пр. Впервые нуклеиновые кислоты были обнаружены в 1868 г. Иоганом Фридрихом Мишером из ядер клеточного гноя
 Скачать 99.51 Kb. Скачать 99.51 Kb.
|

|
Впервые нуклеиновые кислоты были обнаружены в 1868 г. Иоганом Фридрихом Мишером из ядер клеточного гноя. Модель строения ДНК была предложена Уотсоном и Криком в 1953 г. Генетическая роль ДНК была доказана в 1928 г. в опытах Гриффита. Гриффит обнаружил явления трансформации у пневмококка. В 1944 году Эвери и его команда (Маклеод и Маккартни) установили химическую природу вещества. Молекула ДНК состоит из двух гигантских комплементарных и полимерных цепей соединенных между собой водородными связями по принципу комплементарности (т.е. аденин всегда соединяется с тимином, а гуанин с цитозином). В зависимости от вида основания выделяют следующие типы нуклеотидов: пуриновые (аденин, гуанин) и пиримидиновые (цитозин, тимин, урацил). Модель строения ДНК была предложена Уотсоном и Криком в 1953 г. Согласно этой модели, молекула ДНК состоит из двух антипараллельных цепочек соединенных между собой водородными связями по принципу комплементарности (т.е. аденин всегда соединяется с тимином, а гуанин с цитозином). ДНК обладает уникальными свойствами: способностью к самоудвоению (ауторепродукции или репликации) и к транскрипции. Кроме того, последовательность нуклеотидов в ДНК формирует генетический̆ код, посредством которого записывается информация о видовых и индивидуальных особенностях признаков и свойств организма. Перечисленные особенности химической̆ структуры и свойств ДНК обусловливают выполняемые ею функции. ДНК хранит, передает и реализует наследственную информацию. Ауторепликация- процесс самовоспроизведения молекул нуклеиновых кислот, сопровождающийся передачей по наследству (от клетки к клетке) точных копий генетической информации. Репликация происходит полуконсервативным способом, т.е. обе цепи ДНК разделяются, и на каждой̆ синтезируется комплементарная ей цепь. Репликация осуществляется под контролем ряда ферментов и протекает в несколько этапов. Она начинается с раскручивания двойной спирали ДНК с помощью фермента - геликазы – за счет разрыва слабых водородных связей между основаниями с образованием репликационной вилки. Так как ДНК прикреплена на ядерный матрикс, она не может свободно вращаться, поэтому выше точки начала репликации создается суперспирализация или супер напряжение. Топоизомераза и гираза устраняют суперспирализацию путем разрыва одной из цепей ДНК. С каждой из отдельной цепей ДНК связываются ССВ-белки, которые стабилизируют нити в таком состоянии вилки. Перед началом синтеза молекул ДНК, строится короткая последовательность ДНК(10-15) – РНК затравка или праймер при участии фермента РНК-полимераза. Синтез молекулы ДНК осуществляется ДНК-полимеразой III по принципу комплементарности 5’ 3'. ДНК-полимераза может строить только ведущую цепь (так как две цепи ДНК антипараллельны, т.е. направление 5’→3’ у них противоположно, то другие отстающие цепи синтезируют отдельные участки. Праймеры далее удаляются из молекулы ДНК и ДНК сливаются с ДНК-липазой. Конечный итог- 2 молекулы ДНК. Генетический код- это способ записи генетической информации о последовательности аминокислот в белке с помощью нуклеотидов в молекуле ДНК. Свойство генетического кода: 1) Код является триплетным, т.е. одну аминокислоту кодирует последовательность из трех нуклеотидов. Код универсален, одни и те же триплеты кодируют одни и те же амино- кислоты у всех живых организмов. Код является вырожденным, т.е. одна аминокислота может кодировать- ся несколькими триплетами (таблица 2). Код является неперекрывающимся, т.е. соседние триплеты не имеют общих оснований. Код является непрерывным, т.е. не содержит каких либо разделитель- ных знаков между отдельными триплетами. Код является линейным, триплеты расположены в линейном порядке. Код является коллинеарным, т.е. последовательность триплетов в моле- куле ДНК (или мРНК) полностью совпадает с последовательностью аминокислот в полипептиде. Ген – участок молекулы ДНК, кодирующий̆ последовательность аминокислот в полипептиде или последовательность нуклеотидов в молекуле т-РНК и р-РНК. Свойства гена: Ген – это участок молекулы ДНК, имеющий линейный характер. Ген дискретен, т.е. имеет начало и конец. Ген способен к прямым и обратным мутациям (за исключением деле- ций, у которых невозможны обратные мутации). Кроссинговер осуществляется как между генами так и внутри гена. Ген контролирует последовательность аминокислот в полипептиде, или последовательность нуклеотидов в тРНК и рРНК. Главная количественная особенность генетического материала эукариот – наличие избыточной̆ ДНК и по мере усложнения организмов доля кодирующей̆ ДНК уменьшается. Эта избыточность генома эукариот объясняется существованием нуклеотидных последовательностей̆ разной̆ степени повторяемости. Классы нуклеотидных последовательностей̆ в геноме эукариот
Различают следующие уровни структурно-функциональной организации наследственного материала: генный, хромосомный и геномный. Элементарной структурой ГЕННОГО уровня организации служит ген. На этом уровне изучается структура молекулы ДНК, биосинтез белка и др. Благодаря относительной независимости генов возможно дискретное (раздельное) и независимое наследование (III закон Менделя) и изменение (мутации) отдельных признаков. Гены клеток эукариот распределены по хромосомам, образуя ХРОМОСОМНЫЙ уровень организации наследственного материала. Этот уровень организации служит необходимым условием сцепления генов и перераспределения генов родителей у потомков при половом размножении (кроссинговер). Вся совокупность генов организма в функциональном отношении ведет себя как целое и образует единую систему, называемую ГЕНОМОМ. Один и тот же ген в разных генотипах может проявлять себя по-разному. Геномный уровень организации объясняет взаимодействие генов как в одной, так и в разных хромосомах. Хромосомы – это плотные интенсивно окрашивающиеся структуры, которые становятся видимыми (и изучаются) на стадии метафазы митоза. Каждая метафазная хромосома состоит из двух хроматид (рис.9). Хроматиды – сильно спирализованные идентичные молекулы ДНК, образовавшиеся в результате репликации. Хроматиды соединяются между собой в области первичной перетяжки или центромеры. Центромера делит хромосому на два плеча. В зависимости от положения центромеры различают метацентрические (срединное положение центромеры), субметацентрические (близкое к середине положение центромеры) и акроцентрические (близкое к одному из концевых участков положение центромеры) хромосомы. Иногда наблюдаются вторичные перетяжки, отделяющие спутники. Вторичные перетяжки участвуют в образовании ядрышка. 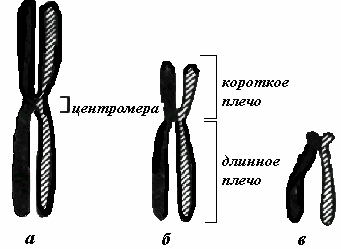 По своему химическому составу хромосомы являются нуклеопротеидами, т.е. в основном состоят из белков и нуклеиновых кислот (ДНК). При этом белки составляют примерно 65% массы. Все хромосомные белки разделяются на две группы: гистоны и негистоновые белки. Гистоны представлены 5 фракциями: Н1, Н2А, Н2В, Н3, Н4. Взаимодействуя с ДНК гистоны выполняют структурную функцию, обеспечивая пространственную организацию ДНК в хромосомах. Негистоновые белки очень разнообразны, как и выполняемые ими функции. Так, некоторые из негистоновых белков участвуют в образовании наднуклеосомных уровней организации ДНК. Кроме того, среди них выделяют ферменты, обеспечивающие процессы синтеза и процессинга РНК, репликации и репарации ДНК. И третью группу негистоновых белков составляют регуляторные белки, контролирующие активность вышеуказанных ферментов. Помимо белков в составе хромосом обнаруживаются также липиды, полисахариды, ионы металлов, РНК. Уровни упаковки молекулы ДНК: Нуклеосомный- Молекула ДНК связана с гистоновыми белками, эти белки собираются в виде глобул, каждая глобула содержит по две молекулы следующих гистоновых белков (2Н2а, 2Н2b, 2H3, 2H4). Она делает вокруг одного белкового комплекса 2,5 оборота вокруг одного белкового комплекса, а затем вокруг другого такого же. Этот уровень позволяет уменьшить размер ДНК в 6-7 раз. Супернуклеосомный- нуклеосома спирально закручивается. На этом уровне формируется соленоидная структура. Хроматидный- соленоиды образуют петли, образуется эухроматин и ДНК уменьшается в 25 раз. Хромосомный- компактизации является образование метафазной хро- мосомы. Таким образом, сокращение идет в 10 000 раз. 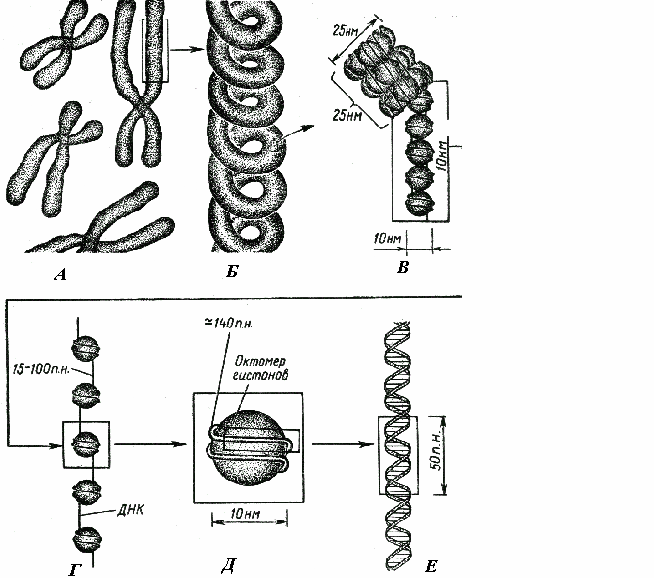 Морфологическими элементами метафазной хромосомы являются: - 2 хроматиды; - центромера; - теломеры; - вторичная перетяжка; - спутник (сателлит); - ломкие (фрагильные) участки. Хроматида представлена одной линейной молекулой ДНК, ассоциированной с гистоновыми и негистоновыми белками и максимально конденсированной. Метафазная хромосома состоит из двух сестринских хроматид, являющихся результатом репликации ДНК в фазе S и, таким образом, генетически идентичных. Хроматиды одной хромосомы соединены в области центромеры и остаются в таком состоянии до анафазы. Эухроматин – функционально активные, практически деконденсированные участки хромосом. Гетерохроматин – сильно спирализованные и функционально неактивные участки хромосом. Гетерохроматин локализован преимущественно на периферии ядра и прилежит к ядерной оболочке. Конститутивный гетерохроматин содержится в околоцентромерных и теломерных участках всех хромосом, а также на протяжении некоторых внутренних фрагментов отдельных хромосом. Он образован только нетранскрибируемой ДНК. Вероятно, его роль заключается в поддержании общей структуры ядра, прикреплении хроматина к ядерной оболочке, взаимном узнавании гомологичных хромосом в мейозе, разделении соседних структурных генов, участии в процессах регуляции их активности. Систематизированный набор хромосом единичной соматической клетки, представленный в графическом виде называют кариотипом. Нормальный кариотип человека включает 46 хромосом, из них 22 пары аутосом и 1 пара половых хромосом. В 1960 г. была разработана первая международная классификация хромосом человека (Денверская). В соответствии с этой классификацией все хромосомы человека в зависимости от размера хромосомы и положения центромеры были разделены на 7 групп (A,B,C,D,E,F,G). После открытия методов дифференциальной окраски хромосом, позволявших установить положение хромосомы внутри группы, в 1971 г. принята унифицированная система идентификации хромосом (Парижская классификация) (рис.10). |