доп материал психология 22-23. Введение в психологию
 Скачать 1.45 Mb. Скачать 1.45 Mb.
|

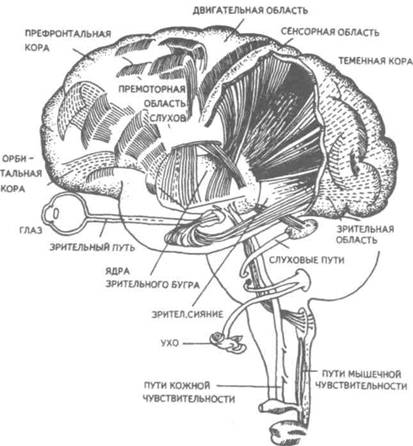 Рис. 14. Корковые зоны систем анализаторов (по Д.Пейпецу) На рис. 14 показаны корковые зоны основных систем анализаторов человека, работа которых совместно с органами чувств и рецепторами порождает множество ощущений разных модальностей и различного качества. На следующих далее рисунках они представлены в отдельности для зрения (рис. 15), слуха (рис. 16), обоняния (рис. 17), вкуса (рис. 18), осязания (рис. 19) и равновесия (рис. 20). В целом физиологический механизм формирования ощущений, включая неосознаваемые, с учетом роли и действия ретикулярной формации, видится следующим образом. На многочисленные интеро- и экстерорецепторы ежесекундно воздействует масса разнообразных стимулов, причем лишь незначитель- 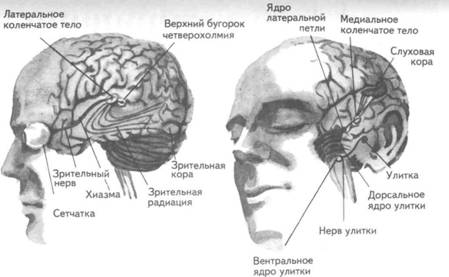 Рис. 15. Зрительная система. Показаны связи, идущие от первичных рецепторов сетчатки через передаточные ядра таламуса и гипоталамуса к первичной зрительной зоне коры Рис. 16. Слуховая система. Показаны связи, идущие от первичных рецепторов улитки через таламус к первичной слуховой зоне коры 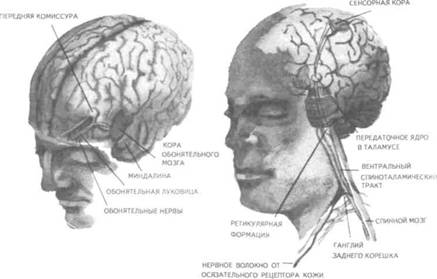 Рис. 17. Обонятельная система. Показаны связи, идущие от рецепторов слизистой носа через обонятельные луковицы и базальные ядра переднего мозга к конечным пунктам в обонятельной коре Рис. 18. Ощущения с поверхности тела. Представлены связи, идущие от кожных рецепторов через вставочные нейроны спинного мозга и таламуса к первичной сенсорной зоне коры ная часть из них вызывает реакции в рецепторах. Попадая на специализированные рецепторы, они возбуждают их; рецепторы преобразуют энергию воздействующих стимулов в нервные импульсы, которые в закодированном виде несут в себе информацию о жизненно важных параметрах стимула. Далее эти импульсы попадают в ц.н.с. и на разных ее уровнях — спинного, промежуточного, среднего и переднего мозга — многократно перерабатываются. В к.г.м. поступает уже переработанная, отфильтрованная и отсеянная информация, где, достигая проекционных зон коры, она порождает ощущения соответствующей модальности. С помощью ассоциативных волокон, связывающих между собой отдельные части к.г.м., эта информация, вначале представленная на уровне отдельных ощущений, интегрируется, вероятно, в образы. Образ, складывающийся в результате восприятия как психофизиологического процесса, предполагает согласованную, ко- 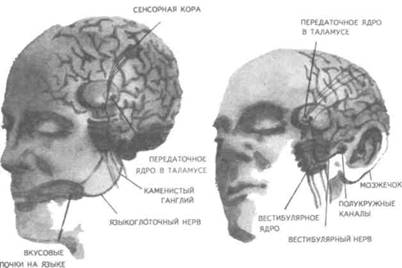 Рис. 19. Вкусовая система. Изображены связи, идущие от рецепторов языка через первоначальные мишени варолиева моста к мишеням следующего порядка в коре больших полушарий Рис. 20. Чувство равновесия. Показаны связи, идущие от первичных рецепторов преддверия внутреннего уха (вестибулярного аппарата) к ядрам ствола мозга и таламуса. Эта информация, по-видимому, не имеет путей для передачи в кору большого мозга ординированную деятельность сразу нескольких анализаторов. В зависимости от того, какой из них работает активнее, перерабатывает больше информации, получает наиболее существенные признаки о свойствах воспринимаемого предмета, различают и виды восприятия. Соответственно выделяют зрительное, слуховое, осязательное восприятие, при которых доминирует один из следующих анализаторов: зрительный, слуховой, тактильный (кожный), мышечный. Зрительное восприятие имеет наиболее важное значение в жизни человека, а его орган — глаз и связанные с ним отделы мозга представляются наиболее сложно устроенными из всех анализаторов. Приведем некоторые данные, касающиеся ана-томо-физиологического устройства зрительной системы. Внутренняя оболочка глазного яблока — сетчатка. В ней находятся особые световоспринимающие элементы, называемые соответственно их форме палочками и колбочками. Центральная часть сетчатки, называемая фовеа, является ее наиболее чувствительным местом. В ней сосредоточены только колбочки (около 50 000), сконцентрированные на площади размером меньше чем 1 см2. В остальной части сетчатки имеются как палочки, так и колбочки, причем от центра к периферии их концентрация постепенно уменьшается. С головным мозгом палочки и колбочки соединены идущими от них нервами, которые имеют переключения через еще два слоя расположенных в сетчатке нервных клеток. Кроме того, через специальные горизонтальные соединительные клетки, также имеющиеся в сетчатке, ряд палочек и колбочек непосредственно соединяется друг с другом. Такая структура обеспечивает многоуровневую вертикально-горизонтальную передачу, переработку и интеграцию стимулов, воспринимаемых светочувствительными элементами: палочками и колбочками. Чем ближе к центру сетчатки, тем меньше палочек и колбочек горизонтально соединено друг с другом; чем дальше от центра, тем крупнее системы взаимно объединенных друг с другом палочек и колбочек. Благодаря такому анатомо-физиологическому устройству части зрительного анализатора воспринимающая система получает сразу два полезных свойства. Во-первых, соединение светочувствительных элементов друг с другом в системы, охватывающие значительные площади и пространства воспринимаемого мира, позволяет улавливать и усиливать (путем их суммирования) сравнительно небольшие воздействия света, ощущать их и обращать на них внимание. Во-вторых, большое количество светочувствительных элементов, сконцентрированных на небольшой площади ближе к центру сетчатки и имеющих отдельные независимые выходы в мозг, позволяет при необходимости лучше различать тонкие детали изображений, выделять и внимательно рассматривать их. Интеграция зрительной информации по вертикали обеспечивается также двумя свойствами анатомо-физиологического устройства зрительного анализатора. Первое из них — наличие многих уровней переключения поступающей с периферии информации, прежде чем она попадет в кору головного мозга. Это позволяет многократно анализировать одну и ту же информацию с разных сторон, а также отбирать из нее наиболее полезные сведения, отсеивая ненужные и второстепенные. Другое свойство связано с наличием рецептивного поля. Рецептивным полем нейрона коры головного мозга, например, называется система периферических рецепторов, воздействие на которые вызывает возбуждение одного и того же нейрона 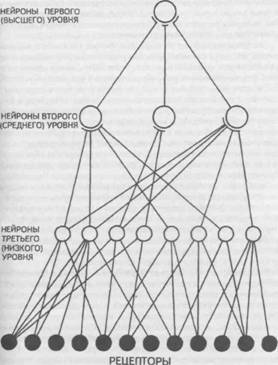 Рис. 21. Схема рецептивных полей разного уровня коры головного мозга (или одного и того же нейрона более высокого уровня в нервной системе). На рис. 21 изображена схема рецептивного поля нейрона коры головного мозга. Из нее видно, что, прежде чем нервное возбуждение от рецептора попадает в кору, оно переключается на двух промежуточных уровнях. По отношению к нейронам каждого из выделенных уровней есть смысл говорить о собственных рецептивных полях, включающих все нейроны нижележащего уровня, которые имеют выход (синаптический контакт) на данный нейрон. На приведенной схеме показано, что рецептивные поля различны по тому, какое количество рецепторов или нейронов из нижележащих уровней они включают. По современным данным формирование образа обеспечивается слаженной работой множества рецептивных полей, которые в свою очередь объединены в так называемые клеточные ансамбли. «Наименьшими единицами восприятия должны быть результаты возбуждения клеточных ансамблей, а не специфических рецепторных нейронов»1. Каждый такой клеточный ансамбль включает множество взаимосвязанных рецептивных полей разного уровня и должен соответствовать простому элементу сенсорного образа: углу или наклону линии в зрении, фонеме или различимому звуку в речи, определенной форме давления в осязании. Уже найдены нейроны, которые приходят в состояние возбуждения при стимулировании всего поля сетчаточных рецепторов определенной конфигурацией, например линией или краем специфической ориентации и наклона. Они не возбуждаются, если стимулируется не весь «клеточный ансамбль», а отдельные палочки и колбочки отдельными световыми точками в той же области сетчатки. Эти клетки, чувствительные к краям и линиям, расположены в сенсорной проекционной зоне зрения. В заключение обсуждения нервных механизмов зрения отметим, что вклад в их работу вносит уже отмеченная нами функциональная асимметрия мозга. Оба полушария, левое и правое, играют различную роль в восприятии и формировании образа. Для правого полушария характерны высокая скорость работы по опознанию, его точность и четкость. Такой способ опознания предметов можно определить как интегрально-синтетический, целостный по преимуществу, структурно-смысловой2. Правое полушарие, вероятно, производит сличение образа с некоторым имеющимся в памяти эталоном на основе вьщеления в воспринимаемом объекте некоторых информативных признаков. С помощью же левого полушария осуществляется в основном аналитический подход к формированию образа, связанный с последовательным перебором его элементов по определенной программе. Но левое полушарие, работая изолированно, по-видимому, не в состоянии интегрировать воспринятые и выделенные элементы в целостный образ. С его помощью производится классификация явлений и отнесение их к определенной категории через обозначение словом. Таким образом, в восприятии с разными функциями одновременно принимают участие оба полушария головного мозга. Рассмотрим теперь физиологические механизмы внимания. Как общее состояние сосредоточенности оно связано с повышением возбудимости к.г.м. в целом или ее отдельных участков. Это в свою очередь соотносится с активностью отдельных частей ретикулярной формации. Те ее отделы, которые своей деятельностью порождают общий эффект возбуждения, входят в структуры, связанные с ориентировочным рефлексом, автоматически возникающим при любых неожиданных и заметных изменениях стимулов, воздействующих на организм. В свою очередь те отделы ретикулярной формации, которые вызывают специфический эффект возбуждения, функционируют, по-видимому, в рамках анатомо-физиологической системы доминанты. С ней же скорее всего соотносим в своем действии и избирательный механизм регуляции внимания через актуализацию потребностей, а также механизм волевого управления вниманием через кортикально-подкорковые связи. Много споров в науке было вокруг вопроса о физиологических основах внимания. Органическая интерпретация процессов, внимания привлекала исследователей на протяжении всего времени его изучения. Т.Рибо одним из первых пытался представить физиологическую схему произвольного внимания, связанного с волевым усилием, возникающим в процессе припоминания (рис. 22). Рассмотрим его концепцию. На рис. 22 М — центр, где хранятся воспоминания о некогда совершенных действиях (предполагаемый центр долговременной памяти на движения); Siи S2 — центры, где сохраняются сенсорные образы (центры сенсорной памяти). Те и другие локализованы в коре головного мозга, представленной на рисунке дугообразной заштрихованной областью. Эти центры между собой соединены волокнами ассоциативного типа; R — центр, расположенный ниже коры и связанный с регуляцией движений; С,, С2, С3 — совокупность сенсорных центров разного уровня, передающих коре больших полушарий информацию о состоянии мышечной системы М («мышечное чувство», по Т.Ри-бо); Т — область таламуса. Допустим, что в кортикальных центрах М, Sxи S2существует некоторый уровень возбуждения, т.е. они находятся в состоянии, связанном с вниманием, субъективно представляя собой ряд воспоминаний сенсорного и двигательного характера о предмете А. Моторное возбуждение из центра Мдалее распространяется вниз до центра R и через него производит сокращение мышц Р. Это сокращение по нервным центростремительным волокнам через ряд промежуточных сенсорных центров С,, С2и С3 вновь попадает в центр М коры. В результате «моторное воспоминание» будет усилено также и иннерва-ционным импульсом, явившимся в Т через рефлекторное возбуждение, восходящее от R. Произойдет двойное дополнительное усиление активности в центре М, которое далее по ассоциативным волокнам распространяется в коре больших полушарий до сенсорных центров 5, и S2и возбудит их дополнительно. В результате весь комплекс воспоминаний, моторных и сенсорных, связанных с предметом А, усилится в сознании, т.е. интенсивность внимания к нему возрастет. Т.Рибо предложил так называемую моторную теорию внимания, согласно которой основную роль в процессах внимания играют движения. Именно благодаря их избирательной и целенаправленной активизации происходят концентрация и усиление внимания на предмете, а также поддержание внимания на данном предмете в течение определенного времени. Пользуясь приведенной выше схемой, Т.Рибо следующим образом объясняет участие движения в описанном процессе. Внимание к предмету А начинается с слабого припоминания о нем. С помощью движения М мы усиливаем это воспоминание. Усиление воспоминания по ассоциации вызывает в свою очередь первоначально слабое припоминание о предмете В, которое мы далее усиливаем движением М, и т.д. «Процесс волевого внимания протекает именно указанным путем, т.е. каждый член этого ряда на момент усиливается и в этот же момент вызывает следующий член ассоциации. Обыкновеннейшим рядом движений служит при этом то, что называется внутренней (про себя) речью; каждый член ряда фиксируется нами произнесением про себя его имени или соответствующего суждения, и этим моментальным усилением его мы пользуемся для перехода к следующему члену ряда»1. 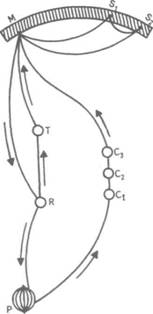 Рис. 22. Схема физиологического процесса, объясняющего усиление произвольного внимания при припоминании (по Т.Рибо) Аналогичную мысль о физиологическом механизме внимания высказывал АА.Ухтомский. Он считал, что физиологической основой внимания является доминантный очаг возбуждения, усиливающийся под воздействием посторонних раздражителей и вызывающий торможение соседних областей. В последние несколько десятилетий в связи с развитием генетики и молекулярной физиологии, а также кибернетики привлекли к себе внимание исследования биологических основ и физиологических механизмов памяти. Часть этих исследований была проведена на нейронном уровне, т.е. на уровне изучения работы отдельных нервных клеток и их ансамблей в процессе запоминания (научения). Было показано, что следы памяти обнаруживаются в изменениях, которые в процессе научения происходят в нервных клетках отдельных внутренних структур головного мозга. Это выражается, в частности, в повышении пластичности (откликаемости на стимулы) нейронов гиппокампа, ретикулярной формации и двигательной коры в процессе научения. Сложились гипотезы о роли глиальных элементов, молекул РНК и ДНК в процессах памяти. Некоторые ученые полагают, что глия — клетки в головном и спинном мозге, заполняющие пространства между нейронами и кровеносными сосудами, — связана с работой долговременной памяти. Предполагается также, что память соотносится с изменениями в структуре молекул рибонуклеиновой кислоты — РНК, а также с содержанием РНК в тех или иных образованиях мозга. В коре головного мозга следы памяти или научения обнаруживаются в виде изменений в клетках ц.н.с, наиболее типичными из которых являются увеличение диаметра афферентных окончаний, увеличение числа и длины терминалей аксона, изменение формы клеток к.г.м., увеличение толщины волокон в клетках к.г.м. I и II слоев. Потери памяти чаще всего можно наблюдать при поражениях лобных и височных долей мозга, поясной извилины, а также ряда подкорковых структур: мамиллярных тел, передних отделов таламуса и гипоталамуса, амигдолярного комплекса и особенно гиппокампа (рис. 23). Его поражение ведет к нарушению процесса выявления следов прошлого опыта, но не к утрате самих следов. Немаловажна в процессах памяти, вероятно, и роль ретикулярной формации, связанных с ней активизирующих структур мозга. Д.О.Хебб предложил гипотезы о физиологических механизмах кратковременной и долговременной памяти. Таким механизмом для кратковременной памяти, по его мнению, является реверберация (вращение) электрической активности в замкнутых цепях нейронов, а долговременная память связана с морфофункцио-нальными изменениями устойчивого характера, происходящими в синапсах, увеличивающими или уменьшающими их проводимость. Из кратковременной в долговременную память информация переходит в результате процесса «консолидации», который развивается при многократном прохождении нервных импульсов через одни и те же синапсы. Этот процесс длителен и требует не менее нескольких десятков секунд реверберации. Консолидация представляет собой облегчение и упрочение синаптической проводимости. В итоге описанных процессов образуются нервные модели стимулов или клеточные ансамбли, причем любое возбуждение, затрагивающее соответствующие нейронные структуры, может привести в состояние активности весь ансамбль. Предполагается, что психологически такая активизация может происходить под влиянием различных стимулов, ощущений, образов, эмоций и т.п. По предположению Хебба, нейронные цепи и структуры, о которых идет речь, могут образовываться с включением во взаимодействие коры, таламуса и гипоталамуса. Нейроны, которые в процессе научения обнаруживают пластические изменения, кроме названных мозговых структур, имеются в гиппокампе, миндалине, хвостатом ядре, ретикулярной формации и двигательной коре. 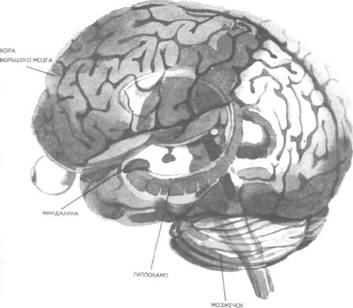 Рис. 23. Структуры мозга, вероятнее всего связанные с памятью Г.Хиденом была предложена гипотеза о роли РНК в процессах долговременной памяти, допускающая изменение в результате научения последовательности оснований в молекуле РНК. Однако носителем самой прочной, самой глубокой памяти человека является, по-видимому, ДНК, которую к тому же считают входящей в механизм наследственности. С двигательной памятью человека, и особенно со сложными формами автоматизированных движений, осуществляемых на подсознательном уровне, связана работа мозжечка. Установлено, что при нарушениях работы мозжечка человек вынужден сознательно контролировать каждый элемент сравнительно простых движений, которые раньше осуществлял автоматически, не задумываясь. Например, для того чтобы взять в руки и откусить яблоко, ему приходится сначала отдельно осуществить и полностью завершить акт хватания, сделав после этого остановку, затем поднять таким же образом руку на уровень рта и только после этого поднести яблоко ко рту. С мозжечком, вероятно, связана и память на множество условных рефлексов. Функция гиппокампа в процессах памяти до сих пор точно не известна, хотя есть данные, свидетельствующие о том, что он как-то связан с кратковременной, памятью, а также, вероятно, с оперативной памятью. Люди с мозговыми нарушениями, локализованными в области гиппокампа, не могут хранить в памяти то, о чем узнали совсем недавно, или забывают о том, что уже начали или намеревались сделать после того, как дело уже начато. Например, им нелегко вспомнить лицо и имя недавно виденного человека, с которым их познакомили, однако память на давние прошлые события у них обычно бывает сохранной. Можно также предположить, что работа гиппокампа как-то связана с другим процессом памяти — узнаванием. Есть также сведения о том, что активность гиппокампа соотносится с переводом информации из кратковременной в долговременную память. В одном описанном в литературе случае, когда в результате неизбежного хирургического вмешательства у человека был удален гиппокамп, выяснилось, что этот человек сохраняет в памяти лишь то, что с ним происходит сейчас. Воспоминания о недавнем прошлом стерлись, а события настоящего надолго также не запоминались. Кроме гиппокампа, в формировании и организации следов памяти участвует, по-видимому, медиальная височная область мозга. Л.Сквайр высказал предположение о том., что височная область мозга не является хранилищем информации, находящейся в долговременной памяти, но участвует в реорганизации нервных структур и в установлении связи с местами хранения такой информации, прежде всего с корой головного мозга. Такая реорганизация может быть связана с физической перестройкой нервных процессов. Таламическая область, как предполагают, нужна для первоначального кодирования некоторых видов информации, получаемой через органы чувств. Что касается коры головного мозга, то есть данные о ее связи с долговременной памятью. Кроме ощущений, внимания, восприятия и памяти, большой интерес к себе традиционно вызывают исследования физиологических механизмов потребностных состояний. Этот интерес связан также с тем, что в мотивационных процессах участвует не только ц.н.с, но организм в целом. Что же касается мозговых структур, то, вероятно, нельзя назвать ни одной из них, которая не имела бы прямого или косвенного отношения к удовлетворению потребностей. Но более всего с ними, по-видимому, связаны таламус, через который проходят почти все нервные пути, идущие в к.г.м. и обратно, подкорка, древняя, старая и новая кора. Считается, что физиологически потребность представляет собой состояние отклонения от нормы во внутренних тканях и органах, которое субъективно выражается в форме ощущений и эмоций. Выделить и назвать какую-либо одну или несколько мозговых структур как общую основу мотивации и эмоций практически невозможно. Однако в психологии различают элементарные эмоции и высшие чувства, причем первые считаются в основном врожденными, а вторые — приобретенными в результате научения. Анатомо-фи-зиологической основой низших или простейших эмоций являются лимбические структуры ц.н.с, а также процессы, происходящие в таламусе и гипоталамусе (рис. 24, 25). Нервным субстратом высших чувств человека скорее всего является к.г.м. Широкую известность в психологии получила теория, объясняющая функционирование и происхождение эмоций их тесной связью с органическими процессами. Такую теорию почти в одно и то же время предложили американский психолог У.Джемс и датский ученый КЛанге. В историю науки она вошла под двойным названием как теория Джемса—Ланге. Согласно этой теории первопричинами возникновения эмоциональных состояний являются изменения физиологического характера, происходящие в организме. Возникнув под влиянием внешних или внутренних стимулов, они затем отражаются в голове человека через систему обратных нервных связей и порождают ощущение определенного эмоционального тона. Сначала, согласно теории Джемса—Ланге, должны произойти соответствующие органические изменения в ответ на воздействия стимулов, и только затем как их субъективно отраженное следствие возникает эмоция. Разница, которая обнаружилась во взглядах У.Джемса и КЛанге, была небольшой. Джемс полагал, что телесные изменения непосредственно следуют за восприятием возбуждающих стимулов, а эмоция есть не что иное, как наше ощущение уже произошедших перемен. КЛанге же считал, что сенсорные сти мулы возникают непосредственно в рецепторах кровеносных сосудов, что эти сосуды в первую очередь реагируют на внешние воздействия, а эмоции появляются затем как отражение произошедших в них изменений. Суть концепции Джемса—Ланге с последовательностью постулируемых ею событий, приводящих к возникновению эмоций, схематически представлена на рис. 26. Концепция происхождения эмоций, предложенная Джемсом и Ланге, наряду с одобрением встретила ряд возражений. Наиболее серьезные замечания в ее адрес были высказаны У.Кен-ноном, который одним из первых обратил внимание на то обстоятельство, что телесные реакции, возникающие при различных эмоциях, весьма похожи друг на друга и как таковые недостаточны для того, чтобы вполне удовлетворительно объяснить качественное многообразие существующих у человека эмоций. Кроме того, внутренние органические структуры, в частности кровеносные сосуды, с изменениями которых КЛанге соотносил возникновение эмоциональных переживаний, инертны и малочувствительны, очень медленно приходят в состояние возбуждения. Что же касается эмоций, то они появляются как субъективные состояния почти мгновенно при возникновении эмоциогенной ситуации, по крайней мере гораздо быстрее, чем на нее своими изменениями внутреннего порядка реагирует организм. 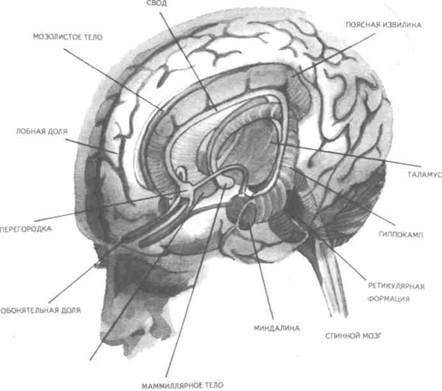 Рис. 24. Важнейшие части мозга, образующие лимбическую систему. Можно видеть, что они располагаются вдоль краев больших полушарий, как бы «окаймляют» их 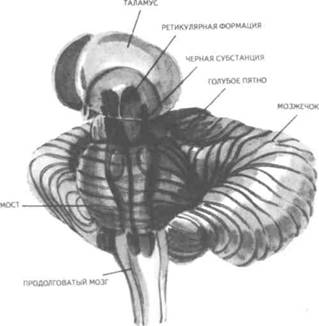 Рис. 25. Структуры ствола мозга, играющие роль в эмоциях. Дофаминовые волокна, идущие от черной субстанции, и норадреналиновые волокна, идущие от голубого пятна, иннервируют весь передний мозг. Обе эти группы нейронов, а также некоторые другие представляют собой части ретикулярной активирующей системы. Самым сильным возражением У.Кеннона против теории Джемса—Ланге явилось следующее: искусственно вызываемые у человека специальными фармакологическими средствами органические изменения, те самые, существованию которых Джемс и Ланге приписывали возникновение эмоций, далеко не всегда сопровождаются эмоциональными переживаниями. Но даже и в том случае, когда в условиях искусственной органической стимуляции подобные эмоциям переживания все же появляются, они субъективно воспринимаются человеком совсем не так, как настоящие эмоции. 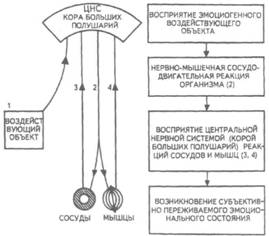 Рис. 26. Концепция Джемса—Ланге В результате У.Кеннон предложил альтернативную теорию происхождения эмоций, представленную в ее основных положениях на рис. 27.  Рис. 27. Концепция возникновения эмоций Кеннона—Барда У.Кеннон считал, что эмоциональные переживания и соответствующие им органические изменения порождаются одновременно и возникают из единого источника. Таким источником — эмоциогенным центром, по мнению У.Кеннона, — является таламус, играющий важную роль в регуляции основных органических процессов. Возникновение эмоции есть результат одновременного возбуждения через таламус симпатической нервной системы и коры головного мозга. Высказанные У.Кенноном положения были дальше развиты П.Бардом. Он показал, что в действительности как телесные изменения, так и связанные с ними эмоциональные переживат ния возникают почти в одно и то же время. Собственно с эмоциями из всех структур головного мозга более всего связан не сам таламус, а гипоталамус и центральная часть лимбической системы. В экспериментах, выполненных на животных, удалось доказать, что электрическими воздействиями на эти структуры в определенной степени можно управлять эмоциональными состояниями, такими, например, как страх и гнев. В результате этих доработок концепция У.Кеннона получила новое, современное название теории Кеннона—Барда. 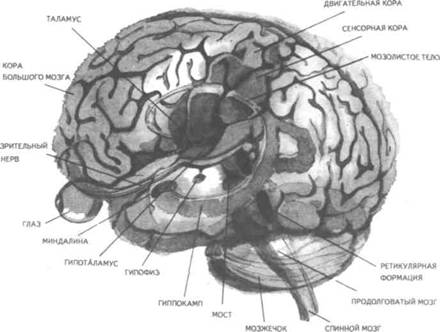 Рис. 28. Нерасчлененный мозг. На рисунке представлены основные структуры, принимающие участие в работе познавательных процессов и в потребительско-эмоциональной регуляции поведения, включая лимбическую систему На рис. 28, заключающем данный параграф учебника, в интегрированном виде представлены основные структуры головного мозга, принимающие участие в познавательных и эмоци-онально-мотивационных процессах, рассмотренных выше. ГЕНЕТИЧЕСКИЕ КОРНИ ПСИХОЛОГИИ И ПОВЕДЕНИЯ Проблема генетических источников психологии и поведения человека является одной из важнейших в психологической и педагогической науках, поскольку от ее правильного решения зависит принципиальное решение вопроса о возможностях обучения и воспитания детей, человека вообще. По современным данным напрямую воздействовать через обучение и воспитание на генетический аппарат практически невозможно, и, следовательно, то, что задано генетически, перевоспитанию не подлежит. С другой стороны, обучение и воспитание сами по себе обладают огромными возможностями в плане психического развития индивида, даже если они не затрагивают собственно генотипа и не воздействуют на органические процессы. В целом поставленная проблема в психологии возникла давно и сформулирована как проблема генотипической и средовой обусловленности психики и поведения человека. Связанный с ней вопрос — о биологической и социальной детерминации психических явлений — касается зависимости функционирования и развития психических явлений от организма, не обязательно только от тех его свойств, которые обусловлены генотипически. Это могут быть органические особенности, которые сформировались у человека в процессе жизни в результате созревания, обучения и воспитания вместе взятых. Наконец, само по себе средовое не обязательно сводится только к социальному: ведь окружающая человека среда не ограничена обществом, это может быть и природная среда. Социальные воздействия на человека связываются, как правило, с миром людей, с их материальной и духовной культурой, в то время как среда, понимаемая в широком смысле слова, включает все остальное, что с рождения и в течение всей жизни окружает человека. На рис. 29 показано соотношение двух обсуждаемых проблем и четырех связанных с ними понятий. Проблемы здесь представлены взаимно перпендикулярными пересекающимися линиями, определяемыми на своих концах словами: «генотип», «среда», «биологическое», «социальное». Фактически на пересечении этих линий и понятий возникает уже не две, а четыре частные проблемы: I. Проблема генотипической и социальной обусловленности психики и поведения. II. Проблема социальной и средовой обусловленности психики и поведения. III. Проблема генотипической и биологической обусловленности психики и поведения. IV. Проблема средовой и биологической обусловленности психики и поведения. В Действительности генотипическое и биологическое начала в человеке часто объединяют, называя их одним словом — «биологическое». Так же точно иногда соединяют средовое и социальное в понятии «социальное». Таким образом, «биологиче ское» оказывается интегрирующим в себе и наследственно закрепленные физические (конституциональные) и заданные с рождения физиологические (процессуальные) свойства человеческого организма. Биологическое, понимаемое в этом, расширительном, значении, может быть врожденным или приобретенным в пренатальный период развития организма ребенка или возникнуть после рождения в результате болезни. Именно так мы и будем далее рассматривать интересующую нас проблему, особо выделяя при ее обсуждении вопрос о генотипической и средовой обусловленности психики человека. Соответствующая область исследований, занимающая промежуточное положение между психологией и генетикой, в последние годы становится особенно популярной и получила двойное наименование: в США — «социобиология», в Европе — «этология человека». 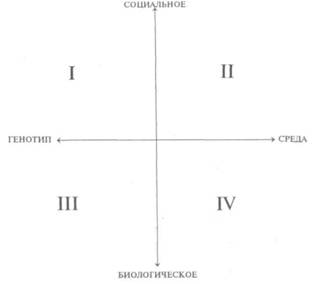 Рис. 29. Схематизированное представление четырех проблем, возникающих на стыке генотипических, средовых, биологических и социальных влияний на человека, его психологию, поведение Большинство концепций, представленных в социобиологии, и выводов, положенных в их основу, базируется на результатах сравнения человека с другими существами или на выявленном сходстве определенных поведенческих реакций, видов поведения различных популяций людей. Речь идет о таких научных данных, которые не зависят от имеющихся в разных группах людей специфических культурно-исторических традиций. Отношение ученых к социобиологии в настоящее время неоднозначное: есть и ее сторонники, и ее противники. Самый существенный аргумент, выдвигаемый против социобиологии как науки, — это отсутствие прямых и достаточно убедительных доказательств непосредственной генетической детерминированности большинства видов и форм человеческого поведения. Неоправданным, в частности, представляется оппонентам предположение о том, что человек в своем поведении не зависит от среды, а работа его ц.н.с. не контролируется генетически. Обе точки зрения, выраженные в этом предложении, в настоящее время почти не имеют сторонников и ни одну из них нельзя признать доказательной. Остается только компромиссный вариант, но и он не решает проблемы. Этот вариант выражен в решении, которое сформулировано в утверждении: «частично от генотипа, частично от среды». Данный вариант в каждом конкретном случае требует определения, что же на самом деле зависит от среды и что от генотипа. Поиски практического решения данного вопроса начались с появления работы английского ученого Ф.Гальтона «Наследование таланта и характера» (1865), где впервые был применен метод близнецов с соответствующими статистическими расчетами, с помощью которых определялись различия между группами близнецов в изучаемых признаках. В результате этого исследования возникла мысль о вероятной генетической обусловленности человеческого интеллекта, а также некоторых психологических и поведенческих признаков, свидетельствующих об умственной отсталости человека. Исследования хромосом человека (их число равно 48), развернувшиеся в конце 50-х—начале 60-х гг. текущего века, положили начало второму важному этапу разработки проблемы генетической обусловленности психики и поведения. Область науки, занятая этой проблемой в настоящее время, называется генетикой поведения. Она касается определения наследственных факторов, влияющих на поведение людей, их познавательные процессы и способности. Генетика развития, дополняющая данную научную область, исследует генетические механизмы нормального и аномального психологического развития человека. В целом, однако, это направление в генетике пока что разработано слабо. Следует признать, что практически все в организме человека: и его развитие, и анатомо-физиологическое строение, и основные жизненные функции — в конечном счете находится под определенным контролем со стороны генных структур, Мозг, как и любой другой орган, обнаруживает генетическую детерминированность и изменчивость, поэтому логично предположить, что должны существовать генетические факторы, которые по крайней мере в ранние годы определяют психологию и поведение человека. Однако на практике действие генотипа и влияние среды отделить друг от друга очень трудно. Много научно полезной информации о генетической детерминации поведения и психологии человека дал близнецовый метод, предполагающий одновременное сравнительное исследование монозиготных (МЗ) и дизиготных (ДЗ) близнецов, первые из которых обладают идентичной, а вторые — различной наследственностью. Поскольку те и другие (МЗ и ДЗ), взятые попарно, живут и развиваются примерно в одинаковых условиях среды, то можно предположить, что межиндивидуальное сходство и различие внутри пар близнецов свидетельствует соответственно о значимости или генетического (сходство), или средо-вого (различие) фактора. Совокупность прирожденных признаков, определяемых исключительно генетически, называют генотипом, а признаки, характерные для уже ставшего взрослым организма, прошедшего длительный и сложный путь развития в определенных условиях среды, — фенотипом. Считается, что фенотип есть результат совместного влияния на организм и генотипа, и среды, в которой организм растет и развивается. ДЗ-близнецы в отличие от МЗ-близнецов имеют примерно половину общих генов, т.е. такое их распределение, которое близко к распределению генов у детей, родившихся в разное время в одной и той же семье, в то время как у МЗ-близнецов все гены одинаковые. Фенотипические различия между МЗ-близнецами обычно объясняются влиянием среды, т.е. всем, что непосредственно не связано с генами. ДЗ-близнецов в этих случаях традиционно используют для сравнения в качестве контрольной группы. В большинстве исследований, проводимых с использованием близнецового метода, ставится и решается следующий вопрос: отличаются ли близнецы от неблизнецов по изучаемому признаку? Сопоставление внутрипарного психологического и поведенческого сходства у МЗ- и ДЗ-близнецов должно показать относительную роль генотипа и среды в порождении и закреплении межиндивидуальных вариаций данного признака. В случае, если он в основном формируется под влиянием среды, внутрипарное сходство МЗ- и ДЗ-близнецов должно быть примерно одинаковым. Если же признак больше контролируется генотипическими факторами, то сходство у МЗ-близнецов должно быть значительно выше, чем у ДЗ-близнецов. Рассмотрим факты, подтверждающие или, напротив, отрицающие генотипическую обусловленность психологии и поведения человека. Генетика поведения за последние несколько десятков лет дала обширный материал, экспериментально подтверждающий наследование форм пищевого и полового поведения, эмоциональности и агрессивности. Датский психогенетик Н.Жуел-Нильсон, исследовав 12 пар разлученных близнецов, показал, что существует высокое внутрипарное сходство в мимике, голосе, походке, но вместе с тем — и большие индивидуальные различия в высокоспециализированных психических процессах и формах поведения (стиле межличностного общения, познавательных интересах и т.п.). Вопрос о том, наследственность или среда более важна для психологического и поведенческого развития человека, давно обсуждается в литературе, но до сих пор удовлетворительного решения не имеет. Очевидно, что развитие ребенка является результатом взаимодействия между врожденными биологическими особенностями организма и приобретенным жизненным опытом. Например, при нормальном процессе развития человеческий индивид довольно быстро усваивает речь. Но при любых обстоятельствах активная речь не может быть сформирована ранее одного года — полутора лет, т.е. того возраста, когда организм в силу генетических факторов достигнет определенного уровня биологической зрелости. Ни один ребенок, к примеру, не начал еще говорить целыми предложениями раньше годовалого возраста. Дети растут в человеческой среде, где окружающие их люди являются носителями развитых речевых умений и навыков. Они сознательно и бессознательно стимулируют ускоренное развитие соответствующей речевой способности у детей. У разных народов она формируется и проявляет себя по-разному. В развитых странах Европы, например, ребенок овладевает речью и начинает говорить в годовалом возрасте. Дети, родившиеся и выросшие в Сан Маркосе, отдаленной деревушке Гватемалы, и имеющие малый опыт общения со взрослыми людьми, овладевают речью гораздо позднее, в возрасте около двух лет. Ребенок всегда говорит на языке той страны, в которой он родился и вырос. Это, пожалуй, самое убедительное доказательство прямого влияния среды на психическое (речевое) развитие. Если же принять в расчет то обстоятельство, что именно речь составляет основу всех других видов психической деятельности, более сложных по уровню своего развития, то можно считать, что влияние среды на психическое развитие человеческого индивида в целом огромно. Вместе с тем — и для этого также имеются доказательства — оно существенно ограничено теми биологическими возможностями, которые предоставлены достигнутым уровнем зрелости организма. Проблема влияния биологического и социального начал на психическое развитие человека отчасти может быть решена за счет введения понятия сензитивного периода развития. Он обозначает возраст, в рамках которого при формировании тех или иных психологических качеств человека можно добиться наилучших педагогических результатов. Для речи, например, таким периодом является возраст от одного года до трех лет. Воздействие генетических факторов лучше всего просматривается в характере и динамике процесса созревания организма. Генотипически детерминированный процесс последовательных органических изменений, проходящих определенные стадии, относительно независим от событий, происходящих во внешней для организма среде. Мы говорим «относительно» потому, что такие изменения в определенной степени зависят от внешних условий, в которых происходит созревание организма. Такие условия могут замедлить или, напротив, ускорить процесс созревания, однако существенного влияния на последовательность стадий развития и его конечный результат не оказывают. Из-за неблагоприятных внешних условий развития человеческий организм может не достичь определенной стадии биологического созревания, но это не определяет его психического развития в будущем. Оно может идти ускоренно, если создать внешне благоприятные условия для дальнейшего созревания организма. На начальном этапе развития человека, в первые месяцы его жизни, ведущим является генотип. От него зависит созревание фиксированных программ развития психики и поведения в их элементарных формах, связанных с деятельностью организма. Определяющую роль в это время играют наиболее древние мозговые структуры (древняя и старая кора), управляющие созреванием общевидовых биологических структур, от которых зависят жизненно наиболее важные формы приспособительного поведения. Древние мозговые структуры поддерживают жизнедеятельность организма; способствуют накоплению информации, формированию умений и навыков, необходимых для перехода на следующий, более высокий уровень развития. Период жизни, о котором идет речь, охватывает первые два месяца и обычно характеризуется как новорожденность. Второй этап развития начинается с двухмесячного возраста. Он связан с началом функционирования и включением в работу коры головного мозга, активным развитием и дифференциацией ее структур. С возрастом количественно и качественно изменяется соотношение генотипических и средовых влияний, причем смена отношения генотипа и среды в детерминации психики и поведения сопровождается их качественной перестройкой, происходящей в ходе развития. Начиная с дошкольного возраста стабильно связанными с генотипом становятся индивидуальные различия в общительности, а также управление такими свойствами темперамента, как динамика непроизвольно совершаемых и автоматически регулируемых движений. К числу врожденных и довольно рано появляющихся относятся некоторые экспрессивные реакции человека, например способность улыбаться. Даже дети, родившиеся слепыми или глухими, отвечают улыбкой на ласку. Люди, принадлежащие любой из человеческих культур, часто от удивления поднимают брови. Вполне возможно, что и эта поведенческая реакция является врожденной. С возрастом, по-видимому, увеличивается вклад генотипа в индивидуальные интеллектуальные различия. Генетические влияния этого типа в детском и взрослом возрасте являются относительно устойчивыми. (Речь идет о формировании соответствующих индивидуальных различий, а не о тенденциях развития, рассматриваемых в качестве нормативных. Психологи же имеют обычно дело именно с последними.) Хотя роль негенетических факторов в формировании психологических и поведенческих индивидуальных различий довольно велика, тем не менее сам генотип редко обусловливает больше половины всех индивидуальных вариаций по соответствующим признакам. Поэтому, «принимая во внимание явную недостаточность наших знаний о генетической детерминации поведения человека, всякие спекуляции относительно природы наследственных изменений, обусловивших последние этапы эволюции нашего вида (имеются в виду люди. — Р.Н.), представляются преждевременными»1. Установлено, что в организме человека функционирует специальный нейроэндокринный аппарат генетической регуляции программы развития. Любая психическая функция благодаря его работе реализуется в пределах некоторого диапазона, заданного уровнем зрелости организма. Однако конечная ее форма в пределах этого диапазона зависит не столько от генотипа, сколько от условий окружающей среды, в частности от обучения и воспитания. В большинстве случаев, имея дело с конкретными психологическими характеристиками человека и формами поведения, невозможно сказать, в какой степени на них повлияли гены, в какой — среда. Сама по себе высокая наследуемость, установленная в исследованиях, проведенных на людях разных возрастов, не свидетельствует о возрастной преемственности именно генетических влияний. Наследственное воздействие на один и тот же психологический или поведенческий признак с возрастом человека может существенно измениться. Психологические и поведенческие характеристики, показывающие высокую устойчивость во времени, вовсе не обязательно связаны именно с генотипом. Наоборот, многие первоначально генетически обусловленные характеристики отнюдь не всегда являются неизменными в течение последующей жизни человека. Многие данные, которыми располагает психогенетика, указывают на следующее: когда в процессе развития обнаруживаются изменения в степени генетической обусловленности индивидуальных особенностей, они часто увеличиваются, а не уменьшаются. «Согласно современной генетике, — считает И.В.Равич-Щер-бо, — наследуется не конкретное значение признака, а некоторая способность иметь данный признак, и уже на основе этой наследственно обусловленной нормы реакции, во взаимодействии генотипа и среды формируется определенное фенотипиче-ское выражение признака»2. Из всех человеческих свойств минимальную зависимость от генотипа обнаруживают черты личности, выражающие отношение человека к людям и обществу. В целом же можно определенно утверждать, что среда в детерминации психического развития человека играет гораздо более важную роль, чем генотип. Даже среди известных свойств нервной системы человека нет ни одного, которое было бы полностью независимым от воздействий среды. «Чем выше в деятельности человека роль специфически человеческой, речевой произвольной саморегуляции, тем меньшую роль играют факторы генотипа, и, наоборот, чем она ниже, тем отчетливее индивидуальные особенности этой деятельности определяются наследственностью»1. Электрофизиологические исследования мозга человека, проведенные в последние годы, позволили установить важный факт о наследуемости электроэнцефалограммы (ЭЭГ) человека — картины электрической активности мозга в целом или отдельных его частей, а также выделить типы ЭЭГ и связать их с элементарными психическими и поведенческими явлениями. Соответствующие результаты в обобщенном виде представлены в табл. 4. Здесь показаны данные о влиянии врожденных особенностей ЭЭГ человека на отдельные стороны его психологии и поведения. Их можно описать так: 1. Люди с мономорфными α-волнами в среднем проявляют себя активными, эмоционально устойчивыми и надежными. Индивиды с особо выраженными признаками такого рода в ЭЭГ с достаточно высокой вероятностью обнаруживают такие поведенческие характеристики, как настойчивость, точность в работе, особенно в условиях стресса. У них развита кратковременная память, однако переработка информации протекает не очень быстро. 2. Лица с низкоамплитудной ЭЭГ демонстрируют низкую спонтанную активность. Те из них, для кого особенно характерен этот признак, часто оказываются экстравертами и склонны в своих мнениях и поступках ориентироваться на окружающих людей. Особенно хорошо развита у них пространственная ориентация. 3. Лица, ЭЭГ которых отличается диффузными β-волнами, делают большое количество ошибок в текстах на внимание, в видах деятельности, требующей повышенной аккуратности не смотря на низкую скорость работы. Их устойчивость к стрессу является довольно низкой. В результате этого они могут показывать сравнительно невысокие, не соответствующие их возможностям результаты тестирования, особенно в заданиях на проверку интеллекта и связанных с пространственной ориентацией. Таблица 4 Наследственные варианты энцефалограммы и их психологические следствия (по Фогель Ф., Мотульски А. Генетика человека. Проблемы и подходы: В 3 т. — Т. 3. — М., 1990)
|