Лекция "Биологическое окисение"_2009. 2010г лекция 4 Тема Биологическое окисление этапы, цикл Кребса, пути использования кислорода в тканях
 Скачать 0.53 Mb. Скачать 0.53 Mb.
|

|
ГОУВПО УГМА Федерального агентства по здравоохранению и социальному развитию кафедра биохимии КУРС ЛЕКЦИЙ ПО ОБЩЕЙ БИОХИМИИ Модуль 2. Биологическое окисление Автор: к.б.н., доцент кафедры биохимии Гаврилов И.В. Екатеринбург, 2010г ЛЕКЦИЯ № 4 Тема: Биологическое окисление: этапы, цикл Кребса,пути использования кислорода в тканях.Факультеты: лечебно-профилактический, медико-профилактический, педиатрический. 2 курс. История развития учения о биологическом окислении Изучение окисления в организме было начато в 18 в. А. Лавуазье; в дальнейшем значительный вклад в исследование БО (его локализация в живых клетках, связь с др. процессами обмена веществ, механизмы ферментативных окислительно-восстановительных реакций, аккумуляция и превращение энергии и др.) внесли О. Варбург, Г. Виланд (Германия), Д. Кейлин, Х. Кребс, П. Митчелл (Великобритания), Д. Грин, А. Ленинджер, Б. Чанс, Э. Рэкер (США), а в СССР - А. Н. Бах, В. И. Палладин, В. А. Энгельгардт, С. Е. Северин, В. А. Белицер, В. П. Скулачев и др. В 1777г французский химик Антуан Лоран Лавуазье (1743-1794), впервые правильно истолковал явление горения как процесс соединения веществ с О2. Так как горение и дыхание сопровождаются потреблением О2 и выделение СО2, он предположил что, в их основе лежит один процесс. Однако у дыхания были существенные отличия от горения: оно шло при низкой температуре, без пламени и в присутствии воды. В конце XIX века русские исследователи А.Н. Бах и В.И. Палладин, работая независимо друг от друга, предложили 2 основные теории для объяснения процессов, протекающих в ходе биологического окисления. В 1897г Алексей Николаевич Бах (1857-1946) сформулировал «перекисную теорию окисления», согласно которой молекула О2 сначала активируется. Активация О2 происходит в результате разрыва одной его связи (-О-О-) и присоединения к органическим веществам – оксидазам. Активированный О2 при взаимодействии с окисляемым веществом образует перекись. 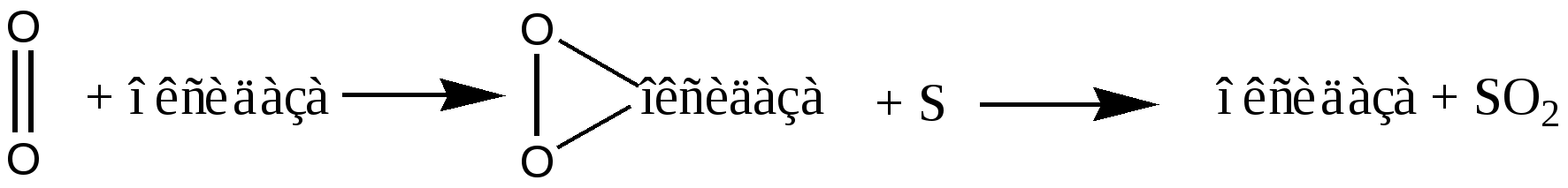 Теория «перекисного окисления» Баха нашла свое подтверждение, однако главный механизм БО оказался иным. Важнейшая заслуга в развитии учения о БО принадлежит русскому ученому ботанику и биохимику В.И. Палладину (1859-1922), который создал теорию дыхания, как совокупности ферментативных процессов. Он предположил, что окисление субстратов может происходить в 2 фазы: 1). Анаэробная фаза. В этой фазе особые вещества хромогены (R) отщепляют Н от субстратов и восстанавливаются (RH2). 2). Аэробная фаза. Восстановленные хромогены RH2 передают Н на О2. 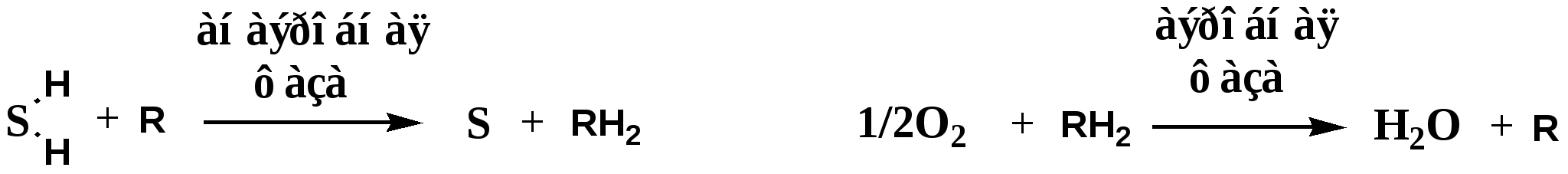 В последствии теория В.И. Паладина подтвердилась для процессов митоходриального окисления, а хромогены, принимающие участие в отнятии водорода от субстратов, в настоящее время называются ферментами дегидрогеназами. Современные представления о биологическом окислении Согласно современной теории БО:
Окислительно-восстановительные реакции (ОВР) – реакции, в которых меняется степень окисления субстрата за счет присоединения/отщепления: 1) 1 е-; 2) 2е- и 2Н+; 3) атомов кислорода. Биологическое окисление (БО) совокупность окислительно-восстановительных реакций, которые протекают во всех живых клетках. Субстрат БО – вещество, способное отдавать электрон. (Любые вещества, способные вступать в реакции окисления). Тканевое дыхание – окисление органических веществ в клетках, сопровождающееся потреблением О2 и выделением воды. Субстрат тканевого дыхания – это вещество, которое отдает электрон непосредственно в цепь окислительного фосфорилирования. (Например, НАДН2, ФАДН2). Дыхательная цепь – цепь переноса электронов. В переносе электронов от субстратов БО к О2 принимают участие: 1) НАД и НАДФ зависимые ДГ; 2) ФАД и ФМН зависимые ДГ; 3) цитохромы; 4) коэнзим Q; 5) белки, содержащие негеминовое железо. Свободная энергия. Каждое органическое вещество обладает определенным запасом внутренней энергии (Е). Часть этой внутренней энергии может быть использована для совершения полезной работы, такую энергию называют свободной (G). Направление химической реакции определяется значением ∆G. У катаболических реакций ∆G отрицательно, эти реакции протекают самопроизвольно (экзергонические реакции). У анаболических реакций ∆G положительно, они протекают только при поступлении свободной энергии извне (эндергонические реакции). Редокс-потенциалы. В каждой окислительно-восстановительной системе участвует окисленная и восстановленная формы одного соединения, которые образуют сопряженную окислительно-восстановительную или редокс-пару. Разные редокс-пары обладают различным сродством к электрону. Мерой сродства редокс-пары к электрону служит окислительно-восстановительный потенциал, или редокс-потенциал (Ео'), величина которого прямо пропорциональна изменению свободной энергии ∆G. Величину Ео' выражают в вольтах; чем она отрицательнее, тем меньше сродство вещества к электронам и наоборот. Самое низкое сродство к электрону -0,42В у водорода. Самое высокое сродство к электрону +0,82В у О2. Компоненты дыхательной цепи имеют редокс-потенциалы, занимающие промежуточное положение между -0,42В и +0,82В. В дыхательной цепи, вещества переносящие электрон, располагаются в порядке увеличения их редокс-потенциала. Электрон перемещается по дыхательной цепи от веществ с низким сродством к электрону к веществам с более высоким сродством к электрону, при этом происходит высвобождение свободной энергии, часть которой фиксируется в виде макроэргических соединений. Электроны в дыхательную цепь поставляют субстраты тканевого дыхания. Этапы унифицирования энергии пищевых веществ и образования субстратов тканевого дыхания Образование субстратов тканевого дыхания осуществляется в несколько этапов:
Пути использования кислорода в клетке В настоящее время выделено 4 основные пути использования кислорода в организме:
В соответствие с путями потребления кислорода и катализируемыми реакциями, все оксидоредуктазы разделены на 5 групп: 1). Оксидазы удаляют Н из субстрата (S), используя в качестве акцептора Н только О2, продуктом реакции является вода. Все оксидазы содержат медь. Например, цитохромоксидаза, последний фермент дыхательной цепи. SН2 + ½О2 → S + Н2О. 2). Аэробные дегидрогеназы, или ФАД зависимые оксидазы, относятся к металлофлавопротеинам (ФАД, ФМН, Fe, Cu, Mo), находятся в пероксисомах и наружной мембране митоходрий. Они отнимают Н от субстрата, и передают его на О2 с образованием перекиси. Дегидрогеназа L-аминокислот (оксидаза L-аминокислот) в почках осуществляет окислительное дезаминирование. Ксантиндегидрогеназа (ксантиноксидаза) превращает пурины в мочевую кислоту. Моноаминоксидазы (МАО) окисляют гормон адреналин и некоторые биогенные амины. Диаминоксидазы (ДАО) окисляют гистамин и другие диамины и полиамины. Образующаяся перекись бактерицидное действие. SН2 + О2 → S + Н2О2 3). Анаэробные дегидрогеназы многочисленная группа, содержат в качестве коферментов НАДН2, НАДФН2, ФАД, ФМН, цитохромы. Они удаляют Н из субстрата, не используя в качестве акцептора протона О2. Анаэробные дегидрогеназы выполняют две главные функции: перенос Н с одного субстрата на другой в сопряженной ОВР и транспорт электронов в дыхательной цепи. Примеры ферментов: изоцитрат ДГ (НАДН2), сукцинат ДГ (ФАДН2), цитохромы в, с1, с, а и а3. SН2 + R → S + RН2 4). Оксигеназы. Включают кислород в молекулу субстрата. Работают в составе мультиферментного комплекса, содержащего ФАД зависимую ДГ, Fe2S2-белок, цитохромы Р450 или В5. Этот комплекс встроен в мембрану ЭПР или внутреннюю мембрану митохондрий. а) Монооксигеназы. Обеспечивают включение 1 атома кислорода в молекулу субстрата. В качестве восстановителей (R) используется НАДФН2, НАДН2 и аскорбат. Участвуют в синтезе стероидных гормонов, обезвреживании ксенобиотиков. Фен-4-монооксигеназа окисляет фен в тир. Пролингидроксилаза (аскорбат) гидроксилирует пролин в проколлагене. SН + О2 + RH2 → SOH + Н2О + R б) Диоксигеназы. Обеспечивают включение молекулы кислорода в молекулу субстрата, окисляют циклические соединения с разрывом цикла. Например, триптофаноксигеназа, превращает триптофан в фенилкенуренин. S + О2 → SO2 5). Гидроксипероксидазы разрушаютперекиси водорода и органические перекиси. Ферментыпероксидаза и каталаза. Пероксидазы, содержат протогем, восстанавливают перекись до воды за счет хинонов, аскорбата или цитохрома с. Глутатионперосидаза, содержит селен, восстанавливает перекись до воды за счет глутатиона. Н2О2 + RH2 → 2Н2О + R Каталаза, гемопротеин, использует одну Н2О2 как донор е-, а вторую Н2О2 как акцептор е-, в результате образуется вода и молекула кислорода. Каталазы много в крови, костном мозге, слизистых оболочках, почках и печени. 2Н2О2 → 2Н2О + О2. Цикл Кребса ЦТК является процессом окисления АцетилКоА - универсального продукта катаболизма углеводов, жиров и белков. ЦТК протекает в митохондриях с участием 8 ферментов, которые локализованы в матриксе в свободном состоянии, или на внутренней поверхности внутренней мембраны. В ЦТК участвуют 5 витаминов В1, В2, РР, пантотеновая кислота и липоевая кислота в виде коферментов тиаминпирофосфата, ФАД, НАД+, КоА и липоата. Основной функции ЦТК является образование водородных эквивалентов, которые в цепи окислительного фосфорилирования обеспечивают синтез макроэргических соединений. Кроме того, ЦТК выполняет ведущую роль в процессах глюконеогенеза, переаминирования, дезаминирования АК, липогенеза и синтеза гема. Интегрирует все виды обмена веществ. Регуляция ЦТК. Осуществляется с участием 4 регуляторных ферментов: цитратсинтазы, изоцитрат ДГ, α-КГ ДГ и СДГ. ЦТК ингибируется в основном НАДН2 и АТФ, которые являются продуктами ЦТК и цепи окислительного фосфорилирования. Активируют ЦТК в основном НАД+ и АДФ. Реакции ЦТК 1). Цитратсинтаза локализуется в матриксе митохондрий, ее активируют ЩУК, НАД+; ингибируют АТФ, НАДН2, Сукцинил-КоА, цитрат.  2). Аконитаза локализуется в матриксе митохондрий. 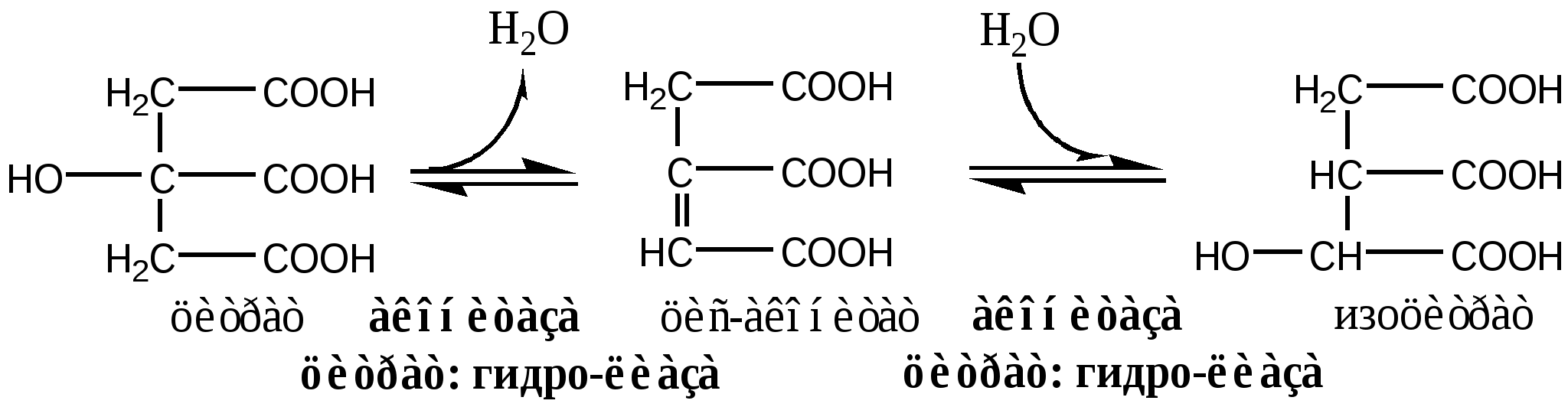 3). Окислительно-восстановительная реакция, самая медленная в ЦТК. Изоцитратдегидрогеназа локализуется в матриксе митохондрий, ее активируют АМФ, Са2+, АДФ, НАД+; ингибируют АТФ, НАДН2. 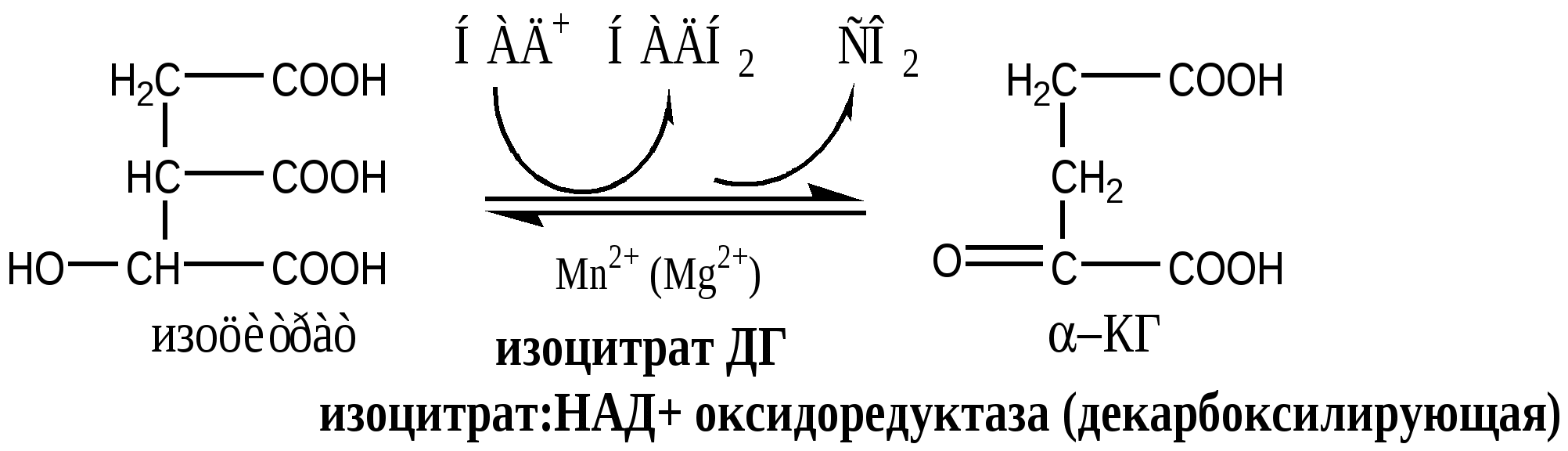 4). Окислительно-восстановительная реакция. α-кетоглутаратдегидрогеназный комплекс состоит из 3 ферментов и содержит 5 коферментов: тиаминдифосфат, кофермент А, липоевая кислота, НАД+, ФАД. α-КГ ДГ активируется Са2+, ингибируется сукцинил-КоА, АТФ, НАДН2.  5). Реакция субстратного фосфорилирования 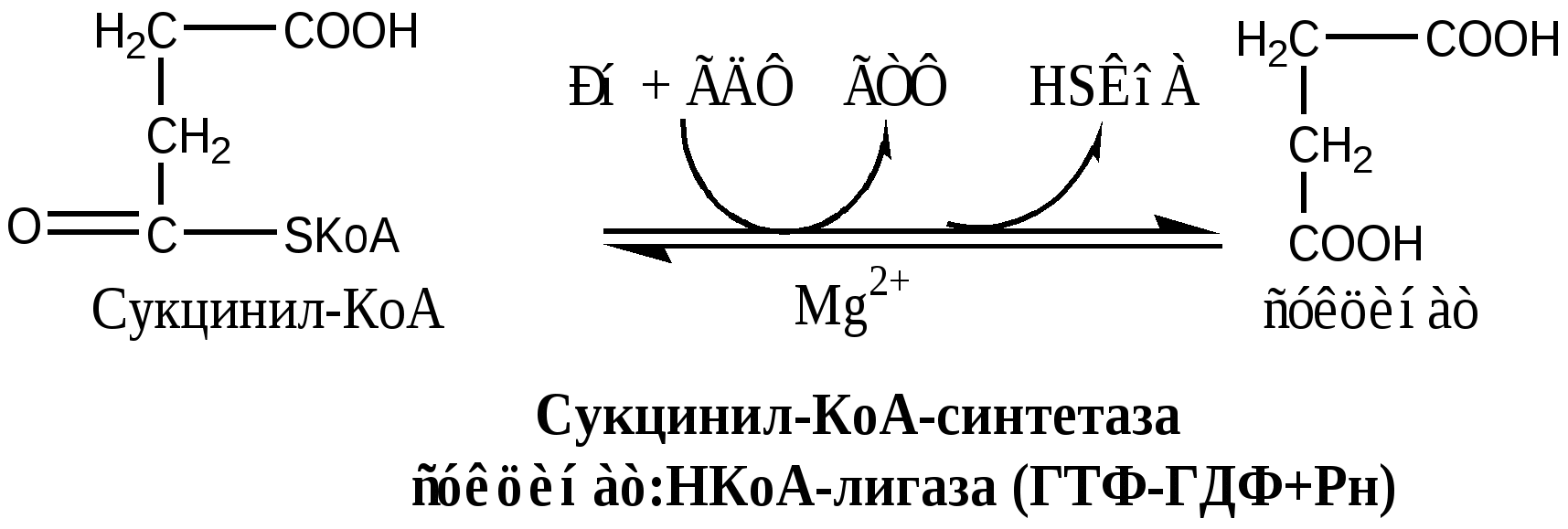 6). Окислительно-восстановительная реакция. Сукцинатдегидрогеназа, флавопротеин содержащий Fe2S2, связана с внутренней мембраной митохондрии. СДГ ингибирует ЩУК и Сукцинил-КоА,  7). Фумараза локализуется в матриксе митохондрий. 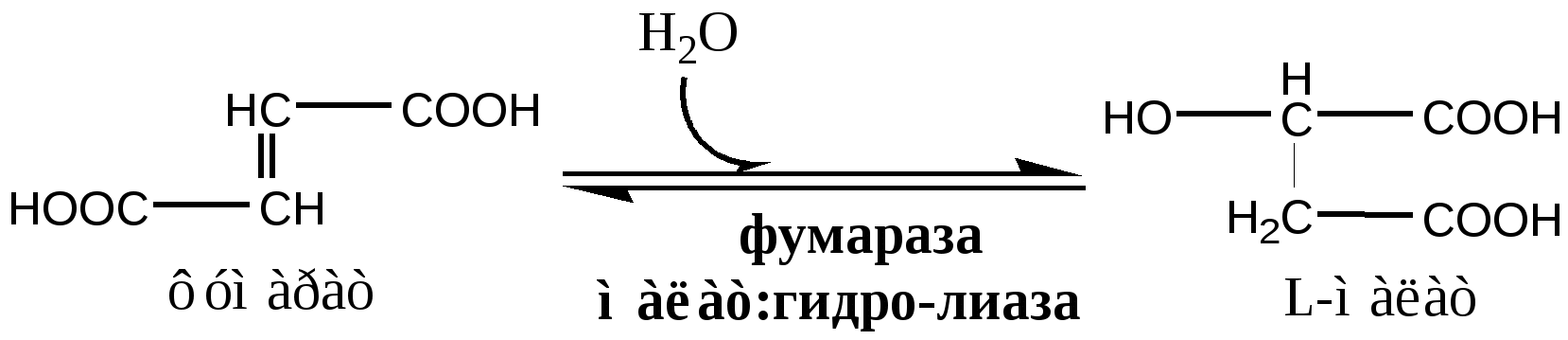 8). Окислительно-восстановительная реакция. Малат ДГ локализуется в матриксе митохондрий. 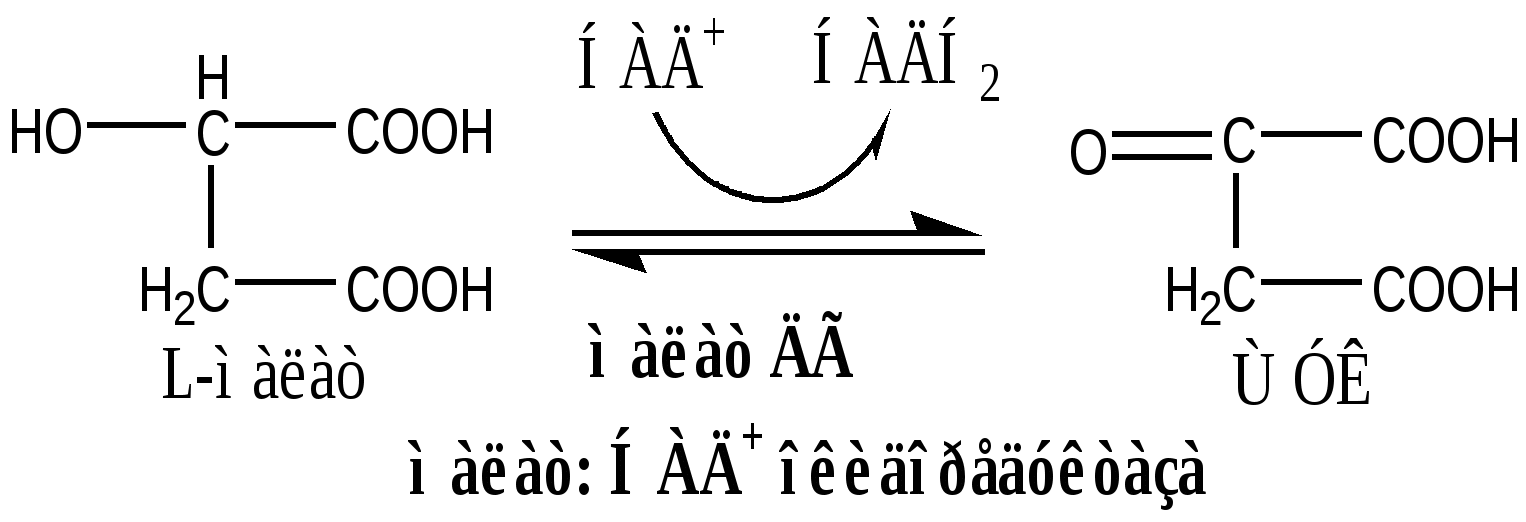 Образовавшиеся молекулы ЩУК реагируют с новой молекулой Ацетил-КоА и цикл повторяется вновь. Энергетический баланс одного оборота ЦТК В 4 окислительно-восстановительных реакциях ЦТК образуются 3 НАДН2 и 1 ФАДН2, которые направляются далее в дыхательную цепь окислительного фосфорилирования. В процессе окислительного фосфорилирования ДЦ из 1 НАДН2 образуется 3 АТФ, из 1 ФАДН2 - 2 АТФ. Из 1 ГТФ, образующейся в ЦТК за счет субстратного фосфорилирования, синтезируется 1 АТФ. Таким образом, за 1 цикл ЦТК из 3 НАДН2, 1 ФАДН2 и 1 ГТФ получается 12 АТФ. ЛЕКЦИЯ № 5 Тема: Окислительное фосфорилирование. Механизмы сопряжения и разобщения, нарушения. |