слюноотделение, чихание, кашель, рвота. 5) Центр лабиринтных рефлексов, осуществляющий распределение мышечного тонуса между отдельными группами мышц и установочные рефлексы позы. 2. Проводниковая функция связана с проводящими путями. Через продолговатый мозг проходят восходящие пути от спинного мозга к головному мозгу и нисходящие пути, связывающие кору больших полушарий со спинным мозгом. Мозжечок (Cerebellum) В мозжечке различают два полушария и непарную среднюю часть – червь мозжечка. Мозжечок состоит из серого и белого вещества. Серое вещество находится снаружи и образует кору мозжечка. Кора представлена тремя слоями нервных клеток. Белое вещество находится внутри и состоит из нервных волокон. На разрезе белое вещество напоминает ветвистое дерево, отсюда и его название «дерево жизни». Волокна белого вещества идут в составе трех пар мозжечковых ножек. Верхние ножки связывают мозжечок со средним мозгом. Средние ножки связывают мозжечок с мостом. Нижние ножки связывают мозжечок с продолговатым мозгом. В толще белого вещества находятся отдельные парные скопления нервных клеток, которые образуют ядра мозжечка: зубчатое, шаровидное, пробковидное и ядро шатра. Функции мозжечка: 1) Координация позы и целенаправленных движений. 2) Регуляция позы и мышечного тонуса. 3) Координация быстрых целенаправленных движений. 4) Регуляция вегетативных функций (изменение работы сердца и сосудов, расширение зрачка). При повреждении мозжечка наблюдается симптом мозжечковой атаксии. Больные с таким симптомом ходят с широко расставленными ногами, совершают лишние движения, покачиваются из стороны в сторону. В клинике этот симптом называют симптомом «пьяного человека». При частичном поражении мозжечка наблюдаются три основных симптома: атония, астения и астазия. Атония характеризуется ослаблением мышечного тонуса. Астения характеризуется слабостью и быстрой усталостью мышц. Астазия проявляется в способности мышц выполнять колебательные и дрожательные движения. Мост Располагается мост между продолговатым мозгом и ножками мозга, а по бокам переходит в средние мозжечковые ножки (рис. 1). Со стороны основания мозга мост представляет собой плотный белый вал размерами 30 X 36 X 25 мм. Передняя поверхность моста выпуклая, обращена вперед и вниз и прилежит на основании черепа к скату. Посередине передней поверхности проходит базилярная борозда (sulcus basilaris), в к-рой лежит базилярная артерия (a. basilaris), являющаяся основным источником кровоснабжения Моста головного мозга. Позади моста головного мозга из борозды между продолговатым мозгом, с одной стороны, мостом и средней мозжечковой ножкой — с другой, последовательно выходят корешки отводящего, лицевого, промежуточного и преддверно-улиткового нервов. Задняя поверхность моста обращена кверху и кзади, в полость четвертого желудочка, и снаружи не видна, т. к. прикрыта мозжечком. Она образует верхнюю половину дна ромбовидной ямки. На поперечных (фронтальных) срезах Моста головного мозга (рис. 2) различают более массивную переднюю (вентральную) часть (pars ant. pontis), или основание (basis pontis, BNA), и небольшую заднюю (дорсальную) часть (pars post, pontis), или покрышку (tegmentum, BNA). Границей между ними служит трапециевидное тело (corpus trapezoideum), образованное преимущественно отростками клеток переднего улиткового ядра (nucleus cochlearis ant.). Скопления нервных клеток образуют переднее и заднее ядра трапециевидного тела (ядра Гуддена). Передняя часть моста содержит гл. обр. нервные волокна, между к-рыми разбросаны многочисленные небольшие скопления серого вещества — ядра моста (nuclei pontis). В ядрах моста заканчиваются волокна корково-мостового пути (tractus corticopontini) и коллатерали от проходящих пирамидных путей. Отростки клеток ядер моста образуют мостомозжечковый путь, волокна к-рого переходят преимущественно на противоположную сторону и являются поперечными волокнами моста (fibrae pontis transversae). Последние образуют средние мозжечковые ножки (pedunculi cerebellares medii). Задняя часть моста (покрышка) значительно тоньше. Она содержит ретикулярную формацию (formatio reticularis) и ядра V, VI, VII, VIII пар черепных нервов. На уровне середины моста располагается двигательное ядро тройничного нерва (nucleus motorius n. trigemini), а несколько латеральнее — верхнее чувствительное ядро (nucleus sensorius sup.). К последнему подходят волокна от чувствительных клеток тройничного узла, к-рые в составе чувствительного корешка вступают в вещество моста на границе его со средней мозжечковой ножкой. К чувствительному корешку прилежит двигательный корешок, представляющий собой отростки клеток двигательного ядра тройничного нерва. На уровне лицевого бугорка располагается ядро отводящего нерва; рядом, в ретикулярной формации, находится двигательное ядро лицевого нерва, отростки клеток к-рого формируют колено, огибающее ядро отводящего нерва. Позади двигательного ядра лицевого нерва находится верхнее слюноотделительное ядро (nucleus salivatorius sup.) и кнаружи от последнего — ядро одиночного пути (nucleus tractus solitarii). В нижнелатеральном отделе покрышки моста расположены ядра преддверно-улиткового нерва (п. vestibulocochlearis). По сторонам трапециевидного тела находятся верхние оливы. Отростки клеток верхней оливы (oliva sup.) составляют латеральную петлю (lemniscus lat.), между волокнами последней располагается ядро латеральной петли . (nucleus lemnisci lat.). В состав латеральной петли входят также отростки клеток заднего ядра улиткового нерва (nuci, cochlearis post.), ядер трапециевидного тела и ядра латеральной петли. Кнутри от верхней оливы над трапециевидным телом расположена медиальная петля (lemniscus med.), представляющая собой пучок волокон проприоцептивной чувствительности, и спинальная петля (lemniscus spinalis) — пучок волокон пути болевой и температурной чувствительности. Функции Важное функциональное значение Моста головного мозга обусловлено, с одной стороны, расположением в нем ядер черепных нервов (V, VI, VII, VIII пар), ретикулярной формации, ядер моста, с другой — прохождением через Мост эфферентных путей (корковоспинномозгового и корково-ядерного, покрышечно-спинномозгового, красноядерно-спинномозгового, ретикулярно-спинномозгового и др.) и афферентных путей (спиноталамического, проводящих путей проприоцептивной — глубокой — чувствительности и др.), имеющих для организма жизненно важное значение и осуществляющих двустороннюю связь между головным мозгом (см.) и спинным мозгом (см.). 41. Средний мозг, строение, функции. Средний мозг состоит из ножек мозга и крыши мозга. Они разделены сильвиевым водопроводом мозга, который соединяет третий и четвертый желудочки головного мозга. Ножки мозга состоят из основания и покрышки, между которыми располагаются пигментированные клетки черной субстанции. Черная субстанция участвует в сложной координации движений. Основание ножек образует пирамидный путь. В покрышке ножек лежат ядра блокового и глазодвигательного нервов (III и IV пара черепных нервов). Также в ней располагается красное ядро, в котором заканчиваются верхние ножки мозжечка. В них идет восходящий путь к зрительному бугру и нисходящий — красноядерно-спинномозговой. Красное ядро отвечает за поддержание тонуса мускулатуры туловища и конечностей. Четверохолмие, или крыша мозга, составляет заднюю часть среднего мозга. Перпендикулярными друг другу бороздами оно делится на верхние и нижние холмики. Верхнее двухолмие заключает в себе центры ориентировочных рефлексов на зрительные раздражения. Посредством отходящих вперед ручек холмики соединяются с латеральными коленчатыми телами промежуточного мозга. По этим ручкам идут волокна зрительного нерва. Нижнее двухолмие служит центром ориентировочных рефлексов на слуховые раздражения. От холмиков к медиальным коленчатым телам идут нижние ручки, по которым проходят волокна слухового нерва. Ядра четверохолмия играют важнейшую роль в раннем онтогенезе, обеспечивая первичные формы сенсорного внимания. В среднем мозге замыкается ряд рефлексов. Нейроны бугров четверохолмия отвечают за ориентировочные зрительные и слуховые рефлексы. Ядра четверохолмия участвуют в осуществлении сторожевого рефлекса, что выражается в усилении тонуса сгибателей. Черная субстанция обеспечивает сложную координацию движений. В ней находятся содержащие дофамин нейроны, регулирующие эмоциональное поведение. Повреждение черной субстанции приводит к нарушению тонких движений пальцев рук, развитию тремора (болезнь Паркинсона). Красное ядро отвечает за тонус мышц-сгибателей. 42. Промежуточный мозг, строение и функции. В промежуточном мозге различают парные зрительные бугры (таламус), латеральные и медиальные коленчатые тела, подбугорную (гипоталамус) и надбугорную (эпиталамус) области. Зрительный бугор (таламус) представляет собой крупное тело овальной формы. Он состоит из серого вещества, группирующегося в ядра. Все ядра делятся на специфические и неспецифические. Специфические ядра получают информацию от определенных видов рецепторов и посылают их в строго определенные зоны коры. Ядра, переключающие информацию на центральные поля анализаторов, относят к проекционным, или релейным. Ядра, передающие информацию на ассоциативные области, являются ассоциативными. Неспецифические ядра представлены ретикулярной формацией. Они располагаются вокруг специфических, диффузно влияют на кору и подкорковые ядра и могут вызывать как возбуждающий, так и тормозной эффект. Эти ядра не выполняют высших интегративных функций, но участвуют в регуляции афферентных влияний. К моменту рождения большая часть ядер зрительных бугров хорошо развита. После рождения их размеры увеличиваются за счет роста нервных клеток и развития нервных волокон. Все сенсорные сигналы, за исключением обонятельных, достигают коры больших полушарий только через таламокортикальные проекции. Таламус представляет собой ворота, через которые в кору поступает информация о состоянии нашего тела и окружающем мире. Афферентные сигналы на пути к коре мозга переключаются на нейронах таламуса, что позволяет обеспечить передачу в кору мозга наиболее важной информации. Система неспецифических ядер таламуса контролирует ритмическую активность коры больших полушарий и выполняет функции внутриталамической интегрирующей системы. Таламус является высшим центром болевой чувствительности. Повреждение неспецифических ядер таламуса приводит к нарушению сознания. Это свидетельствует о том, что импульсация, поступающая по неспецифической восходящей системе таламуса, поддерживает уровень возбудимости корковых нейронов, необходимый для сохранения сознания. Кроме того, таламус является надсегментарным центром рефлекторной деятельности. Латеральное коленчатое тело располагается кнаружи от корешка зрительного пути. Медиальное коленчатое тело лежит на уровне поперечной борозды четверохолмия. Волокна нервных клеток коленчатых тел в составе зрительных и слуховых путей направляются к коре больших полушарий. Гипоталамус хорошо заметен на основании головного мозга. В задней его области располагаются два сосцевидных тела. Волокна этих тел образуют сосково-бугорный путь, по которому импульсы идут к передним ядрам зрительного бугра. Сосцевидные тела, как и передние ядра зрительных бугров, относят к лимбической системе, которая отвечает за организацию поведенческих реакций. Спереди от сосцевидных тел лежит серый бугор. Суживаясь, он переходит в воронку, проникающую в ямку турецкого седла через его диафрагму. На воронке подвешен гипофиз. Серый бугор является центром автономной нервной системы, которая влияет на сохранение гомеостаза организма и на его приспособление к условиям внешней среды. Впереди серого бугра зрительные нервы образуют перекрест (хиазму), после которого получают название зрительных путей. Над перекрестом лежит супраоптическое ядро. Его клетки вырабатывают нейросекреты, проникающие в заднюю долю гипофиза. Этими веществами являются антидиуретический гормон, регулирующий водный метаболизм, и окситоцин, влияющий на деятельность матки. По-иному, т.е. нейрогуморальным путем, через кровь, осуществляется связь гипоталамуса с передней долей гипофиза, вырабатывающей такие гормоны, как адренокортикотропный, фолликулостимулирующий и лютеинизирующий, тиреотропный, гормон роста. Таким образом, здесь образуется гипоталамо-гипофизарная система, где объединяются два уровня регуляции функций организма человека — нервная и гуморальная. Дифференцировка ядер гипоталамуса к моменту рождения не завершена и протекает в онтогенезе неравномерно. Развитие ядер заканчивается в период полового созревания. В функциональном отношении ядра гипоталамуса неоднородны. Латеральная и дорсальная группы ядер повышают тонус симпатической нервной системы, средние ядра (серый бугор) — снижают его. В гипоталамусе располагаются центр сна и центр пробуждения, он участвует в процессе чередования сна и бодрствования. Гипоталамус играет важную роль в терморегуляции. Раздражение задних ядер приводит к гипертермии в результате повышения теплопродукции. В области средних и боковых ядер располагаются центры насыщения и голода, которые активируются в результате изменения химического состава протекающей крови. Дорсолатерально от супраоптического ядра находится центр жажды. Активация его приводит к увеличению потребления воды (полидипсия), а разрушение сопровождается отказом от воды (адипсия). В гипоталамусе расположены центры, связанные с регуляцией полового поведения, названные центрами удовольствия. Они являются компонентом нейронной системы, участвующей в регуляции эмоциональной сферы полового поведения. В результате связей гипоталамуса с гипофизом образуется гипоталамо-гипофизарная система. Надбугорная область (эпиталамус) связана с обонятельной системой. Эпиталамус участвует в образовании стенок третьего желудочка головного мозга и состоит из мозговых полосок, сзади расширяющихся в поводковые треугольники. От последних отходят поводки (белые тяжи), которые соединяют эпиталамус с эпифизом. В треугольниках 12.2. Строение, функции и возрастные особенности отделов ЦНС 315 лежат поводковые ядра, отдающие нисходящие волокна к ядрам среднего мозга. Промежуточный мозг у новорожденного развит относительно хорошо. Внутри промежуточного мозга находится третий желудочек головного мозга, имеющий вид вертикальной щели, ограниченной с боков медиальными поверхностями зрительных бугров, снизу гипоталамусом, спереди — столбами свода, сзади — эпиталамусом, сверху — сводом. Между зрительными буграми расположены межжелудочковые отверстия, которые соединяют полость третьего желудочка с боковыми желудочками больших полушарий. Развитие структур промежуточного мозга состоит в увеличении их взаимосвязей с другими мозговыми образованиями, что создает условия для совершенствования координационной деятельности его различных отделов. В развитии промежуточного мозга существенная роль принадлежит нисходящим влияниям коры больших полушарий. 43. Полушария большого мозга: строений (извилины, борозды, доли, зоны). Анатомическая и функциональная асимметрия правого и левого полушария. 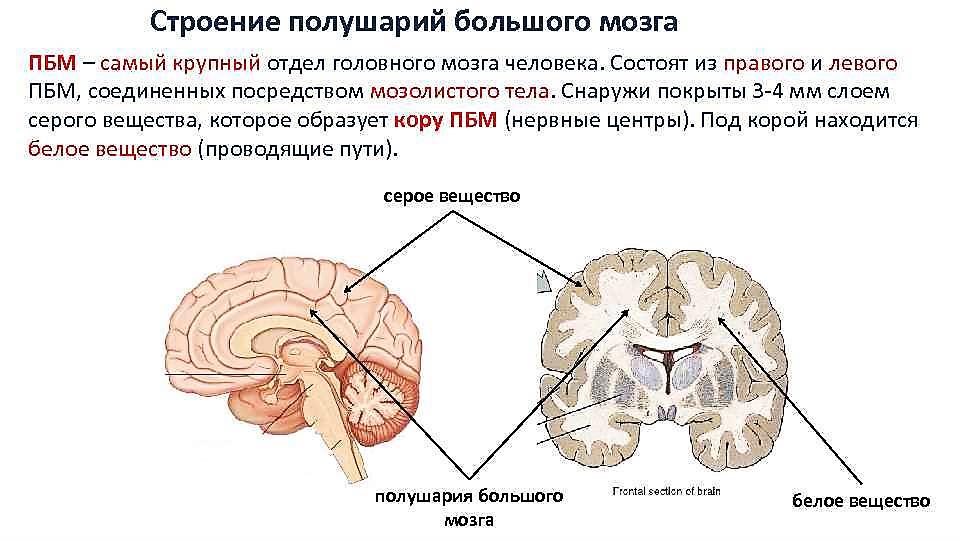 Головноймозгсостоитиз переднего мозга (конечного и промежуточного), среднего мозга, ромбовидного мозга (задний мозг – мост, мозжечок, продолговатый мозг). Между ромбовидным и средним мозгом расположен перешеек ромбовидного мозга. Головной мозг находится в полости мозгового черепа. Имеет выпуклую верхнелатеральную поверхность и нижнюю поверхность и уплощенную – основание головного мозга. Масса мозга взрослого человека составляет от 1100 до 2000 грамм; от 20 до 60 лет масса m и объем V остаются максимальными и постоянными, после 60 лет несколько уменьшается. Ни абсолютная, ни относительная масса мозга не является показателем степени умственного развития. Масса мозга Тургенева была 2012 г, Байрона – 2238 г, Кювье – 1830 г, Шиллера – 1871 г, Менделеева – 1579 г, Павлова – 1653 г. Головной мозг состоит из тел нейронов, нервных трактов и кровеносных сосудов. Головной мозг состоит из 3 частей: полушарий большого мозга, мозжечка и мозгового ствола. Полушария большого мозга достигают максимального развития у человека, позднее других отделов. Большой мозг состоит из двух полушарий – правого и левого, которые связаны одно с другим толстой спайкой (комиссурой) – мозолистым телом. Правое и левое полушария делятся с помощью продольной щели. Под комиссурой находится свод, представляющий собой два изогнутых волокнистых тяжа, которые в средней части соединены между собой, а спереди и сзади расходятся, образуя столбы и ножки свода. Спереди от столбов свода находится передняя спайка. Между мозолистым телом и сводом натянута тонкая вертикальная пластинка мозговой ткани – прозрачная перегородка. Полушария имеют: - верхнелатеральную, - медиальную, - нижнюю поверхности. Верхнелатеральная – выпуклая, медиальная – плоская, обращенная к такой же поверхности другого полушария, и нижняя – неправильной формы. На трех поверхностях располагаются глубокие и мелкие борозды, и между ними извилины. Борозды – углубления между извилинами. Извилины – возвышения мозгового вещества. Поверхности полушарий большого мозга отделены друг от друга краями – верхним, нижнелатеральным и нижневертикальным. В пространстве между двумя полушариями входит серп большого мозга – большой серповидный отросток, представляющий собой тонкую пластинку твердой оболочки, которая проникает в продольную щель большого мозга, не достигая мозолистого тела, и отделяет друг от друга правое и левое полушария. Наиболее выступающие участки полушарий получили название полюсов: лобного, затылочного и височного. Рельеф поверхностей полушарий большого мозга очень сложен и связи с наличием более или менее глубоких борозд большого мозга и расположенных между ними валикообразных возвышений – извилин. Глубина, протяженность некоторых борозд и извилин, их форма и направление очень изменчивы. Каждое полушарие делится на доли – лобную, теменную, затылочную, височную, островковую. Центральная борозда (Роландова борозда) отделяет лобную долю от теменной, латеральная борозда (Сильвиева борозда) – височную от лобной и теменной, теменно-затылочная разделяет теменную и затылочную доли. Латеральная борозда закладывается к 4-му месяцу внутриутробного развития, теменно-зытылочная и центральная – к 6-му месяцу. Во внутриутробном периоде происходит гирификация – формирование извилин. Эти три борозды возникают первыми и отличаются большой глубиной. Вскоре к центральной борозде прибавляется еще пара ей параллельных: одна проходит впереди центральной и соответственно называется предцентральной, которая распадается на две – верхнюю и нижнюю. Другая борозда располагается позади центральной и называется постцентральной. Постцентральная борозда лежит позади центральной борозды и почти параллельно ей. Между центральной и постцентральной бороздами располагается постцентральная извилина. Вверху она переходит на медиальную поверхность полушария большого мозга, где соединяется с предцентральной извилиной лобной доли, образуя вместе с нею парацентральную дольку. На верхнелатеральной поверхности полушария, внизу постцентральная извилина также переходит в предцентральную извилину, охватывая снизу центральную борозду. Она параллельна верхнему краю полушария. Кверху от внутритеменной борозды находится группа мелких извилин, получивших название верхней теменной дольки. Ниже этой борозды лежит нижняя теменная долька, в пределах которой выделяют две извилины: надкраевую и угловую. Надкраевая извилина охватывает конец латеральной борозды, а угловая – конец верхней височной борозды. Нижняя часть нижней теменной дольки и прилежащие к ней нижние отделы постцентральной извилины месте с нижней частью предцентральной извилины, нависающие над островковой долей, образуют лобно-теменную покрышку островка. Доли мозга – дорсальную и латеральную поверхность коры головного мозга – принято делить на четыре доли, которые получили наименование от соответствующих костей черепа: лобная, теменная, затылочная, височная. Затылочная доля располагается позади теменно-затылочной борозды и ее условного продолжения на верхнелатеральной поверхности полушария. По сравнению с другими долями она имеет небольшие размеры. Кзади затылочная доля кончается затылочным полюсом. Борозды и извилины на верхнелатеральной поверхности затылочной доли очень вариабельны. Наиболее часто и лучше других выражена поперечная затылочная борозда которая является как бы продолжением кзади внутритеменной борозды теменной доли мозга. Височная доля занимает нижнебоковые отделы полушария и отделяется от лобной и теменной долей глубокой латеральной бороздой. Край височной доли, прикрывающий островковую долю, получил название височной покрышки островка. Передняя часть височной доли образует височный полюс. На боковой поверхности височной доли видны две борозды: верхняя и нижняя височные, почти параллельные латеральной борозде. Извилины височной доли ориентированы вдоль борозд. Верхняя височная извилина расположена между латеральной бороздой вверху и верхней височной внизу. На верхней поверхности этой извилины, скрытой в глубине латеральной борозды, располагаются 2-3 короткие поперечные височные извилины (извилины Гешля), разделенные поперечными височными бороздами. Между верхней и нижней височными бороздами находится средняя височная извилина. Нижнелатеральный край височной доли занимает нижняя височная извилина ограниченная сверху одноименной бороздой. Задний конец этой извилины продолжается в затылочную долю. Островковая доля (островок) находится в глубине латеральной борозды. Эту долю можно увидеть, если раздвинуть или удалить прикрывающие островок участки: лобной, теменной и височной долей, которые получили наименование покрышки. Глубокая круговая борозда островка отделяет островок от окружающих его отделов мозга. Поверхность островка представлена длинной и короткими извилинами. Между длинной извилиной, находящейся в задней части островка и ориентированной сверху вниз и вперед, и короткими извилинами, занимающими верхнюю часть островка, находится центральная борозда островка. Нижнепередняя часть островка лишена борозд, имеет небольшое утолщение, получившее название «порог островка». Медиальная поверхность полушария. Все доли полушария, за исключением островковой, принимают участие в образовании его медиальной поверхности (Рис. 5). Над мозолистым телом, отделяя его от остальных отделов полушария, находится борозда мозолистого тела. Огибая сзади валик мозолистого тела, она направляется книзу и вперед и продолжается в борозду гиппокампа или гиппокампальную борозду. Выше борозды мозолистого тела находится поясная борозда. Эта борозда начинается кпереди и книзу от клюва мозолистого тела, поднимается вверх, затем поворачивает назад и следует параллельно борозде мозолистого тела, заканчиваясь выше и кзади от валика мозолистого тела под названием подтеменной борозды. На уровне валика мозолистого тела от поясной борозды вверх ответвляется краевая часть, уходящая вверх и кзади к верхнему краю полушария большого мозга. Между бороздой мозолистого тела и поясной бороздой находится поясная извилина, охватывающая мозолистое тело спереди, сверху и сзади. Сзади и книзу от валика мозолистого тела поясная извилина сужается, образуя перешеек поясной извилины. Далее книзу и кпереди перешеек переходит в более широкую парагиппокампальную извилину, ограниченную сверху бороздой гиппокампа. Поясная извилина, перешеек и парагиппокампальная извилина известны как сводчатая извилина. В глубине борозды гиппокампа находится довольно тонкая полоска серого цвета, разделенная мелкими поперечными бороздками, – зубчатая извилина. Участок медиальной поверхности полушария, находящийся между поясной бороздой и верхним краем полушария, относится к лобной и теменной долям. К переди от верхнего края центральной борозды находится медиальная поверхность верхней лобной извилины, а непосредственно к указанному участку центральной борозды прилегает парацентральная долька, ограниченная сзади краевой частью поясной борозды. Между краевой частью спереди и теменно-затылочной бороздой сзади находится предклинье – принадлежащий теменной доле участок полушария большого мозга. На медиальной поверхности затылочной доли расположены сливающиеся друг с другом под острым углом, открытым кзади, две глубокие борозды. Это теменно-затылочная борозда, отделяющая теменную долю от затылочной, и шпорная борозда, начинающаяся на медиальной поверхности затылочного полюса и направляющаяся вперед до перешейка поясной извилины. Участок затылочной доли, лежащий между теменно-затылочной и шпорной бороздами и имеющий форму треугольника, обращенного вершиной к месту слияния этих борозд, называется «клином». Хорошо заметная на медиальной поверхности полушария шпорная борозда ограничивает сверху язычную извилину, простирающуюся от затылочного полюса сзади до нижней части перешейка поясной извилины. Снизу от язычной извилины располагается коллатеральная борозда, принадлежащая уже нижней поверхности полушария. Нижняя поверхность полушария. Рельеф нижней поверхности полушария очень сложен (Рис. 6). Передние отделы нижней поверхности образованы лобной долей полушария, позади которой выступает височный полюс, а также находятся нижние поверхности височной и затылочной долей, переходящие одна в другую без заметных границ. На нижней поверхности лобной доли, несколько латеральнее и параллельно продольной щели большого мозга, находится обонятельная борозда. Снизу к ней прилегают обонятельная луковица и обонятельный тракт, переходящий сзади в обонятельный треугольник, в области которого видны медиальная и латеральная обонятельные полоски. Участок лобной доли между продольной щелью большого мозга и обонятельной бороздой получил название прямой извилины. Поверхность лобной доли, лежащая латеральнее от обонятельной борозды, разделена неглубокими глазничными бороздами на несколько вариабельных по форме, расположению и размерам глазничных извилин. В заднем отделе нижней поверхности полушария хорошо видна коллатеральная борозда, лежащая книзу и латерально от язычной извилины на нижней поверхности затылочной и височной долей, латерально от парагиппокампальной извилины. Несколько кпереди от переднего конца коллатеральной борозды находится носовая борозда, ограничивающая с латеральной стороны изогнутый конец парагиппокампальной извилины – крючок. Латеральнее коллатеральной борозды лежит медиальная затылочно-височная извилина. Между этой извилиной и расположенной кнаружи от нее латеральной затылочно-височной извилиной находится затылочно-височная борозда. Границей между латеральной затылочно-височной и нижней височной извилинами служит не борозда, а нижнелатеральный край полушария большого мозга. Верхнелатеральная поверхность полушария – находящаяся в переднем отделе каждого полушария большого мозга лобная доля, оканчивающаяся спереди лобным полюсом и ограничивающая снизу латеральной (сильвиевой) бороздой, а сзади – глубокой центральной бороздой. Ряд отделов головного мозга, расположенных преимущественно на медиальной поверхности полушария и являющихся субстратом для формирования таких общих состояний, как бодрствование, сон, эмоции и др., выделяют под названием «лимбическая система». Поскольку эти реакции сформировались в связи с первичными функциями обоняния (в филогенезе), их морфологической основой являются отделы мозга, которые развиваются из нижних отделов мозгового пузыря и относятся к так называемому обонятельному мозгу. Лимбическую систему составляют обонятельная луковица, обонятельный тракт, обонятельный треугольник, переднее продырявленное вещество, которые расположены на нижней поверхности лобной доли (периферический отдел обонятельного мозга), а также поясная и парагиппокампальная (вместе с крючком) извилины, зубчатая извилина, гиппокамп (центральный отдел обонятельного мозга) и некоторые другие структуры. Включение этих отделов мозга в лимбическую систему оказалось возможным в связи с общими чертами их строения (и происхождения), наличием взаимных связей и сходством функциональных реакций. Полушария состоят из серого и белого вещества. Слой серого вещества называется корой головного мозга. Кора покрывает в виде плаща остальные образования большого мозга и поэтому и называется плащом. Под корой белое вещество, а в нем островки серого вещества – базальные ядра, в основном расположенные в лобной доле. К ним относят полосатое тело (хвостатое ядро и чечевицеобразное ядро), ограду и миндалевидное тело. Полосатое тело (стриопаллидарная система) состоит из 2-х ядер – хвостатого и чечевицеобразного, разделенных прослойкой белого вещества – внутренней капсулой. В эмбриональном периоде полосатое тело составляет одну серую массу, затем оно разделяется. Хвостатое ядро расположено около таламуса. Оно имеет подковообразную форму, состоит из головки, тела и хвоста. Чечевицеобразное ядро имеет форму чечевичного зерна, находится латеральнее таламуса и хвостатого ядра. Чечевицеобразное ядро делится на 3 части, благодаря белому веществу. Наиболее латерально лежит скорлупа, имеющая темную окраску, а две более светлые части называются латеральным и медиальным бледными шарами. Для скорлупы характерно участие в организации пищевого поведения: пищепоиска, пищенаправленности, пищезахвата и пищевладения; ряд трофических нарушений кожи, внутренних органов возникает при нарушениях функции скорлупы. Раздражения скорлупы приводят к изменениям дыхания, слюноотделения. Бледный шар имеет преимущественно крупные нейроны Гольджи 1-го типа. Связи бледного шара с таламусом, скорлупой, хвостатым ядром, средним мозгом, гипоталамусом, соматосенсорной системой и др. свидетельствуют о его участии в организации простых и сложных форм поведения. Раздражение бледного шара с помощью вживленных электродов вызывает сокращение мышц конечностей, активацию или торможение γ-мотонейронов спинного мозга. У больных с гиперкинезами раздражение разных отделов бледного шара (в зависимости от места и частоты раздражения) увеличивает или снижает гиперкинез. Стимуляция бледного шара, в отличие от стимуляции хвостатого ядра, не вызывает торможения, а провоцирует ориентировочную реакцию, движения конечностей, пищевое поведение (обнюхивание, жевание, глотание и т.д.). Повреждение бледного шара вызывает у людей гипомимию, маскообразность лица, тремор головы, конечностей (причем этот тремор исчезает в покое, во сне и усиливается при движениях), монотонность речи. При повреждении бледного шара наблюдается миоклония – быстрые подергивания мышц отдельных групп или отдельных мышц рук, спины, лица. Ядра полосатого тела конечного мозга являются подкорковыми двигательными центрами в составе экстрапирамидной системы, регулирующими сложные автоматизированные двигательные акты. К экстрапирамидной системе относят черное вещество и красные ядра ножек мозга. Полосатое тело управляет процессами теплорегуляции и обмена углеводов. К наружи от чечевицеобразного ядра расположена тонкая пластинка серого вещества – ограда. Ограда находится в белом веществе полушария сбоку от скорлупы, между последней и корой островковой доли. Она содержит полиморфные нейроны разных типов и образует связи преимущественно с корой большого мозга. Глубокая локализация и малые размеры ограды представляют определенные трудности для ее физиологического исследования. Миндалевидное тело (большая спайка мозга) находится в переднем отделе височной доли, входит в состав лимбической системы. К белому веществу полушария относятся внутренняя капсула и волокна, проходящие спайки (мозолистое тело, передняя спайка, спайка свода) и направляющиеся к коре и базальным ядрам. Внутренняя капсула – толстая изогнутая пластинка белого вещества. Она делится на три отдела: 1) переднюю ножку внутренней капсулы, 2) заднюю ножку внутренней капсулы, 3) место соединения этих двух отделов – колено внутренней капсулы. В колене внутренней капсулы располагаются корково-ядерные пути, идущие к двигательным ядрам черепных нервов. В переднем отделе располагаются корково-спинномозговые волокна, находящиеся в предцентральной извилине и идущие к двигательным ядрам передних рогов спинного мозга. В задней ножке располагаются таламокортикальные волокна, идущие в кору постцентральной извилины. В состав этого проводящего пути соединяются волокна проводников всех видов общей чувствительности (большой температуры, осязания, давления, проприоцентивной). В задних отделах задней ножки располагаются слуховой и зрительный проводящие пути. Оба берут начало от подкорковых центров слуха и зрения и заканчиваются в соответствующих центрах. Таким образом, базальные ядра головного мозга являются интегративными центрами организации моторики, эмоций, высшей нервной деятельности, причем каждая из этих функций может быть усилена или заторможена активацией отдельных образований базальных ядер. Мозолистое тело представляет собой толстую изогнутую пластинку, состоящую из поперечных волокон. В мозолистом теле разделяют: колено, клюв, между ними ствол, который переходит в валик. Волокна, проходящие в колене, соединяют кору лобных долей правого и левого полушарий. Волокна ствола соединяют серое вещество теменных и височных долей. В валике проходящие волокна соединяют кору затылочных долей. Под мозолистым телом располагается свод, который состоит из двух дугообразно изогнутых тяжей, соединенных при помощи спайки. Свод состоит из тела, парного столба и парных ножек. Ножки, срастаясь с гипокампом, образуют бахромку. Боковой желудочек – полость полушарий (I и II желудочки) и сообщающяеся через межжелудочковое отверстие с III желудочком. В каждом желудочке выделяют центральную часть и три рога: передний (лобный) – в лобной доле; задний (затылочный) – в затылочной доле и нижний (височный) – в височный доле. Боковые желудочки, как и другие желудочки головного мозга, и центральный канал спинного мозга изнутри выстланы слоем эпендимоцитов. Эпендимные клетки образуют спинномозговую жидкость и регулируют ее состав. Ромбовидная ямка представляет собой ромбовидной формы вдавление, длинная ось которой направлена вдоль мозга. Она ограничена с боков в своем верхнем отделе верхними мозжечковыми, в нижнем – нижними мозжечковыми ножками. 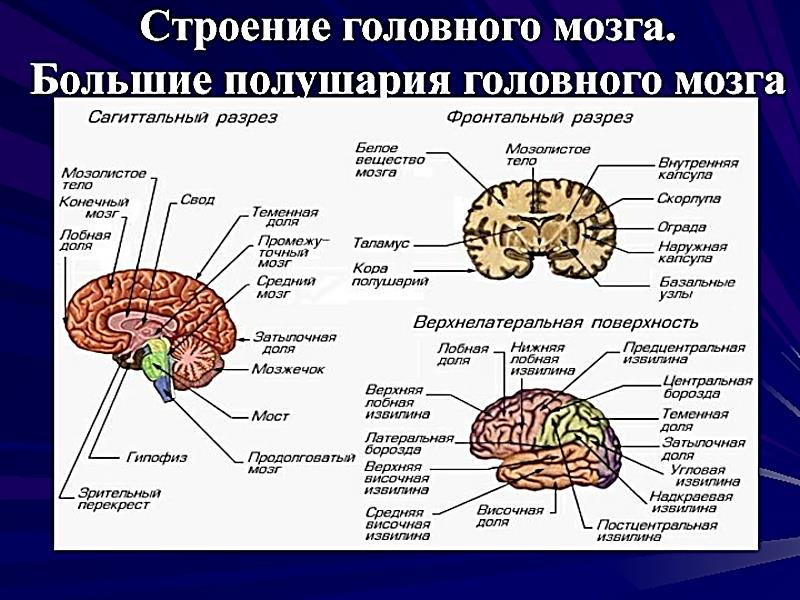 44. Локализация функций и коре головного мозга Совокупность нервных образований, обеспечивающих: трансформацию энергии раздражения в нервный импульс, проведение возбуждения, его анализ и синтез, обеспечивающих возникновение ощущений, называется анализатором. Следует уяснить, что каждый анализатор состоит из трех морфологических частей: 1) рецептор; 2) кондуктор; 3) корковый конец анализатора, где возбуждение воспринимается как ощущение. Под корковым концом анализатора следует понимать - участок коры головного мозга, в котором происходит высший анализ, синтез и интеграция функций. По И.П. Павлову: «Корковый конец анализатора – это ядро и рассеянные вокруг клеточные элементы». Данное определение объясняет частичное восстановление функции при повреждении ядра. Это позволяет говорить о динамической локализации функций в коре полушарий большого мозга. |