1. Дыхание Дыхание
 Скачать 313.54 Kb. Скачать 313.54 Kb.
|

|
«Дыхание» 1.Дыхание… Дыхание - это совокупность процессов, благодаря которым организм потребляет кислород из окружающей среды и выделяет углекислый газ. Этапы дыхания: 1. Внешнее дыхание /вентиляция легких/ - обмен газов между атмосферным воздухом и альвеолярным, легочная вентиляция. 2. Диффузия газов в легких - обмен газов между альвеолярным воздухом и кровью в капиллярах легких. 3. Транспорт газов кровью - этот этап осуществляется за счет деятельности сердечно-сосудистой системы, в результате чего кислород доставляется к тканям, а углекислый газ - к легким. 4. Диффузия газов в тканях - обмен газов между кровью и тканями. 5. Тканевое дыхание - окислительно-восстановительные реакции, протекающие с потреблением кислорода и выделением углекислого газа. Первые 4 этапа изучает физиология, последний, 5-ый - биохимия. Обеспечение тканей О2 и удаление из организма СО2 зависит от четырех процессов: 1.Вентиляция легких 2.Диффузия газов в альвеолы и ткани из крови и в кровь. 3.Перфузия легких кровью /интенсивность кровотока в легких/. 4.Перфузия тканей кровью Внешнее дыхание В обеспечении вентиляции легких участвуют три анатомо-физиологических образования: 1) дыхательные пути, обладают небольшой растяжимостью и сжимаемостью, формируют поток воздуха, 2). легочная ткань, обладает высокой растяжимостью и эластичностью/ способность принимать исходное положение после прекращения деформирующей (растягивающей) силы, 3) грудная клетка, пассивная костно–хрящевая основа, ригидная к внешним воздействиям, объединенная в целое связками и дыхательными мышцами, снизу – подвижная диафрагма. Взаимодействие грудной клетки и легкихГрудная клетка и легкие разделены плевральной полостью, которая представляет собой герметичную щель, содержащую небольшое количество жидкости (5 мл). Объем грудной клетки больше, чем объем легких. Поэтому легкие все время растянуты. Степень растяжения легких определяется транспульмональным давлением /разница между давлением в легких (альвеолах) и плевральной полости. В области диафрагмы это давление обозначается как трансдиафрагмальное. При этом в легких постоянно действует сила, стягивающая их, которая получила название "эластической тяги легких". Она зависит не только от эластичности легких, но, в значительной степени, и от силы поверхностного натяжения слизи, покрывающей альвеолы. Жидкость покрывает огромную поверхность альвеол и тем самым стягивает их. Однако сила поверхностного натяжения альвеол уменьшается за счет вырабатываемого в легких вещества сурфактанта. Благодаря этому легкие становятся более растяжимыми. Эластичная тяга легких создает отрицательное давление в плевральной полости. При выдохе оно равно - 6 мм рт.ст. На вдохе при растяжении грудной клетки давление в плевральной полости становится еще более отрицательным - 10 мм рс.ст. Понятие о пневмотораксе. Попадание воздуха в плевральную полость извне /открытый пневмоторакс /или из полости легких/закрытый пневмоторакс/ уравновешивает давление в плевральной полости с атмосферным и легкое за счет эластической тяги спадается. У человека в связи с особенностями грудной полости происходит спадание одного легкого. Легкие - максимально приспособлены для газообмена. Наличие газообмена между легкими и кровью постоянно требует обновления воздуха в легких /альвеолярного воздуха/, т.к. газовый состав воздуха будет постоянно изменяться в сторону снижения концентрации О2 и накопления СО2. Вентиляция легких, т.е. обмен газов между внешней средой и альвеолярным воздухом обеспечивается за счет вдоха /инспирация/ и выдоха /экспирация/, которые характеризуютсяглубиной вдоха и выдоха и частотой дыхания. Выделяют два вида дыхательных движений - спокойный вдох и выдох и форсированный вдох и выдох. Для нормального газообмена в атмосфере с обычным газовым составом здоровому взрослому человеку в спокойном состоянии необходимо 14-18 дыхательных движений в минуту, при длительности вдоха 2 с., объемной скорости вдоха 250 мл/с. При вдохе преодолевается ряд сил:1) эластическое сопротивление грудной клетки,2)эластическое сопротивление внутренних органов, оказывающих давление на диафрагму,3) эластическое сопротивление легких,4) вязко-динамическое сопротивление всех перечисленных выше тканей,5) аэродинамическое сопротивление дыхательных путей,6) силу тяжести грудной клетки,7) силы инерции перемещаемых масс/органов/Воздухоносные пути Верхняя часть воздухоносных путей представлена полостью носа и носоглотки. В легких воздухоносные пути (ВП) рассматриваются как ряд дихотомических трубок. В легком человека насчитывают 23 генерации бронхиального дерева. Первые 16 относятся к проводящей зоне трахеобронхиального дерева, 7 - транзиторной и респираторной зоне. Общая площадь поперечного сечения воздухоносных путей постепенно увеличивается с 2,5 см2 в трахее (0 генерация), на уровне 16 генерации (терминальные бронхиолы) - 180 см", на уровне 18 генерации - около 1000 см2 и далее - более 10 000 см2. Объем до 16 генерации включительно (анатомическое мертвое пространство, не принимает участия в газообмене) - 150 мл. Общий объем 23 генераций + 0 генерации (трахея) составляет 5700 мл (общая емкость легких). Функции воздухоносных путей (полости носа, носоглотки, респираторной зоны трахеобронхиального дерева) 1.Кондиционирование воздуха. 2.Проведение потока воздуха. 3.Иммунная защита. Кондиционирование воздуха Полость носа и носоглотки Посторонние частицы (более 10-15 мкм) задерживаются волосами преддверия носа и слизистой носовых ходов и носоглотки. Здесь происходит эффективное согревание воздуха за счет хорошего кровоснабжения слизистых оболочек, а так же увлажнение воздуха. Трахея, бронхи Происходит дальнейшее увлажнение воздуха. На слизистой этих образований осаждаются частицы менее 10 мкм, которые со слизью перемещаются в сторону входных / выходных ворот дыхательной системы. Осаждение частиц происходит за счет слизи, которая в виде пленки (толщина 5-10 мкм) располагается на слизистой островками, имеет свойства геля, секретируется преимущественно бокаловидными клетками, за сутки - 100 мл: 90 мл абсорбируется эпителиальными клетками, 10 мл передвигается по поверхности эпителия в глотку -проглатывается или откашливается (мокрота). В мокроте наряду с чужеродными частицами выделяются погибшие клетки слизистой, микроорганизмы. Секреция слизи находится под холин- и адренергическим контролем: ацетилхолин стимулирует секрецию слизи, а катехоламины ее тормозят. Гистамин, лейкотриены С4, D4> E4 стимулируют отделение слизи. Трахея и бронхи имеют механизм самоочищения - мукоцилиарный транспорт. Он обеспечивается мерцательными ресничками, которые скоординированно, однонаправленно (по направлению к входным / выходным воротам дыхательной системы) с частотой 900-1200 колебаний в минуту перемещают слизь со скоростью 5-20 мм/мин. Бронхиолы и альвеолы В них отсутствует система мукоцилиарного транспорта. Однако очищение потока осуществляется альвеолярными макрофагами, клетками Клара, сурфактантом. Альвеолярные макрофаги Альвеолярные макрофаги расположены на поверхности альвеол. Они фагоцитируют погибшие клетки, микроорганизмы, мелкие пылевые частицы, выделяют а-антитрипсин, который предохраняет альвеолы от действия протеаз. Альвеолярные макрофаги способны мигрировать вверх по ВП. Клетки Клара Участвуют в инактивации токсинов за счет цитохрома Р450, в образовании сурфактанта. Сурфактант предотвращает контакт эндотелия альвеол с инородными частицами, микробами. обволакиваемые сурфактантом чужеродные частицы фагоцитируются альвеолярными макрофагами и транспортируются в вышележащие отделы ВП опсонизирует микробные антигены и тем самым ускоряет их фагоцитоз альвеолярными макрофагами. 2. Биомеханика спокойного вдоха и выдоха… Биомеханика спокойного вдохаВ развитии спокойного вдоха играют роль: сокращение диафрагмы и сокращение наружных косых межреберных и межхрящевых мышц. Под влиянием нервного сигнала диафрагма /наиболее сильная мышца вдоха/ сокращается, ее мышцы расположены радиально по отношению к сухожильному центру, поэтому купол диафрагмы уплощается на 1,5-2,0 см, при глубоком дыхании -на 10 см, растет давление в брюшной полости. Размер грудной клетки увеличивается в вертикальном размере. Под влиянием нервного сигнала сокращаются наружные косые межреберные и межхрящевые мышцы. У мышечного волокна место прикрепления его к нижележащему ребру дальше от позвоночника, чем место его прикрепления к вышележащему ребру, поэтому момент силы нижележащего ребра при сокращении этой мышцы всегда больше, чем таковой у вышележащего ребра. Это приводит к тому, что ребра как бы приподнимаются, а грудные хрящевые концы как бы слегка скручиваются. Так как при выдохе грудные концы ребер располагаются ниже, чем позвоночные /дуга под углом/, то сокращение наружных межреберных мышц приводит их в более горизонтальное положение, окружность грудной клетки увеличивается, грудина приподнимается и выходит вперед, межреберное расстояние увеличивается. Грудная клетка не только приподнимается, но и увеличивает свои саггитальный и фронтальный размеры. За счет сокращения диафрагмы, наружных косых межреберных и межхрящевых мышц увеличивается объем грудной клетки. Движение диафрагмы обуславливает примерно 70-80% вентиляции легких. Грудная клетка выстлана изнутри париетальным листком плевры, с которым крепко сращена. Легкое покрыто висцеральным листком плевры, с которым также крепко сращено. В нормальных условиях листки плевры плотно прилегают друг к другу и могут скользить /благодаря выделению слизи/ относительно друг друга. Силы сцепления между ними велики и листки плевры невозможно разъединить. При вдохе париетальный листок плевры следует за расширяющейся грудной клеткой, тянет за собой висцеральный листок и тот растягивает ткань легкого, что приводит к увеличению их объема. В этих условиях воздух, находящийся в легких /альвеолах/ распределяется в новом, большем объеме, это приводит к падению давления в легких. Возникает разница давлений между окружающей средой и легкими /трансреспираторное давление/. Трансреспираторное давление(Ртрр) - это разница между давлением в альвеолах (Ральв) и внешним /атмосферным/ давлением (Рвнеш). Ртрр= Ральв. - Рвнешн,. Равняется на вдохе - 4 мм рт. ст. Эта разница и заставляет войти порцию воздуха через воздухоносные пути в легкие. Это и есть вдох. Биомеханика спокойного выдохаСпокойных выдох осуществляется пассивно, т.е. не происходит сокращения мышц, а грудная клетка спадается за счет сил, которые возникли при вдохе. Причины, вызывающие выдох: 1. Тяжесть грудной клетки. Поднятые ребра опускаются под действием тяжести. 2. Органы брюшной полости, оттесненные диафрагмой вниз при вдохе, поднимают диафрагму. 3. Эластичность грудной клетки и легких. За счет них грудная клетка и легкие занимают исходное положение Трансреспираторное давление в конце выдоха составляет =+ 4 мм.рт.ст. Биомеханика форсированного вдохаФорсированный вдох осуществляется за счет участия дополнительных мышц. Кроме диафрагмы и наружных косых межреберных мышц в нем участвуют мышцы шеи, мышцы позвоночника, лопаточные мышцы, зубчатые мышцы. Биомеханика форсированного выдохаФорсированный выдох активен. Он осуществляется за счет сокращения мышц - внутренних косых межреберных мышц, мышц брюшного пресса. 3. Клинико-физиологическая оценка внешнего дыхания. Легочные объемы… Анатомо-физиолгические показатели - легочные объемы определяются антропометрическими данными индивидуума : 1)росто-весовыми показателями, 2) строением грудной клетки, 3) дыхательных путей, 4) строением и свойствами легочной ткани (эластическая тяга легких, поверхностное натяжение альвеол), 5) силой дыхательных мышц Легочные объёмы и ёмкости
Легочные объемы: Общая емкость легких (ОЕЛ) - количество воздуха, находящееся в легких после максимального вдоха. ОЕЛ колеблется в больших пределах (от 0,5 до 8 литров) и зависит от роста, возраста, пола, состояния легких и грудной клетки. ОЕЛ состоит из 2 частей: жизненной емкости легких (ЖЕЛ) - объема, который человек может максимально выдохнуть после глубокого вдоха (в норме ЖЕЛ=Д(олжная)ЖЕЛ±10%), и остаточного объема (ОО) - объема воздуха, который остается в дыхательной системе даже после максимального выдоха (N=1-1,2 л). Увеличение ОО снижает эффективность дыхания. Делится на коллапсный объем /выходит при спадании легкого/ и минимальный объем /истинный остаточный/. Увеличение ЖЕЛ свидетельствует о повышении функциональных возможностей дыхательного аппарата. ЖЕЛ подразделяют на 3 составные части: 1. Дыхательный объем (ДО) - это объем воздуха, который человек вдыхает и выдыхает при каждом дыхательном цикле. В покое он составляет в среднем 20% от ЖЕЛ (0,3-0,6 л). 2. Резервный объем вдоха (РОвд) - воздух, который пациент может дополнительно вдохнуть, после спокойного вдоха /40% от ЖЕЛ/ (1,5-2,5 л). 3. Резервный объем выдоха (РОвд) - воздух, который пациент может максимально выдохнуть после спокойного выдоха /40% от ЖЕЛ/ (1,5-2,5 л). Соотношение составных частей ЖЕЛ очень изменчиво. При физической нагрузке ДО может увеличиться до 80%, что сопровождается уменьшением РОвд и РОвыд до 10 %. ДО является показателем глубины дыхания. Сумма ДО и РОвд получила название емкость вдоха(ЕВ) Сумма ОО и РОвыд получила название функциональной остаточной емкости (ФОЕ; объем воздуха, оставшийся после спокойного выдоха; N=2,5-3,5). Величина ФОЕ отражает эффективность дыхания. Все легочные объемы имеют должные величины, которые представлены в специальных таблицах. В таблицах отражена зависимость показателей не только от антропометрических параметров, но и от пола и возраста. Должные показатели приняты за индивидуальную норму. Объем мертвого пространства (ОМП) - это воздух, находящийся в носоглотке, трахее и бронхах и не участвующий в газообмене. Это анатомическое мертвое пространство. Этот объем не доходит до альвеол и не обменивается кислородом с кровью. ОМП у взрослого составляет в среднем 140-150 мл. Чем больше этот объем, тем менее эффективно дыхание. Есть понятие физиологического мертвого пространства - к нему относятся не только воздухоносные пути, но и альвеолы, которые не кровоснабжаются /альвеолярное мертвое пространство/. Коэффициент альвеолярной вентиляции (КАВ) указывает на то, какая часть воздуха обменивается при одном дыхании: КАВ=(ДО-ОМП) / ФОЕ В спокойном состоянии КАВ равен 1/7, то есть в альвеолах седьмая часть воздуха обменивается на атмосферный. Методы измерения легочных объемов1. Спирометрия - измерение легочных объемов. Позволяет определить ЖЕЛ, ДО, РОвд, РОвыд. 2. Спирография - регистрация легочных объемов. Позволяет документально зарегистрировать ЖЕЛ, ДО, РОвд, РОвыд, а также частоту дыхания. 3. Определение остаточного объема - с помощью спирографа с замкнутым контуром с использованием гелия /по степени разведения гелия/. - Общая плетизмография тела /бодиплетизмография/. Сложная дорогостоящая методика, выполняемая в специальной герметичной камере. Позволяет с высокой точностью определять общую емкость легких /ОЕЛ/ и остаточный объем /ОО/. Вышеуказанные показатели характеризуют не столько саму функцию дыхания, сколько потенциальную способность к выполнению этой функции. 4.Клинико-физиологическая оценка внешнего дыхания. Функциональные показатели... Физиологические показатели являются динамическими, т.к. характеризуют саму функцию внешнего дыхания во времени. 1. Минутный объем дыхания (МОД) - объем воздуха, который проходит через легкие за 1 минуту. Этот показатель можно определить двумя методами: с помощью спирографии (ДО умножается на частоту дыхания) и путем сбора воздуха в мешок Дугласа. В покое МОД составляет 4-6 литров в минуту. При физиологической нагрузке учащение и углубление дыхания приводят к возрастанию МОД до 30 л/мин. (4-11 л) 2. Максимальная вентиляция легких (МВЛ). МВЛ - это максимальное количество воздуха, которое может вдохнуть и выдохнуть пациент за 1 минуту (ЧД – более 50 уд/мин; N=14-18). В норме человек должен за минуту максимально провентилировать объем, равный ЖЕЛ * 40. (МВЛ=ДЖЕЛ*25 ±10%) 3. Форсированная жизненная емкость легких (ФЖЕЛ) - количество воздуха, которое пациент может выдохнуть за счет экспираторного маневра /максимально быстро и полно/. Характеризуется объемом форсированного выдоха за 1 секунду /ОФВ1сек/ (Форсированный экспираторный поток за 1 сек - дословный перевод термина с англ.).– Нормируется как ФЖЕЛ/ЖЕЛ, это индекс Тифно. В норме он составляет не менее 80% ЖЕЛ. Его снижение указывает на нарушение проходимости бронхиального дерева. Основные показатели, регистрируемые при выполнении ФЖЕЛ -Пиковая экспираторная объемная скорость /ПОС/-максимальный показатель объемной скорости потока (л/сек) при выполнении ФЖЕЛ. Характеризует силу дыхательных мышц и калибр «главных» бронхов. -Максимальная объемная скорость потока на уровне 25%, 50%, 75% от ФЖЕЛ. /МОС25%, МОС50%, МОС75%/. Определяется мгновенная скорость в данный момент форсированного маневра. Показатель характеризует уровень обструкции, т.е. уровень нарушения проходимости в бронхиальном дереве. МОС25% характеризует проходимость на уровне крупных бронхов, МОС50%- на уровне средних бронхов, МОС75%- на уровне мелких бронхов. Для ПОС и МОС существуют должные величины, с которыми проводится сопоставление полученных результатов. Vвд, Vвыд – максимальная скорость вдоха (выдоха) – определяется методом пневмотахометрии: Vвыд в норме 5-8 л/сек для мужчин, 4-6 л/сек для женщин; Vвд в норме не менее 90% от Vвыд. РД – резервы дыхания – резервные возможности дыхательной системы, которые могут быть мобилизованы при переходе от спокойного к форсированному дыханию. РД=МВЛ-МОД. (N=60% от МВЛ) Коэффициент альвеолярной вентиляции (КАВ) указывает на то, какая часть воздуха обменивается при одном дыхании: КАВ=(ДО-ОМП) / ФОЕ В спокойном состоянии КАВ равен 1/7, то есть в альвеолах седьмая часть воздуха обменивается на атмосферный. КИК – коэффициент использования кислорода – характеризует количество кислорода, потребляемого из вдыхаемого воздуха за одну минуту (ПО2) N=40 ±10% КИК=ПО2 (мл) / МОД (л) Показатели объемной скорости нельзя получить при спирографии, для этого используется пневмотахография Пневмотахография проводится с помощью приборов пневмотахометров, снабженных специальными датчиками - термоанемометрами, при прохождении струи выдыхаемого воздуха меняется электрическое сопротивление пропорционально объемной скорости воздушного потока, что позволяет по показаниям прибора вычислить основные параметры внешнего дыхания. Компьютерный анализ позволяет представить полученную информацию в виде кривой «поток-объем», которая отражает проходимость различных участков дыхательных путей. 5.Газообмен в легких и тканях… В процессе внешнего дыхания происходит газообмен в легких. За счет этого формируется состав альвеолярного и выдыхаемого воздуха. Газовый состав вдыхаемого, альвеолярного и выдыхаемого воздуха
Внешнее дыхание необходимо для обновления альвеолярного воздуха, т.к. в процессе жизнедеятельности идет постоянный процесс потребления О2 и выделения СО2, это поддерживает концентрацию дыхательных газов в нем на постоянном уровне. Интенсивность внешнего дыхания подчинена задачам обеспечения оптимальных условий для газообмена в организме. Оптимальные условия сохраняются в организме определенное время (3-4 секунды). Этим и определяется частота дыхания (14-18 в минуту). Таким образом, аппарат дыхания обладает резервами, которые позволяют обменивать воздух с определенной периодичностью. Процесс газообмена состоит из 3-х этапов дыхания: 2 этапа дыхания. Обмен газов между альвеолярным воздухом и кровью. 3 этапа дыхания. Транспорт газов кровью. 4 этапа дыхания. Обмен газов между кровью и тканями. В основе 2 и 4 этапов дыхания лежат одни и те же механизмы, т.е. в основе обмена газов между альвеолами и кровью, а также кровью и тканями лежит одно физическое явление - процесс диффузии. Механизмы 2-го и 4-го этапов дыхания. Мембраны клеток хорошо проницаемы для газов, следовательно для перемещения газов из одной среды в другую не надо активного транспорта, а достаточно физического процесса диффузии. В основе диффузии лежит разность концентраций. Молекулы из области большей концентрации распространяются в область меньшей концентрации. Если газ находится над жидкостью, он также легко в неё переходит, растворяясь в ней. Интенсивность перехода газов в жидкость зависит от парциального давления газа над ней. Давление газа в смеси с другими газами, выраженное в мм рт. ст., принято называть "парциальным давлением". Давление газа, растворенного в жидкости, обозначают как "напряжение". При относительно длительном контакте газов и жидкости в определенный момент времени парциальное давление газа над жидкостью и напряжение газа в жидкости выровняются. При резком снижении парциального давления одного из газов либо снижении суммарного атмосферного давления жидкость с растворенными в ней газами начинает "кипеть" (до тех пор, пока вновь не выровняются парциальное давление и напряжение газов (примеры с шампанским, "кессонная болезнь" - помощь - экстренное помещение в барокамеру с постепенным снижением давления)). Содержание дыхательных газов в альвеолярном воздухе, крови и тканях
Примечание: стрелочкой указано направление диффузии. При этом следует иметь в виду, что аэрогематический барьер легких обладает определенной проницаемостью, которая характеризуется диффузионной способностью легких. Диффузионная способность легких - это количество мл газа которое проходит за 1 минуту через легочную мембрану при разнице парциальных давлений по обе стороны мембраны 1 мм.рт.ст. Для О2 составляет 20-25 мл, для СО2 она существенно больше/т.к. разница парциального давления меньше многократно/, а объем выделяемого СО2 такой же как и О2. С возрастом диффузионная способность легких снижается. Гистагематический 6.Транспорт газов кровью… Механизмы связывания газов кровью 1. Физическое растворение 2. Химическое связывание 1. Физическое растворение. В жидкой части крови растворены газы воздуха: кислород, углекислый газ, азот. Растворение О2 и СО2 в воде не играет физиологической роли. 2. Химическое связывание кислорода кровью. Насыщение кровью кислородом зависит от: Альвеолярной вентиляции /pO2 в альвеолах/ Кровотока в легких Диффузионной способности легких Содержания гемоглобина в эритроцитах 1 г HHb способен связать 1,35 мл О2. При содержании гемоглобина 150 г/л (норма) каждые 100 мл крови переносят 20,8 мл О2. Это кислородная емкость крови. Другой показатель-содержание кислорода в крови, взятой в различных участках сосудистого русла: артериальной/20 мл О2/100 мл крови/ и венозной/14 млО2/100 мл крови/. Следующий показатель - артерио-венозная разница/норма 5-6 мл О2/100 мл крови/. Отношение кислорода, связанного с гемоглобином к кислородной емкости крови/все выраженное на 100 мл крови/ называется насыщение гемоглобина кислородом. В артериальной крови оно составляет в норме 96%. Гемоглобин присоединяет кислород с помощью непрочной водородной связи, с образованием оксигемоглобина Эта реакция обратима: Нв+О2=НвО2 Направленность реакции зависит от содержания кислорода: если количество кислорода в крови увеличивается, то реакция идет в сторону образования оксигемоглобина, если уменьшается - то в противоположную сторону. 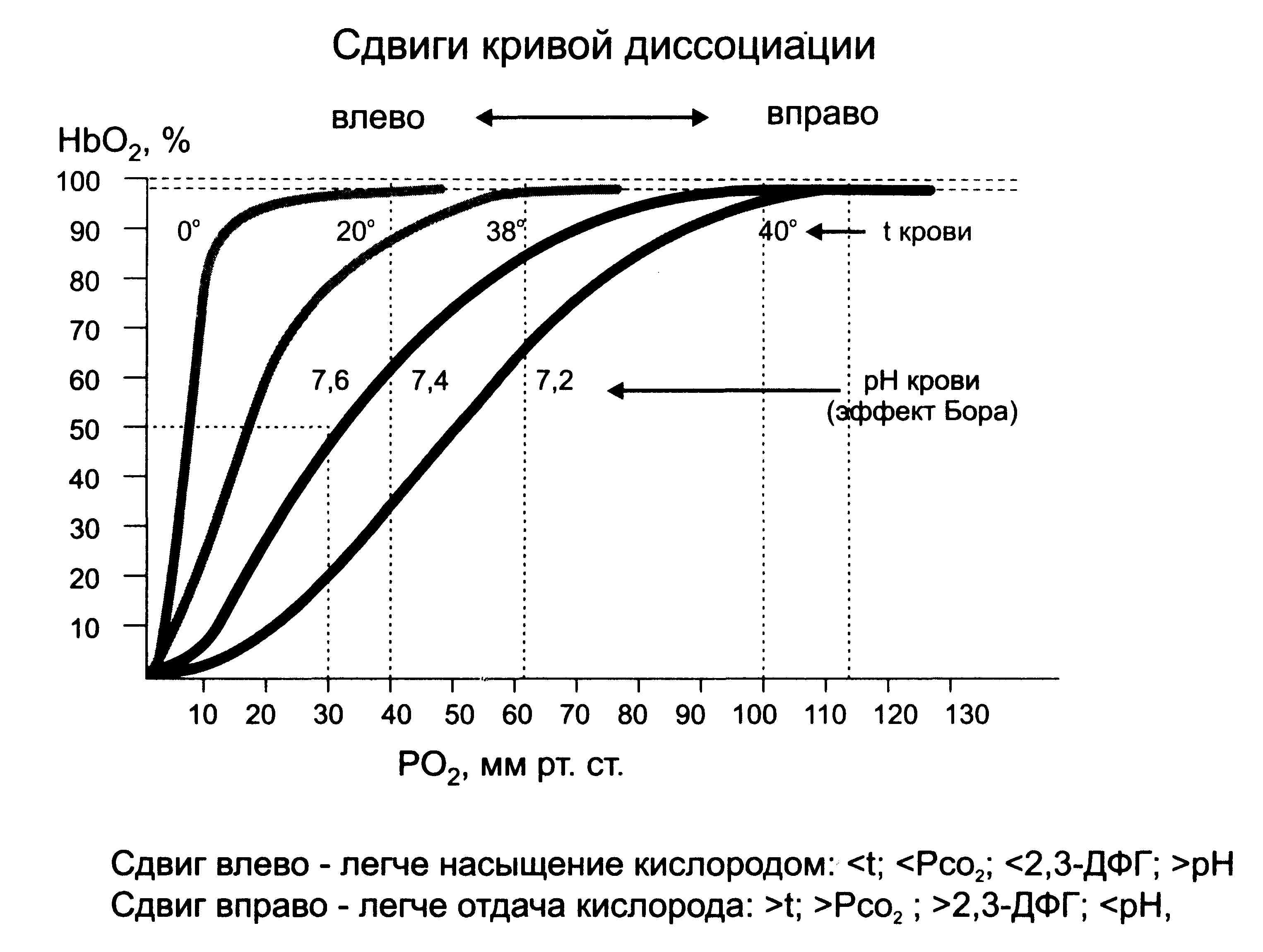 Динамика взаимодействия Нв и О2 отражается кривой диссоциации оксигемоглобина. Эта кривая количественно определяет приведенную выше реакцию связывания гемоглобином кислорода. Кривая отражает общую закономерность: увеличение количества кислорода сопровождается усиленным образованием оксигемоглобина. Кривая диссоциации оксигемоглобина имеет S-образный вид. Это связанно с тем, что до 10 мм рт. ст. кислород связывается гемоглобином медленно, затем до 60-50 мм рт. ст. скорость реакции резко увеличивается, кривая круто поднимается вверх, при давлении 90 мм рт. ст., когда более 98% гемоглобина связано с кислородом, она вновь идет почти горизонтально. Избыток СО2 и ацидоз сдвигает кривую диссоциации вправо, а недостаток СО2 и алкалоз – влево(эффект Бора). В легких реакция взаимодействия гемоглобина с кислородом идет в сторону образования оксигемоглобина, т.к. венозная кровь имеет напряжение кислорода 40 мм рт. ст., а в альвеолярном воздухе парциальное давление кислорода составляет 100 мм рт. ст. В тканях напряжение О2 равно 20-40 мм рт. ст., а в артериальной крови - 100 мм рт. ст., в связи с этим реакция идет в сторону распада оксигемоглобина. Кровь отдает ткани часть О2.. Этот процесс оценивается коэффициентом утилизацией/ кислорода(КУК). КУК это отношение потребленного кислорода к кислородной емкости крови. В норме в покое 30-40%, при физ. нагрузках существенно возрастает. Для оценки эффективности газообмена вычисляют коэффициент использования кислорода (КИК). Он показывает количество кислорода в мл, которое потребляется из 1 литра воздуха. В норме он составляет 40 мл. Химическое присоединение СО2 Напряжение СО2 в тканях составляет 60 мм.рт.ст., а в притекающей крови 50-60 мм.ст.рт. Благодаря этому СО2 переходит из ткани в кровь/46 мм.рт.ст./. Основная форма связывания СО2 кровью - это образование бикарбонатов натрия и калия. СО2 + Н2О = Н2СО3 Эта реакция обратима, ее направление зависит от количества СО2. Его увеличение сдвигает реакцию вправо, уменьшение - влево. Образующаяся угольная кислота диссоциирует: Н2 СО3 ---- Н+ + НСО3- Следовательно, в эритроците образуются катионы Н+ и анионы НСО3-.катионы водорода вступают в реакцию восстановления гемоглобина: Н+ + Нв ННв, Анионы НСО3- - частично выходят из эритроцитов в плазму из-за разности концентраций. Таким образом, в плазме и в эритроцитах появляется значительное количество анионов НСО3 - , которые в плазме взаимодействуют с катионами натрия/55%/, а в эритроцитах – калия/35%/, образуя гидрокарбонаты Na и К. Ключом всех этих реакций служит фермент карбоангидраза, который содержится в мембранах эритроцитов и катализирует обратимую реакцию соединения углекислого газа с водой. Кроме того, небольшое количество углекислого газа /10%/ транспортируется в виде карбогемоглобина - соединения СО2 с гемоглобином. 7.Регуляция дыхания… Главная задача регуляции дыхания - чтобы потребление кислорода, поставка его тканям за счет внешнего дыхания были адекватны функциональным потребностям организма. Самый эффективный способ регуляции дыхания в целом - это регуляция внешнего дыхания. Интенсивность внешнего дыхания зависит от варьирования его частоты и глубины. При этом изменяется доставка кислорода организму и выведение из него углекислого газа. В регуляции дыхания можно выделить 3 группы механизмов: 1. Обеспечение организации дыхательного акта (последовательность вдоха и выдоха). 2. Перестройка дыхания в соответствии с потребностями организма - изменение частоты и глубины дыхания. 3. Регуляция тонуса кровеносных сосудов легких и бронхиального дерева. 1-ая группа. Механизмы организации дыхательного акта Чередование вдоха и выдоха организуется благодаря деятельности дыхательного центра. Отличия морфологического и функционального понятия НЦ. Дыхательный центр представляет собой совокупность нейронов, объединенных общей функцией организации и регуляции дыхания и расположенных в разных "этажах" центральной нервной системы. Выделяют 4 "этажа" : - спинной мозг, - продолговатый мозг, - варолиев мост, - высшие отделы ЦНС (гипоталамус, лимбическая система, кора больших полушарий). Каждый из перечисленных отделов имеет определенную функцию. 1 этаж: Спинной мозг содержит двигательные центры дыхательной мускулатуры. Представлены мотонейронами передних рогов спинного мозга: - грудной отдел (Th1 - Th6 - nn. intercostales) - межреберные нервы иннервируют наружные косые межреберные мышцы. - шейный отдел (С3 - С5) - n. frenicus. Диафрагмальный нерв иннервирует диафрагму. При перерезке ЦНС между спинным и продолговатым мозгом процесс дыхания прекращается (т.к. центры спинного мозга не обладают автоматией). При перерезке спинного мозга между шейным и грудным отделами дыхание сохраняется за счет сокращения диафрагмы (диафрагмальное дыхание). 2 этаж: Дыхательный центр продолговатого мозга (собственно дыхательный центр) обеспечивает последовательную смену вдоха и выдоха. Открыт в 1885 г. русским исследователем Н.А. Миславским на дне 4-го желудочка продолговатого мозга. Это - парное образование. Связан проводящими путями с выше- и нижерасположенными нервными центрами (мотонейроны спинного мозга - 1-ый этаж дыхательного центра). В составе дыхательного центра часть нейронов ответственна за вдох, другая часть - за выдох. Т. е. Выделяют т.н. Экспираторный и Инспираторный центры. Это - функциональные образования, т.к. морфологически их выделить нельзя. Между центрами - реципрокные взаимоотношения. Это и обеспечивает чередование процессов вдоха и выдоха, т.к. активация нейронов одного отдела вызывает угнетение другого. Собственно дыхательный центр обладает автоматией. 4-5 раз в минуту в ДЦ возникает самопроизвольное возбуждение, не связанное с поступлением импульсов из других центров, а обусловленное особенностью метаболизма клеток ДЦ. Это обеспечивает автономность от других влияний и поддержание жизненно важной функции на базальном уровне. Таким образом, при пересечении ЦНС выше продолговатого мозга будет наблюдаться глубокое и редкое дыхание (дыхание Куссмауля), другие виды патологического дыхания: Чейн-Стокса, Биотта, Грокко. Третий "этаж" дыхательного центра расположен в варолиевом мосту и назван пневмотАксическим(таксис). Он способствует переключению возбуждения с центра вдоха на центр выдоха и наоборот. Возбуждение пневмотаксического центра приводит к угнетению центра вдоха, а нейроны, ответственные за выдох - активируются. Существует и обратный механизм, который обеспечивает переключение с выдоха на вдох. Перерезка ЦНС выше Варолиева моста позволяет поддерживать частоту дыхания на уровне 14-18 в минуту. Роль периферических процессов в функционировании 2 и 3 этажа дыхательного центра, в организации дыхательного акта. Четвертый этаж - высшие отделы ЦНС. Гипоталамус - регулирует дыхание во время простых поведенческих актов: - при общей защитной реакции организма (боль, физическая работа); - высший центр терморегуляции, поэтому при гипертермии наблюдается учащение дыхания без изменения его глубины (значительно увеличивается вентиляция ОМП, что увеличивает теплоотдачу: дыхание собаки в жару). Лимбическая система - регуляция дыхания при эмоциях ("хмыкнул" - разная интонация м.б., "чего сопишь ?", крайние формы выражения эмоций - смех и плач - это измененные дыхательные движения). Кора больших полушарий принимает участие: - в выработке условных дыхательных рефлексов, - в приспособлении дыхания к изменяющимся условиям окружающей среды (глотание, пение, речь, ныряние, произвольное апное и гиперпное). Дыхание - единственная функция внутренних органов, подверженная сознательной регуляции без предварительной тренировки (йоги), так как висцеральная функция реализуется через соматическую мускулатуру. 8.Механизмы перестройки внешнего дыхания… 1. Роль хеморецепторов 1.1. Влияние углекислого газа (СО2) Мощным регулятором дыхания является СО2. Пути воздействия СО2: - Рефлекторно (через хеморецепторы сосудистого русла, прежде всего –каротидного синуса). - Прямое воздействие СО2 на нейроны ствола мозга, собственно ДЦ. Накопление СО2 в крови /гиперкапния/ стимулирует дыхание - человек будет дышать глубже и чаще. С другой стороны, после искусственного гиперпное возникает апное (т.к. СО2 вымывается из крови /гипокапния/). 1.2. Значение О2. Хеморецепторы каротидного синуса реагируют на изменение рО2, но менее выраженно, чем на СО2, гипоксия стимулирует дыхание, гипероксия снижает объем легочной вентиляции. 1.3. Влияние ацидоза и алкалоза Ацидоз стимулирует дыхание, алкалоз тормозит 2.Роль механо- и барорецепторов 2.1. Влияние на рецепторы растяжения в легких. Это рефлекс Геринга –Брейера, его открытию предшествовала серия опытов: 1. Двусторонняя перерезка блуждающих нервов сопровождалась регистрацией глубокого и редкого дыхания (4-5 в минуту). 2. Механическое растяжение легких приводит к стимуляции выдоха. Оказалось, что в деятельности дыхательного центра имеет значение информация с рецепторного поля легких, о чем свидетельствует, в частности, рефлекс Геринга-Брейера. Он заключается в следующем: при вдохе легочная ткань растягивается, это вызывает возбуждение альвеолярных рецепторов, передающееся по чувствительным волокнам, идущим в составе блуждающего нерва (не имеют отношения к парасимпатике) в пневмотаксический центр. При этом центр вдоха тормозится, вдох сменяется на выдох. Интенсивность импульсации пропорциональна степени растяжения. Т.о. рефлекс Геринга-Брейера - защитный рефлекс, предохраняющий легкие от перерастяжения. 2.2 Влияние на ирритантные рецепторы трахеи и бронхов. Это защитные дыхательные рефлексы - чихание и кашель. Чихание представляет собой глубокий вдох и быстрый выдох через носовую полость при воздействии на ее слизистую оболочку химических или слабых механических раздражителей. Кашель - при раздражении рецепторов слизистой гортани, глотки, трахеи и бронхов (глубокий вдох, закрытие голосовой щели, резкое её открытие и выдох). Нередко - бронхоконстрикция. 2.3 Влияние на j-рецепторы. В интерстиции альвеол и бронхиол находится этот вид рецепторов /н.окончаний/. Возбуждаются при повышении давления в малом круге кровообращения и увеличении объема интерстициальной жидкости в легких. Вызывают частое поверхностное дыхание, нередко бронхоконстрикцию. 2.4. Раздражение рецепторов скелетных мышц. Выполнение мышечной работы делает дыхание более глубоким и частым (рефлекторно с проприорецепторов, т.е. ещё до повышения уровня СО2 в крови). 2.5. Влияние на барорецепторы сосудистого русла - повышение гидростатического давления сопровождается снижением вентиляции. 2.6. Влияние на болевые рецепторы. Боль - стимулирует вентиляцию. 3. Температура тела - гипертермия и незначительная гипотермия стимулируют вентиляцию. 4. Гормоные воздействия - повышают вентиляцию, воздействуя непосредственно на Дыхательный Центр: - Адреналин (при физической и умственной работе), -Прогестерон - при беременности обеспечение кислородом организма матери и плода. 5. Участие коры головного мозга в регуляции дыхания. При этом дыхание заранее приспосабливается к определенной нагрузке. Происходит опережающее приспособление аппарата дыхания к будущим затратам (спортсмен на старте). 3-я группа. Механизмы регуляция тонуса сосудов и бронхиол. Регуляция тонуса сосудов легких 1) Ведущая роль принадлежит газовому составу крови: - понижение содержания в крови СО2 приводит к повышению тонуса легочных сосудов (при этом уменьшается количество крови, которое успевает обогатиться в легких О2 за единицу времени); - увеличение СО2, наоборот, уменьшает тонус легочных сосудов (а значит повышается кровоток и газообмен). 2) На тонус сосудов влияет газовый состав альвеолярного воздуха: При гипоксии и гиперкапнии альвеолярного воздуха повышается тонус сосудов легких, т.е. кровоток перераспределяется в пользу участков легочной ткани с лучшей оксигенацией. Регуляция просвета бронхиального дерева. Сужение бронхов вызывают парасимпатические нервы, а также гормоны воспаления - гистамин, ацетилхолин, серотонин. Расширение - симпатическая нервная система и адренорецепторы. Первый вдох. После рождения прекращается поступление кислорода из крови матери. Накопление углекислоты стимулирует дыхательный центр, в результате чего сокращаются дыхательные мышцы. У плода грудная клетка находится в спавшемся состоянии, т.к. головки ребер расположены вне своих суставных ямок. При первом вдохе ребра не просто поднимаются, а головки занимают свои суставные ямки, грудная клетка меняет форму, она резко увеличивается в размере, легкие остаются растянутыми. |