Физиология растений контрольная работа. физиология растений. Взаимодействие факторов при фотосинтезе. Использование принципа взаимодействия факторов для регулирования фотосинтетической деятельности растений
 Скачать 274.5 Kb. Скачать 274.5 Kb.
|

 Факультет заочного образования КОНТРОЛЬНАЯ РАБОТА № 1 по дисциплине «ФИЗИОЛОГИЯ РАСТЕНИЙ» 3 семестр Вариант № 08 Выполнила Студентка Комарова К.С. Подготовка бакалавров Направление 35.03.10 Ландшафтная архитектура Профиль "Ландшафтное проектирование", Бакалавриат группа З-Л 204 Заочная форма обучения Проверил Доцент кафедры физиологии растений К.с.-х.н. Панфилова О.Ф. Москва, 2020 Реферат на тему: «Взаимодействие факторов при фотосинтезе. Использование принципа взаимодействия факторов для регулирования фотосинтетической деятельности растений» Фотосинтез — чрезвычайно сложный окислительно-восстановительный процесс, в результате которого вода окисляется, а диоксид углерода восстанавливается. Под окислением понимают процесс, при котором данная молекула отдает электроны другой молекуле, а под восстановлением — процесс, при котором данная молекула присоединяет электроны. Следовательно, в процессе фотосинтеза вода служит донором электронов и протонов, а диоксид углерода — их акцептором. Фотосинтез осуществляется путем последовательного чередования двух фаз: световой, протекающей с большой скоростью (10 ‘с) и не зависящей от температуры, и темновой, для химических реакций которой свет не нужен. Вторая фаза протекает медленнее первой и сильно зависит от температуры. Световые реакции осуществляются в мембранах тилакоидов, а темновые — в строме хлоропластов, т.е. они разобщены и во времени, и в пространстве. Внутренние факторы регуляции фотосинтеза К ведущим факторам внутренней регуляции фотосинтеза относятся скорость оттока ассимилятов из хлоропластов и концентрация хлорофилла в листьях. Интенсивность фотосинтеза листа в значительной степени зависит от запроса на ассимиляты со стороны потребляющих органов (акцепторов). Мощными потребителями являются меристематические ткани, в которых идет активный биосинтез. Растущее дерево активно потребляет ассимиляты на рост ствола, ветвей, корневых систем, на формирование плодов, семян, на биосинтез живицы у хвойных. Любое ускорение оттока ассимилятов усиливает интенсивность фотосинтеза. Неблагоприятные лесорастительные условия задерживают оттокассимилятов из хлоропластов и за счет этого снижают активность фотосинтеза. К внешним факторам, влияющим на фотосинтез, относят свет, концентрацию диоксида углерода, температуру и водоснабжение растений. Зависимость фотосинтеза от освещенности, температуры и водоснабжения как функция времени В природе напряженность внешних факторов непрерывно изменяется. Это влияет на фотосинтетический аппарат. Если освещенность уменьшается или увеличивается постепенно, то интенсивность фотосинтеза соответственно медленно снижается или растет. Однако, если внезапно ярко осветить листья, которые до этого находились в темноте, то интенсивность фотосинтеза изменяется непрямолинейно: после включения света она сначала увеличивается, затем падает и далее вновь растет до величины, соответствующей данной освещенности (рис. 1). Может пройти несколько минут, прежде чем скорость фотосинтеза достигнет величины, характерной для данной освещенности. Эта начальная задержка получила название индукции (от лат. induction — возбуждение), или индукционного эффекта. Впервые ее наблюдали в опытах с водорослями У. Остергаут (1918) и О. Варбург (1920). Затем возникновение индукции как неотъемлемой черты фотосинтеза было обнаружено в опытах с хлоропластами и протопластами. 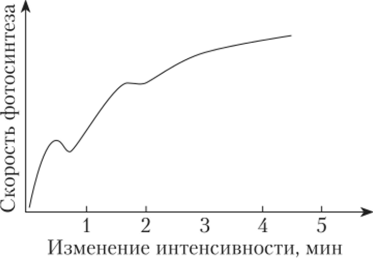 Рис. 1 Изменение интенсивности фотосинтеза во времени, вызванное переходом от темноты к свету (индукционный эффект) (по Ауфдемгартену, 1939). Поскольку большинство фотохимических процессов, например, транспорт электронов, идет с более или менее максимальной скоростью с самого начала, то индукция связана непосредственно с реакциями темновой фазы фотосинтеза [2]. В настоящее время индукцию объясняют, во-первых, необходимостью активировать некоторые ферменты светом. Во-вторых, для нормального течения реакций С3-цикла в хлоропластах должно накопиться некоторое количество молекул ФГК. Это увеличение количества ФГК как раз и происходит в течение первых 5 мин освещения. Индукционные эффекты наблюдаются также при переходе от длинноволнового к коротковолновому свету. При медленном обезвоживании интенсивность фотосинтеза уменьшается, но быстрое изменение водного режима листа, даже очень незначительное (0,06%), вызывает сначала увеличение, а потом уменьшение интенсивности фотосинтеза. Скорость нагрева или охлаждения воздуха тоже сказывается на временном ходе фотосинтеза. В условиях его медленного нагрева или охлаждения интенсивность фотосинтеза постепенно увеличивается или уменьшается. В условиях быстрого охлаждения уменьшению фотосинтеза предшествует его активация, а в условиях быстрого нагревания активации фотосинтеза предшествует его угнетение (рис.2). Следовательно, в условиях быстрой смены напряженности фактора интенсивность фотосинтеза изменяется двухфазно: кратковременная активация сменяется депрессией или активации предшествует угнетение. Причем даже оптимальные температуры способны при известной скорости [1] нагрева вызывать временное угнетение фотосинтеза. Оптимальные значения фактора способны влиять на фотосинтез как его экстремальные значения, если их напряженность изменяется внезапно. 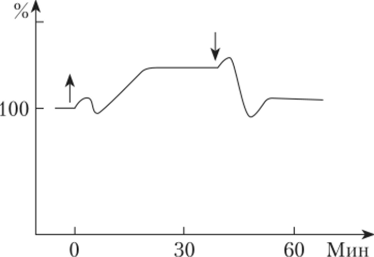 Рис. 2 Изменение интенсивности фотосинтеза во времени, вызванное внезапным изменением температуры листьев фасоли (по Беликову, Асафову, 1966). Взаимодействие факторов при для регулирования фотосинтеза В предыдущем разделе данной главы рассмотрено влияние на фотосинтез отдельно света, температуры и других внешних факторов. Однако в природе на растение все эти факторы действуют одновременно, одни влияют на световую фазу, а другие — на темповую или на обе. Темповая фаза фотосинтеза (С3-цикл) состоит примерно из 12 ферментативных реакций. Однако ее скорость определяется главным образом активностью двух ключевых реакций цикла: реакцией карбоксилирова- ниЯу скорость которой регулируется концентрацией С02, и реакцией восстановления ФГК в ФГАУ скорость которой зависит от содержания АТФ и НАДФН. Концентрация С02 в хлоропласте зависит от его концентрации в атмосфере и от сопротивления его диффузии, а количество АТФ и НАДФН — от транспорта электронов, вызванного светом. Кроме того, активность всех ферментативных реакций зависит от температуры. [3] Для начала рассмотрим влияние на фотосинтез любой пары факторов, например, интенсивности света и концентрации С02. Предположим, что мы увеличиваем освещенность, оставляя содержание углекислого газа на низком уровне. Это может наблюдаться днем в оранжерее. С того момента, когда интенсивность света достигнет такого уровня, при котором в единицу времени будет восстанавливаться вся поглощенная хлоропластами двуокись углерода, дальнейшее увеличение интенсивности фотосинтеза станет невозможным, и кривая зависимости фотосинтеза от освещенности будет параллельна оси абсцисс. Если повторим опыт при увеличенном содержании С02 в воздухе, то и перелом произойдет при большей, чем раньше, интенсивности света (рис.3). [1] 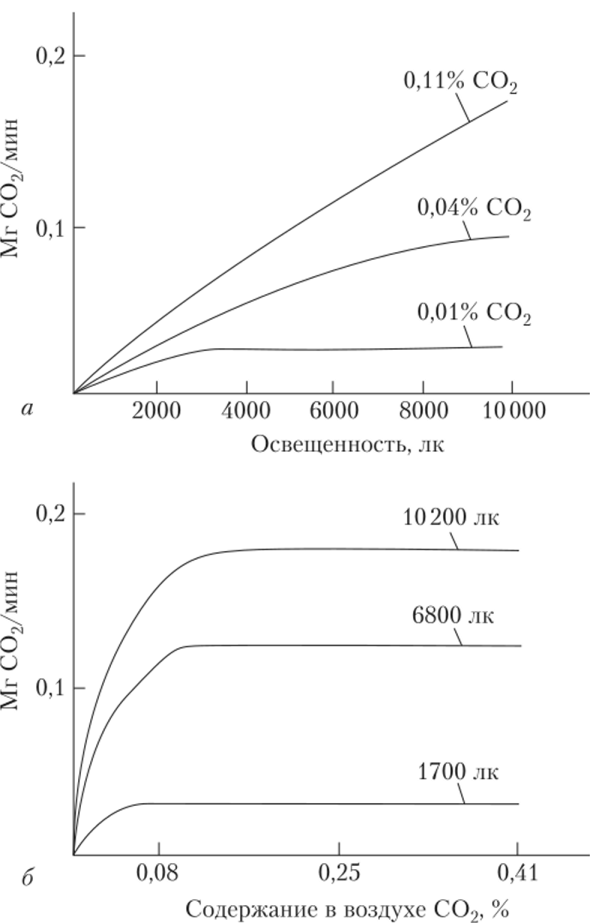 Рис. 3 Зависимость интенсивности фотосинтеза: а — от освещенности при разной концентрации углекислого газа; 6 — концентрации углекислого газа при разной освещенности. Таким образом, при малой концентрации углекислого газа интенсивность фотосинтеза нс зависит от освещенности, световое насыщение в этих условиях наступает раньше. При малой интенсивности света фотосинтез не зависит от концентрации С02. Чем меньше освещенность, тем скорее наступает световое насыщение. Световая компенсационная точка наступает раньше при более высокой концентрации С02, так как в этих условиях увеличивается скорость фотосинтеза. Анализ кривых на рис.3 показывает, что или освещенность, или концентрация С02 могут ограничивать фотосинтез. Влияние температуры на фотосинтез зависит от того, является ограничивающим фактором свет или концентрация углекислого газа. При низких интенсивностях света температура не влияет на интенсивность фотосинтеза. При большой интенсивности света более высокие температуры увеличивают интенсивность фотосинтеза, если достаточно С02. Если С02 мало, то увеличение температуры не повлияет на интенсивность фотосинтеза при увеличении освещенности. При повышении температуры компенсационная точка наступает при большей освещенности, так как в этих условиях дыхание активируется сильнее, чем фотосинтез. Понижение температуры приводит к более раннему достижению как светового, так и углекислотного компенсационных пунктов. Изучая влияние на фотосинтез нескольких факторов одновременно, сделали вывод, что один из факторов, находящийся в минимуме, ограничивает влияние других факторов. Эта зависимость получила название закона ограничивающих факторов (Либих, Германия, 1840). Именно эти наблюдения позволили еще в 1905 г. Ф. Блэкману (Англия) сделать вывод о том, что фотосинтез состоит из двух фаз: зависимой от света (световой фазы) и зависимой от концентрации углекислого газа и температуры (темновой фазы). При малой освещенности фотосинтез увеличивается пропорционально интенсивности света и почти не зависит от факторов, влияющих на темповые реакции фотосинтеза. Это понятно, так как при малой освещенности темповая фаза идет медленно. Таким образом, для получения максимального урожая необходимо поддерживать оптимальное соотношение всех факторов, влияющих на фотосинтез. [1] - Цит. по: Беликов П. С., Дмитриева Г. А. Физиология растений. Список используемых источников. Беликов П.С. - Физиология растений. Учеб. пособие для вузов по направлениям "Агрономия", "Биология" и спец. "Физиология растений" / П. С. Беликов, Г. А. Дмитриева. - М.: Издательство Ростовского университета Дружбы народов, 1992. – 248 с. Шумакова Е.В. - Ботаника и физиология растений: учебник для студ. учреждений сред. проф. образования / Е.В.Шумакова. — М.: Издательский центр «Академия», 2013. — 208 с. Интернет источник: https://cyberlesson.ru/kakie-processy-proishodjat-v-temnovuju-fazu/ (дата обращения 13.04.2021). Напишите, какие органеллы клетки называются полуавтономными и почему? Полуавтономные органоиды- органоиды, содержащие собственные ДНК, все типы РНК, рибосомы, способные синтезировать белки, а также преобразовывать Е в ЕАТФ (синтез АТФ). К таким органоидам относятся двумембранные органоиды: митохондрии и хлоропласты. Митохондрии и пластиды способны к самовоспроизведению и никогда не образуются из других компонентов клетки. Вне клеток они существовать не могут. Напишите, как изменяется водный дефицит растений в течение суток в летний день при недостаточном водообеспечении растений. В жаркие летние дни корни не успевают покрывать расход воды на возрастающую транспирацию, что приводит к водному дефициту в растении. Полуденный водный дефицит и временное завядание не причиняют растению особого вреда. Их значительному увеличению препятствует способность растения к частичному (рис. 7, В) или полному (рис. 7, Г) гидроактивному закрыванию устьиц и снижению транспирации. К вечеру водный дефицит снижается, а в ночные часы за счет активной деятельности корневой системы водный баланс полностью восстанавливается. Во время летней засухи устьица ненадолго открываются лишь в утренние часы, и идет только кутикулярная транспирация (рис. 7, Д).  В условиях засухи, когда в почве почти не остается доступной для растения влаги, водный баланс за ночь не восстанавливается, наблюдается длительное увядание растения. Такой не покрываемый к утру водный дефицит получил название остаточного водного дефицита. Рассчитайте коэффициент водопотребления насаждения при урожайности 50ц/га и сезонном водопотреблении 4500м3/га. Коэффициент водопотребления характеризует эффективность использования воды и определяется как количество сезонного водопотребления на единицу урожая. При этом, обе величины должны иметь одинаковые единицы измерения. 1м3 = 1т воды. Урожай также выражаем в т/га: 50ц/га = 5т/га И получаем коэффициент водопотребления: 4500 / 5 = 900. Сравнение с литературными данными позволяет сделать вывод о достаточно эффективном расходовании воды насаждением. |
