ЗАНЯТИЕ 7. ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ. Занятие Физиологические особенности скелетных и гладких мышц
 Скачать 1.78 Mb. Скачать 1.78 Mb.
|

|
Занятие 7. Физиологические особенности скелетных и гладких мышц Классификация мышечных тканей, их физиологические свойства. Мышечные ткани - ткани различного происхождения и строения, для которых ведущей функцией является сократимость, а основными органеллами - миофибриллы. В организме человека по структуре и физиологическим свойствам выделяют 3 типа мышечной ткани: 1. Поперечно-полосатую (скелетную). 2. Гладкую. 3. Сердечную Поперечно-полосатая (скелетная) мышечная ткань является произвольной возбудимой тканью. Она формирует скелетные мышцы, мышцы ротовой полости, верхней трети глотки, наружные сфинктеры мочеиспускательного канала и прямой кишки. Тканевыми элементами служат мышечные волокна - симпласты, которые содержат 4 элемента, обеспечивающих сократительную функцию мышц: а) миофибриллы, образованные параллельно ориентированными нитями актина и миозина; б) тропонин-тропомиозин регулирующий комплекс (управляет актом сокращения); в) саркоплазматический ретикулум, образующий систему T-трубочек и L-каналов, которые содержат ионизированный кальций, запускающий сокращение; г) систему энергетического обеспечения (митохондриальные комплексы). Скелетные мышцы обеспечивают сохранение позы, взаимное расположение частей тела и перемещение тела в пространстве, дыхание, прием пищи. Гладкомышечная ткань является непроизвольно возбудимой. Она формирует мышечные оболочки внутренних органов, все внутренние сфинктеры, входит в состав стенки сосудов. Тканевым элементом является гладкомышечная клетка - миоцит веретеновидной или звездчатой формы. Сократительные элементы представлены: а) миозиновыми нитями, проходящими вдоль оси клетки и актиновыми нитями, образующими 3-мерную сеть; б) слабо развитым тропонин-тропомиозиновым комплексом; в) не развитым эндоплазматическим ретикулумом, (необходимый для инициирования мышечного сокращения ионизированный кальций поступает из внешней среды клетки); г) системой энергетического обеспечения. Гладкие мышцы обеспечивают перистальтику желудка, кишечника, мочеточника, маточной трубы, тонус кровеносных и лимфатических сосудов, опорожнение кишечника, мочевого пузыря. В функциональном отношении различают фазные мышечные волокна (обеспечивают движения, связанные с перемещением тела в пространстве) и тонические мышечные волокна (обеспечивают длительно протекающие сократительные процессы, например, сохранение позы). В фазных волокнах генерируется ПД, который и распространяется по мембране. В тонических волокнах возбуждение возникает градуально в зависимости от силы стимула. Фазные мышечные волокна в свою очередь подразделяются на быстрые и медленные мышечные волокна. Различия между ними заключаются в:
Количество медленных и быстрых мышечных волокон в разных мышцах неодинаково и у разных людей оно тоже различно. Соотношение мышечных волокон генетически запрограммировано. Переход быстрых мышечных волокон в медленное и, наоборот, в течение жизни не происходит. Все типы мышц обладают некоторыми свойствами: 1. Возбудимость. 2. Проводимость. 3. Сократимость – изменение длины или напряжения. Способность расслабляться. 4. Эластичность – способность развивать напряжение при растяжении В естественных условиях деятельность мышц носит рефлекторный характер. Зарегистрировать электрическую активность мышцы можно с помощью электромиографа. Электромиография используется в спортивной медицине. Формы и типы мышечного сокращения. Режимы сокращения мышц.  Различают несколько форм и типов мышечных сокращений. 1. Динамическая форма мышечного сокращения. При таком типе сокращений изменяется длина мышцы, но не изменяется напряжение. Эта форма включает два типа: а) Изотонический тип или концентрационный (мышца укорачивается, но не изменяет своего напряжения). Например, ходьба. б) Эксцентрический тип. Если нагрузка на мышцу больше, чем ее напряжение, то мышца растягивается. Например, при опускании тяжелого предмета. 2 Статическая форма мышечного сокращения. Эта форма наблюдается при поддержании позы или преодолении силы земного притяжения. Данная форма включает один тип мышечного сокращения – изометрический. При изометрическом сокращении мышца изменяет свое напряжение, но не изменяет длины. 3. Смешанная форма (ауксотонические, ауксометрические сокращения). Деление на формы и типы мышечных сокращений является условным т.к. все сокращения являются смешанными. Однако преобладает какой-то один тип. Режимы сокращения мышц. Характер или режим сокращения мышцы зависит от частоты импульсов, которые поступают от мотонейрона. Выделяют одиночные и тетанические мышечные сокращения. Если на мышцу подействовать одиночным импульсом, то происходит одиночное мышечное сокращение, в котором выделяют несколько фаз: 1. Латентный (скрытый) период – время после действия раздражителя до начала сокращения. 2. Фаза укорочения (при изотоническом сокращении) или фаза напряжения (при изометрическом сокращении). 3. Фаза расслабления. 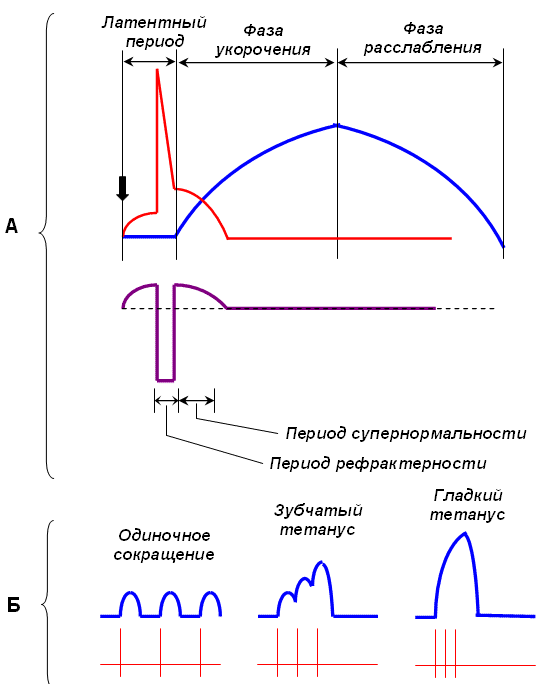 Одиночное мышечное сокращение характеризуется незначительной утомляемостью, но при этом мышца не способна реализовать свои возможности. Если на мышечное волокно воздействуют два быстро следующих друг за другом импульса, то сокращения накладываются и возникает сильное сокращение. Наложение двух следующих друг за другом импульсов называется суммацией. Выделяют два вида суммации: 1. Если второй раздражитель поступает в момент, когда мышца начала расслабляться, то кривая имеет вершину отдельную от вершины первого сокращения. Этот вид суммации называется неполной. 2. Если второй раздражитель поступает в момент, когда сокращение мышцы еще не дошло до вершины т.е. мышца не начала расслабляться, то оба сокращения сливаются в единое целое. Этот вид суммации называется полной. Длительное и сильное сокращения мышцы, под влиянием ритма импульсов с последующим расслаблением называется тетанусом. У человека тетанус можно получить при частоте 50—70 имп/сек. Выделяют два вида тетануса: 1. Зубчатый. Возникает при малой частоте подачи импульсов (до 150 имп/cек). 2. Гладкий. Возникает при высоком ритме подачи импульсов (более 150 имп/cек). При этом различают оптимальный и пессимальный ритмы работы мышцы. Так, если частота подачи и сила импульсов вызывает максимальный сократительный эффект, то это оптимальный ритм работы. Оптимальный ритм работы формируется через фазу экзальтации (т.е. супернормальности). Если частота подачи импульсов и сила раздражителя слишком велики, то это вызывает снижение силы сокращения. Такой ритм называется пессимальным. Этот ритм работы мышцы формируется через фазу абсолютной рефрактерности. Контрактура. Иногда, в работе мышцы, наблюдается стойкое непрерывное стационарное обратимое сокращение мышцы с сильно замедленным его расслаблением. Такое сокращение мышцы называется контрактура (судорога). От тетануса она отличается отсутствием распространяющегося потенциала действия вдоль мышцы. Выделяют 3 вида контрактур: 1. Калиевая. Развивается, если в окружающей мышечное волокно жидкости накапливается много ионов калия. 2. Кофеиновая. Под влиянием высокой концентрации кофеина внутрь мышечного волокна поступают ионы кальция и вызывают длительное сокращение мышцы. 3. Посттетаническая. Это остаточное укорочение мышцы после снятия действия раздражителя. Например, если долго нести тяжелую сумку и не разжимать пальцы рук, то после освобождения от сумки пальцы сразу не разгибаются. Сила и работа мышечного волокна. Двигательные единицы. Величина сокращения (сила мышцы) зависит от морфологических свойств и физиологического состояния мышцы: 1. Исходной длины мышцы (длинны покоя). Чем сильнее мышца растянута в покое, тем сильнее сокращение (закон Франка-Старлинга). 2. Диаметра мышцы или поперечного сечения. Выделяют два диаметра: а) анатомический диаметр – поперечное сечение мышц. б) физиологический диаметр – перпендикулярное сечение каждого мышечного волокна. Чем больше физиологическое сечение, тем большей силой обладает мышца. Сила мышцы измеряется весом максимального груза поднятого на высоту или максимальным напряжением, которое она способна развить в условиях изометрического сокращения. Измеряется в килограммах или ньютонах. Методика измерения силы мышцы называется динамометрия. Выделяют два вида силы мышцы: 1. Абсолютная сила – отношение максимальной силы к физиологическому диаметру. 2. Относительная сила – отношение максимальной силы к анатомическому диаметру. При сокращении мышца способна выполнять работу. Работа мышцы измеряется произведением поднятого груза на величину укорочения. Работа мышцы характеризуется мощностью. Мощность мышцы определяется величиной работы в единицу времени и измеряется в ваттах. Наибольшая работа и мощность достигается при средних нагрузках. Двигательные единицы Мотонейрон с группой иннервируемых им мышечных волокон составляет двигательную единицу. Аксон мотонейронов может ветвиться и иннервировать группу мышечных волокон. Так, один аксон может иннервировать от 10 до 3000 мышечных волокон. 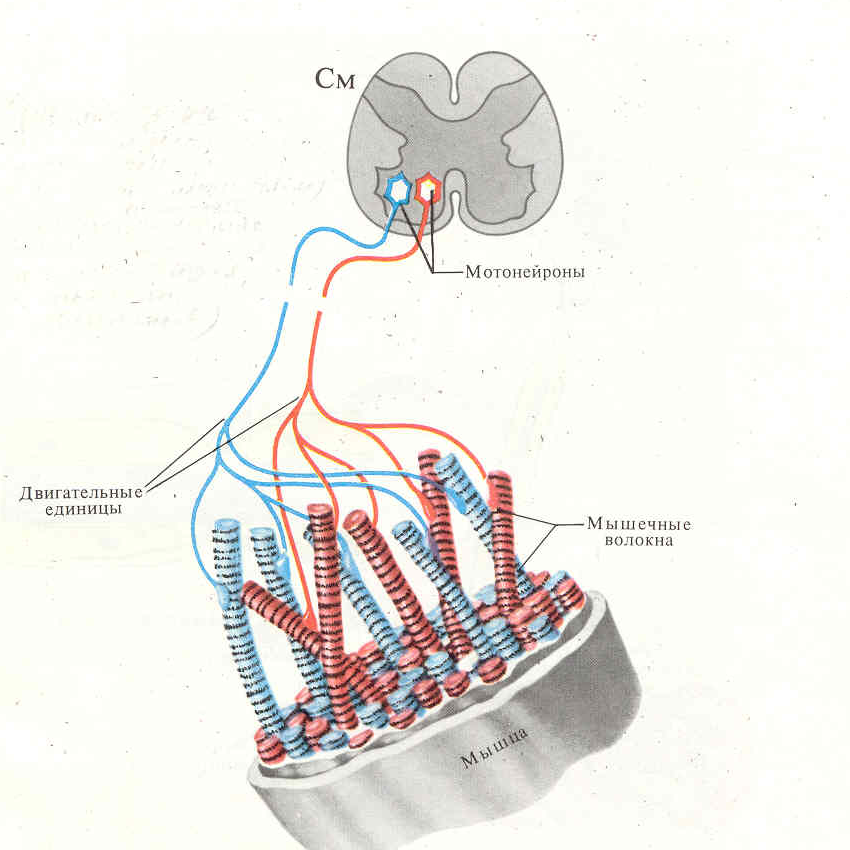 Различают двигательные единицы по строению и функциям. По строению двигательные единицы делятся на: 1. Малые двигательные единицы, которые имеют малый мотонейрон и тонкий аксон, способный иннервировать 10-12 мышечных волокон. Например, мышцы лица, мышцы пальцев рук. 2. Большие двигательные единицы представлены крупным телом мотонейрона, толстым аксоном, который способен иннервировать более 1000 мышечных волокон. Например, четырехглавая мышца. По функциональному значению двигательные единицы делятся на: 1. Медленные двигательные единицы. Они включают малые двигательные единицы, являются легко возбудимыми, характеризуются невысокой скоростью распространения возбуждения, в работу включаются первыми, но при этом они практически не утомляемы. 2. Быстрые двигательные единицы. Они состоят из больших двигательных единиц, плохо возбудимы, обладают большой скоростью проведения возбуждения. Обладают высокой силой и скоростью ответа. Например, мышцы боксера. Эти особенности двигательных единиц обусловлены рядом свойств. Мышечные волокна, которые входят в двигательные единицы, имеют сходные свойства и различия. Так, медленные мышечные волокна обладают: 1. Богатой капиллярной сетью. 2. Содержат много миофибрилл. 3. Содержит много миоглобина (т.е. способны связывать большое количество кислорода). 4. В них содержится много жиров. Благодаря этим особенностям эти мышечные волокна обладают высокой выносливостью, способны к небольшим по силе сокращениям, но длительным по времени. Отличительные особенности быстрых мышечных волокон: 1. Содержат большее, чем медленные волокна, миоглобина. 2. Обладают большей скоростью и силой сокращения. 3. Содержат мало капилляров. 4. Содержат мало жиров. В связи с этими особенностями быстрые мышечные волокна быстро утомляемы, но обладают большой силой и высокой скоростью ответа. 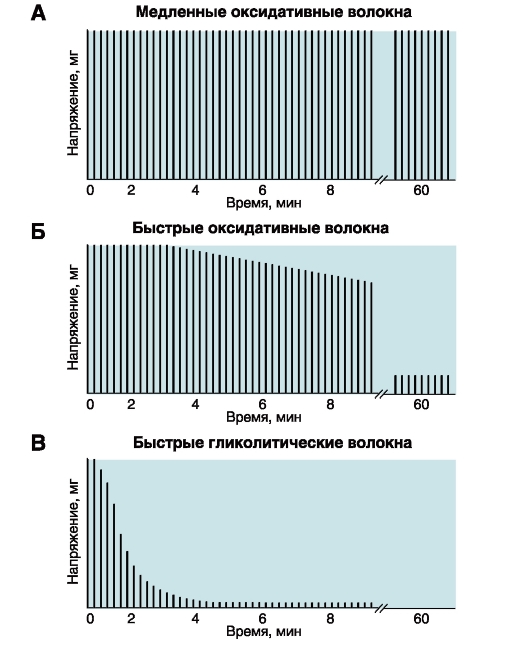 Типы волокон скелетных мышц. Скорость развития утомления в волокнах трех типов. Структура мышечного волокна. Теория сокращения мышц (скольжение нитей). Расслабление и утомление мышц. Тренировка. Гипертрофия и атрофия мышц. Скелетные мышцы состоят из клеток (мышечных волокон). Каждое мышечное волокно - это многоядерная клетка цилиндрической формы диаметром 20-80 мкм и длиной от нескольких миллиметров до десятков сантиметров, соответствующей длине мышцы. Многоядерные волокна исчерченных мышц образуются путём слияния одноядерных клеток - миобластов. В мышце волокна объединены в пучки по 20-40 волокон и отделены друг от друга соединительной тканью. К этой оболочке со стороны мышечной клетки прилегает плазматическая мембрана - сарколемма. Содержимое мышечного волокна называется саркоплазмой. В составе саркоплазмы выделяют миофибриллы, миоглобин, гликоген, саркоплазматический ретикулум (система продольных Т-трубочек и вытянутых мешочков, которые содержат кальций). В каждом волокне имеется около 2000 миофибрилл диаметром 1-2 мкм, проходящих от одного конца волокна до другого. Миофибриллы сгруппированы в пучки и проходят через все волокно, не прерываясь. Они разделяются на темные и светлые диски. Темные диски называются анизотропными, а светлые – изотропными. Светлые полосы в центре имеют Z-мембрану, а темные Н-полоску. Участок миофибриллы между двумя Z-мембранами длинной 2-3 мм называется саркомером. 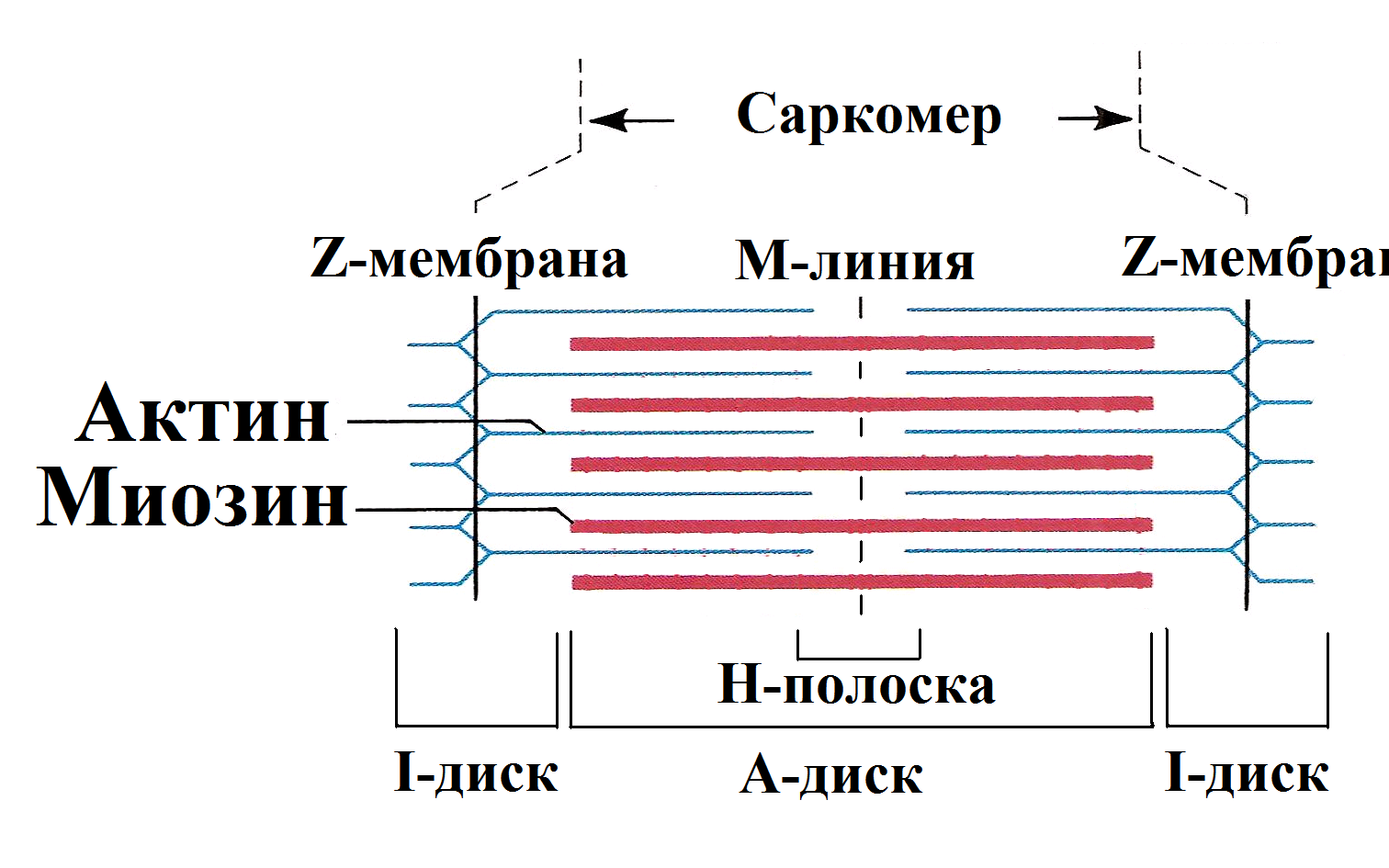 Каждый саркомер состоит из перекрывающихся толстых сократительных белков (нитей) – миозиновых и тонких (актиновых) Миозин – образован из тяжелых и легких цепей (по мол.массе), которые образуют хвост и 2 головки. Каждый толстый филамент имеет 500 головок. На каждой головке имеется 2 участка связывания – один для актина, другой – для АТФ. Сайт связывания АТФ обладает АТФ-азной активностью для гидролиза АТФ длиной 1,6 мкм и диаметром 16 нм. Актин — две спирально скрученные нити фибриллярного актина (F актин), образованные из глобулярного актина G актин длиной 1 мкм и диаметром 8 нм. Тонкие нити прикрепляются к Z-дискам и образуют изотропную I-полосу. Толстые нити в центре саркомера образуют анизотропную А-полосу. В области перекрытия миофибриллы образуют гексогональную структуру, в которой одну миозиновую нить окружают шесть актиновых. Взаимодействие между толстыми и тонкими нити осуществляется с помощью головок, которые выступают из миозиновых филаментов. Кроме актина, тонкая актиновая нить содержит фибриллярный белок тропомиозин и глобулярный белок тропонин. Три белка в тонкой нити находятся в определённом соотношении: на 7 глобул двойной нити актина приходится по одной молекуле тропомиозина и тропонина. Стержнеподобные молекулы тропомиозина длиной около 40 нм располагаются в бороздках актиновой суперспирали и взаимодействуют конец к концу. С каждой молекулой тропомиозина связывается молекула тропонина, состоящая из трёх субъединиц: тропомиозин - связывающей субъединицы, ингибиторной субъединицы и кальций-связывающей субъединицы, имеющей четыре кальцийсвязывающих центра. В процессе гидролиза АТФ миозином освобождается энергия, обеспечивающая движение головок миозина. Теория сокращения мышц (скольжение нитей) Хаксли Сокращение мышцы связано с возникновением потенциала действия на мембране мышечного волокна, который распространяется по сарколемме и поступает во внутрь волокна. В СПР имеются потенциалзависимые каналы (рианодиновые рецепторы) для Са2+, Са2+ выходит в цитоплазму по градиенту концентрации (в цитоплазме 10-8), (в СПР 10 -4). 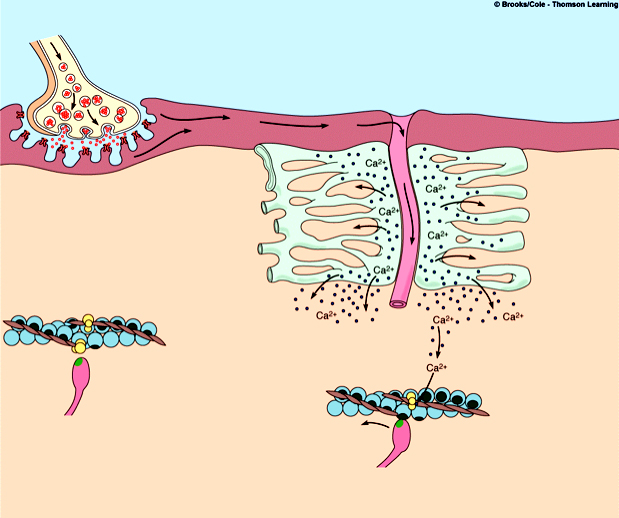 Одиночный цикл образования поперечных мостиков 1. Выход ионов Са из саркоплазматического ретикулума (СПР), связывание с С-субъединицей тропонина, смещение тропонина на актиновой нити, освобождение актиновых центров актина для связывания с миозином. 2. Связывание головок миозина с актином – образование поперечных мостиков (головка миозина энергизирована: АТФ расщеплена, но АДФ и фосфат не отсоединились. 3. Завершение гидролиза АТФ (АДФ и фосфат отсоединяются), поворот головки миозина, скольжение нитей актина, укорочение саркомера. 4. Присоединение новой молекулы АТФ, разъединение мостиков между актином и миозином. 5. Энергизация головки миозина – частичный гидролиз АТФ (АДФ и фосфат не отделяются), восстановление исходной конформации головки миозана. 6. Откачивание ионов Са назад в СПР. Кальций в саркоплазме активирует Са-АТФ-азу, кальциевый насос осуществляет активный перенос этого иона в СПР. Возврат мышцы в исходное, растянутое положение определяется массой костей скелета, связанных с данными мышцами и создающими растягивающее усилие после прекращения процесса сокращения. Вторым моментом является упругость мышцы, которая преодолевается в момент сокращения. 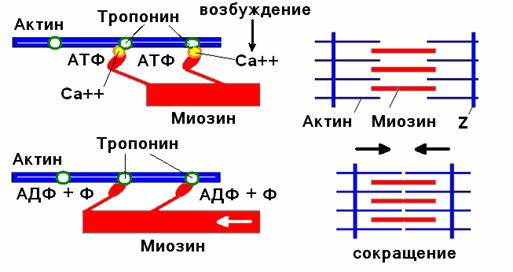 Затраты АТФ. Энергия АТФ расходуется на: – образование поперечных мостиков, осуществляющих продольное скольжение актиновых филаментов (основная часть энергии гидролиза АТФ); – Ca2+-насос: выкачивание Ca2+ из саркоплазмы в саркоплазматический ретикулум после окончания сокращения; – Na+/K+-насос: перемещение ионов натрия и калия через мембрану МВ для обеспечения соответствующего ионного состава вне- и внутриклеточной среды. Энергетические потребности. Мышечное сокращение требует значительных энергетических затрат. Основной источник энергии — гидролиз макроэрга АТФ. В митохондриях в процессе цикла трикарбоновых кислот и окислительного фосфорилирования генерируется АТФ. Гликоген запасaется в сaркоплазме в виде включений. Анаэробный гликолиз сопряжён с синтезом АТФ. Креатинфосфокиназа, связанная в области М-линии, катализирует перенос фосфата от фосфокреатина на АДФ с образованием креатина и АТФ. Миоглобин, как и Hb, обратимо связывает кислород. Запасы кислорода необходимы для синтеза АТФ при длительной непрерывной работе мышцы. На один рабочий цикл затрачивается 1 молекула АТФ. В МВ концентрация АТФ равна 4 ммоль/л. Такого запаса энергии достаточно для поддержания сокращения не более 1–2 сек. 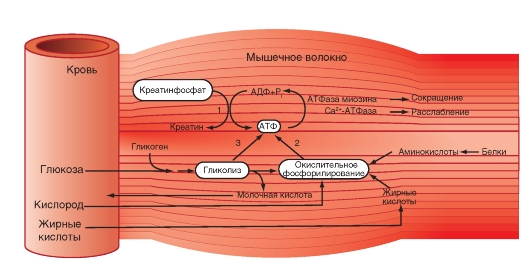 Энергетический метаболизм скелетной мышцы. Три ресурса для образования АТФ во время мышечного сокращения: 1 - креатинфосфат; 2 - окислительное фосфорилирование; 3 - гликолиз 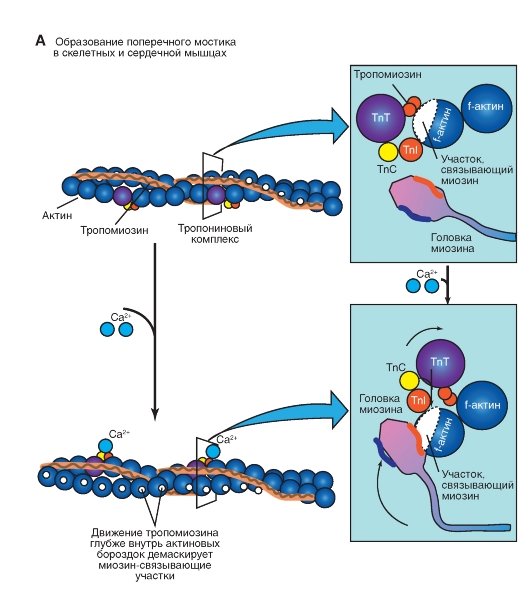 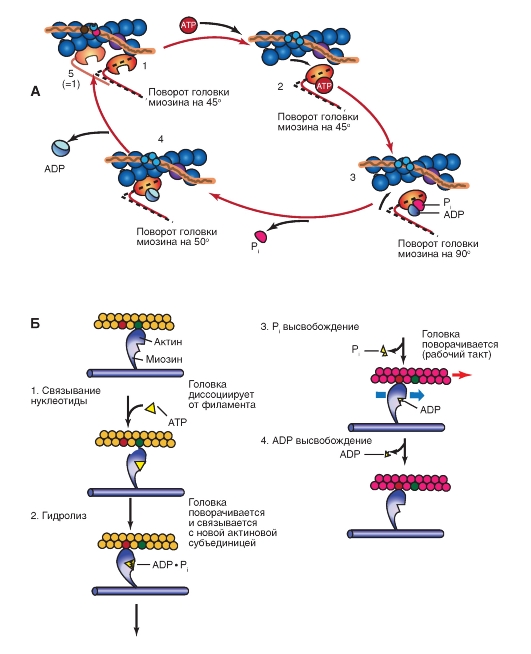 Механизм сокращения. Представлен рабочий цикл поперечных мостиков - головки миозина. Утомление мышц. Утомление – временное снижение работоспособности, наступающее в процессе выполнения мышечной работы и исчезающее после отдыха. Причины утомления: 1. Накопление продуктов обмена (молочная кислота) в мышцах, что ведет к угнетению генерации потенциала действия. 2. Кислородное голодание, т.е. к мышце не успевает доставляться кислород. 3. Истощение энергии. 4. Центрально-нервная теория утомления. По этой теории утомление нервных клеток наступает быстрее, чем мышц. 5. Утомление синапсов, через которые импульсы передаются к мышцам. В целом нет ни первой, ни последней причины. Все они действуют одновременно. Изучая влияние СНС на скелетную мышцу лягушки А.Г. Гинецинским было установлено, что если на мышцу утомленную до полной невозможности сокращаться подействовать стимуляцией симпатических волокон которые иннервируют эту мышцу, а затем начать стимулировать ее через моторные нервы сокращения восстанавливались. Выяснилось, что эти изменения связанны с тем, что под влиянием СНС в мышце происходит укорочение хроноксии, укорачивается время передачи возбуждения, повышается чувствительность к ацетилхолину, повышается потребление кислорода. Данное явление получило название феномена Орбели-Геницинского. Тренировка. Гипертрофия и атрофия мышц Тренировка - система физических упражнений и режим их реализации. Ее целью является увеличение долговременности и объема выполняемой мышечной работы. Это достигается за счет увеличения в организме массы мышечной ткани, перестройки обмена веществ в мышце, изменения в системах кровоснабжения мышц, дыхания, нервной регуляции мышечной деятельности. Метаболические изменения в мышце позволяют выполнять работу в условиях сниженного поступления к мышце кислорода (в анаэробных условиях), увеличиваются кислородная емкость крови, минутный объем сердечного выброса, минутный объем дыхания, скорость кровотока. Систематические тренировки приводят к гипертрофии мышцы. Выделяют два вида гипертрофии: 1. Миофибриллярный тип. Развивается при статической работе (поднятие тяжести). При этом типе гипертрофии увеличивается число миофибрилл и значительно увеличивается сила мышцы. Например, тяжелоатлеты. 2. Саркоплазматический тип – увеличение объема саркоплазмы (гликогена, креатининфосфата, миоглобина, числа капилляров). При этом типе гипертрофии развивается выносливость. Например, бегуны на длинной дистанции. Атрофия мышцы развивается при ее бездеятельности. Атрофия способствует постельный режим, перерезка сухожилий, заболевания нервной системы, гипсовая повязка. Физиологические особенности гладких мышц. Строение гладкой мышцы По структуре гладкая мышца отличается от поперечнополосатой скелетной мышцы и мышцы сердца. Она состоит из клеток веретенообразной формы длиной от 10 до 500 мкм, шириной 5-10 мкм, содержащих одно ядро. Гладкомышечные клетки лежат в виде параллельно ориентированных пучков, расстояние между ними заполнено коллагеновыми и эластическими волокнами, фибробластами, питающими магистралями. Мембраны прилежащих клеток образуют нексусы, которые обеспечивают электрическую связь между клетками и служат для передачи возбуждения с клетки на клетку. Кроме того плазматическая мембрана гладкомышечной клетки имеет особые впячивания - кавеолы, благодаря которым площадь мембраны увеличивается на 70%. Снаружи плазматическая мембрана покрыта базальной мембраной. Комплекс базальной и плазматической мембраны называют сарколеммой. В гладкой мышцы отсутствуют саркомеры. Основу сократительного аппарата составляют миозиновые и актиновые миофиламенты. В ГМК актиновых миофиламентов намного больше, чем в поперечно-полосатом мышечном волокне. Соотношение актин/миозин = 5:1. Толстые и тонкие миофиламеты распылены по всей саркоплазме гладкого миоцита и не имеют такой стройной организации, как в поперечно-полосатой скелетной мышце. При этом тонкие филаменты прикрепляются к плотным тельцам. Некоторые из этих телец расположены на внутренней поверхности сарколеммы, но большинство из них находятся в саркоплазме. Плотные тельца состоят из альфа-актинина – белка обнаруженного в структуре Z-мембраны поперечнополосатых мышечных волокон. Некоторые из плотных телец расположенных на внутренней поверхности мембраны соприкасаются с плотными тельцами прилегающей клетки. Тем самым сила, создаваемая одной клеткой может передаваться следующей. Толстые миофиламенты гладкой мышцы содержат миозин, а тонкие – актин, тропомиозин. При этом в составе тонких миофиламентов не обнаружен тропонин. Гладкие мышцы встречаются в стенках кровеносных сосудах, коже и внутренних органах. Гладкая мышца играет важную роль в регуляции
Классификация гладких мышц:
Мультиунитарная гладкая мышца.
Унитарная гладкая мышца (висцеральная).
Важной особенность гладкомышечных клеток является их способность к самовозбуждению (автоматии), то есть они способны генерировать потенциал действия без воздействия внешнего раздражителя. Постоянный мембранный потенциал покоя в гладких мышцах отсутствует, он постоянно дрейфует и в среднем составляет -50мВ. Дрейф происходит спонтанно, без каких-либо влияний и когда мембранный потенциал покоя достигает критического уровня возникает потенциал действия, который и вызывает сокращение мышцы. Продолжительность потенциала действия достигает нескольких секунд, поэтому и сокращение тоже может длиться несколько секунд. Возникшее возбуждение затем распространяется через нексус на соседние участки вызывая их сокращения. Спонтанная (независимая) активность связана с растяжением гладкомышечных клеток и когда они растягиваются возникает потенциал действия. Частота возникновения потенциалов действия зависит от степени растяжения волокна. Например, перистальтические сокращения кишечника усиливаются при растягивании его стенок химусом. Унитарные мышцы в основном сокращаются под влиянием нервных импульсов, но иногда возможны и спонтанные сокращения. Одиночный нервный импульс не способен вызывать ответной реакции. Для ее возникновение необходимо суммировать несколько импульсов. Для всех гладких мышц при генерации возбуждения характерна активация кальциевых каналов, поэтому в гладких мышцах все процессы идут медленнее по сравнению со скелетной. Скорость проведения возбуждения по нервным волокнам к гладким мышцам составляет 3-5 см в секунду. Одним из важных раздражителей инициирующих сокращение гладких мышц является их растяжение. Достаточное растяжение гладкой мышцы обычно сопровождается появлением потенциалов действия. Таким образом, появлению потенциалов действия при растяжении гладкой мышцы способствует два фактора:
Данное свойство гладкой мышцы позволяет ей автоматически сокращаться при растяжении. Например, во время переполнения тонкого кишечника возникает перистальтическая волна, которая и продвигает содержимое. Сокращение гладкой мышцы. Агонист (адреналин, норадреналин, ангиотензин, вазопрессин) через свой рецептор активирует G белок (Gp), который в свою очередь активирует фосфолипазу С. Фосфолипаза С катализирует образование инозитолтрифосфата (ИТФ). Инозитолтрифосфат стимулирует высвобождение Ca2+ из кальциевых депо. Са2+связывается с кальмодулином, активирующим киназу миозина, которая фосфорилирует легкую цепь миозина. Это приводит к гидролизу АТФ и запускает цикл образования поперечных мостиков. В гладкой мышце движение актомиозиновых мостиков является более медленным процессом. Распад молекул АТФ и высвобождение энергии, необходимой для обеспечения движения актомиозиновых мостиков происходит не так быстро как в поперечнополосатой мышечной ткани. Экономичность энергозатрат в гладкой мышце является чрезвычайно важным в общем потреблении организмом энергии, так как, кровеносные сосуды, тонкий кишечник, мочевой пузырь, желчный пузырь и другие внутренние органы постоянно находятся в тонусе. Во время сокращения гладкая мышца способна укорачиваться вплоть до 2/3 ее первоначальной длины (скелетная мышца от 1/4 до 1/3 длины). Это позволяет полым органам выполнять свою функцию изменяя свой просвет в значительных пределах. 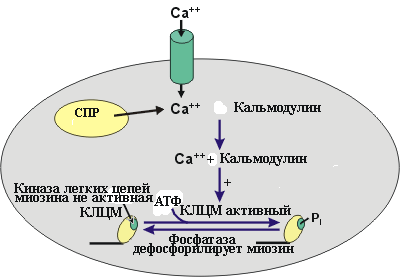 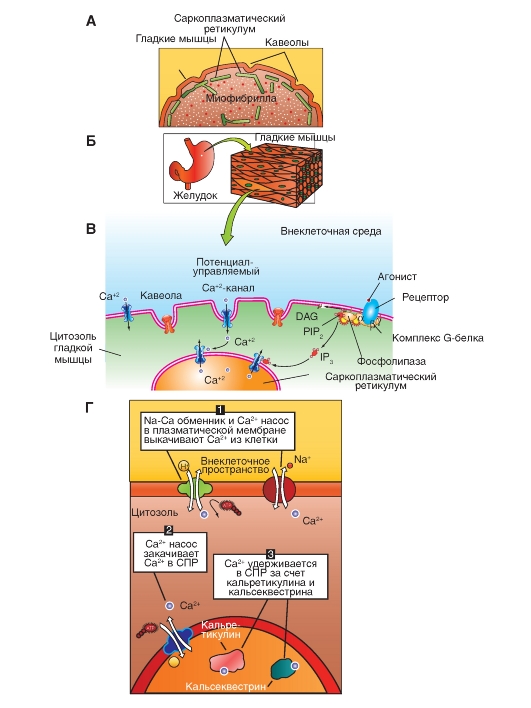 Саркоплазматический ретикулyм гладких мышц. А - структура саркоплазматического ретикулума. Б - источники поступления кальция через ионные каналы. В - источники поступления кальция через насосы и обменники Расслабление. При уменьшении содержания Ca2+ в миоплазме (постоянное откачивание Ca2+ в депо кальция) происходит дефосфорилирование лёгкой цепи миозина при помощи фосфатазы лёгких цепей миозина. Дефосфорилированный миозин теряет сродство к актину, что предотвращает образование поперечных мостиков. Расслабление ГМК заканчивается разборкой миозиновых нитей. 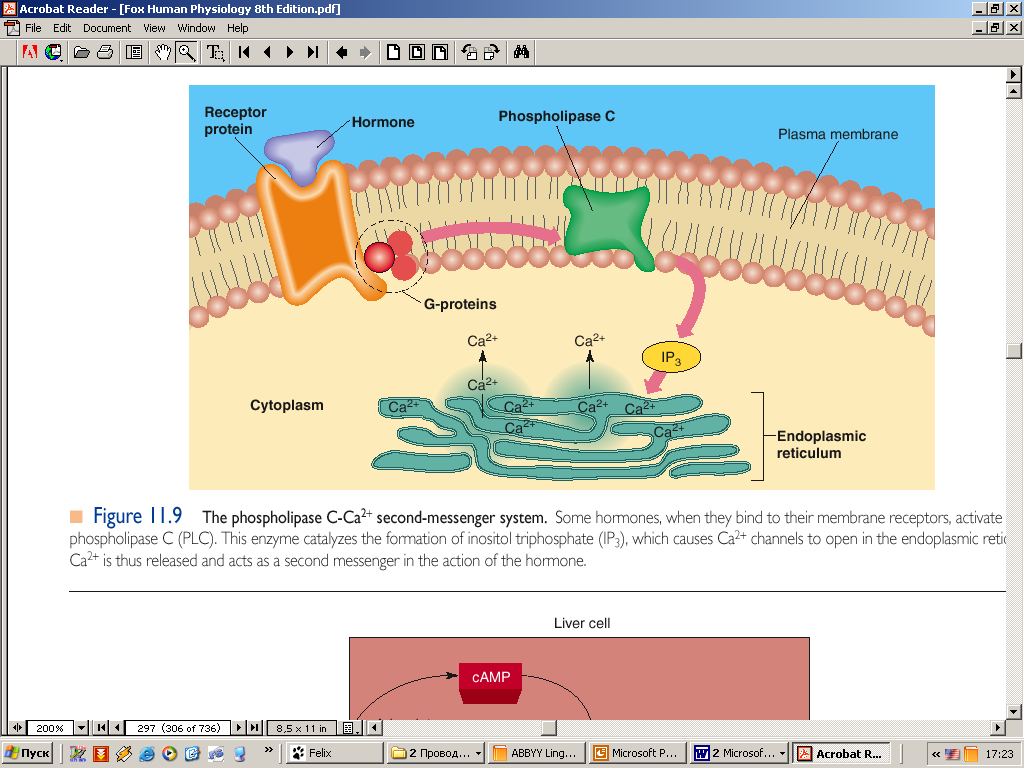 Стресс-релаксация (пластичность) гладкой мышцы. Важной особенностью висцеральной гладкой мышцы многих полых органов является ее способность изменять напряжение без закономерной связи с ее длиной. Например, внезапное увеличение объема жидкости в полости мочевого пузыря, сопровождается растяжением гладкой мышцы его стенки, что обязательно приводит к увеличению внутрипузырного давления. Однако в последующие от 15 сек до нескольких минут, несмотря на постоянно действующую растягивающую силу внутрипузырное давление возвращается к почти исходному значению и сокращение не наступает. Такую способность сохранения при растяжении длину, без изменения напряжения называют пластичностью ГМ. Рост гладкой мышцы, чувствительность к действию физиологически активных веществ В связи с тем, что пролиферация гладкой мышцы вносит большой вклад в развитие таких патологических процессов, как атеросклероз, гипертензия и утолщение сосудистой стенки, наблюдающееся при повреждении эндотелиоцитов, важно знать основные механизмы регуляции роста. Данный процесс является чрезвычайно сложным и включает в себя действие различных факторов роста. Катехоламины, ангиотензин-II активирует рост и пролиферацию гладкой мышцы. Глюкокортикоиды ингибируют рост. Гладкие мышцы обладают высокой чувствительностью к различным физиологически активным веществам: адреналину, норадреналину, АХ, гистамину и др. находящимся в крови. Это обусловлено наличием специфических рецепторов мембраны ГМК. В отличии от них скелетные мышцы отвечают на действие веществ только через синапс. Действие биологически активных веществ на гладкие мышцы находящиеся в различных органах не однозначно. Так, ацетилхолин возбуждает гладкие мышцы, которые находятся во внутренних органах, но тормозит в сосудах; адреналин способен расслаблять небеременную матку, но вызывает сокращение беременной. |