Биопотенциалы. Биопотенциалы. Понятие и виды биопотенциалов. Природа биопотенциалов. Причина возникновения потенциала покоя. Стационарный потенциал Гольдмана.
 Скачать 139.5 Kb. Скачать 139.5 Kb.
|

|
Биопотенциалы.
Понятия и виды биопотенциалов. Природа биопотенциалов. Биопотенциалы – любые разности потенциалов в живых системах: разность потенциалов между клеткой и окружающей средой; между возбуждённым и невозбуждённым участками клетки; между участками одного организма, находящимися в разных физиологических состояниях. Разность потенциалов - электрический градиент – характерная черта всего живого. Виды биопотенциалов:
Биопотенциалы тесно связаны с метаболическими процессами, следовательно, являются показателями физиологического состояния системы. Величина и характер биопотенциалов являются показателями изменений в клетке в норме и патологии. Существует большая группа электрофизиологических методов диагностики, основанных на регистрации биопотенциалов (ЭКГ, ЭМГ и т.д.). В основе возникновения биопотенциалов лежит несимметричное относительно мембраны распределение ионов, т.е. различные концентрации ионов по разные стороны мембраны. Непосредственная причина – различная скорость диффузии ионов по их градиентам, определяющаяся селективностью мембраны. Биопотенциалы – ионные потенциалы, преимущественно мембранной природы – это основное положение Мембранной теории биопотенциалов (Бернштейн, Ходжкин, Катц). Причина возникновения потенциала покоя. Стационарный потенциал Гольдмана. Натриевый насос – создаёт и поддерживает градиент концентрации иона натрия, иона калия, регулируя их поступление в клетку и выведение из неё. В состоянии покоя клетка проницаема главным образом для ионов калия. Они диффундируют по градиенту концентрации через клеточную мембрану из клетки в окружающую жидкость. Крупные органические анионы, содержащиеся в клетке не могут преодолеть мембрану. Таким образом внешняя поверхность мембраны заряжается положительно, а внутренняя – отрицательно. Изменение зарядов и разности потенциалов на мембране продолжается пока силы, обуславливающие градиент концентрации калия не уравновесятся силами возникающего электрического поля, следовательно, не будет достигнуто стационарное состояние системы. Разность потенциалов через мембрану в этом случае и есть – потенциал покоя. Вторая причина возникновения потенциала покоя – электрогенность калий-натриевого насоса. Теоретическое определение потенциала покоя: При учёте лишь калиевой проницаемости мембраны в состоянии покоя потенциал покоя можно вычислить по уравнению Нернста: R– универсальная газовая постоянная T– абсолютная температура F– число Фарадея СiK – концентрация калия внутри клетки CeK – концентрация калия снаружи клетки На самом деле, помимо ионов калия, клеточная мембрана проницаема также и для ионов натрия и хлора, однако в меньшей степени. Если градиент натрия поступает внутрь клетки, то мембранный потенциал уменьшается. Если градиент хлора поступает внутрь клетки, то мембранный потенциал увеличивается. Если учесть диффузию всех ионов и считать электрическое поле однородным по всей толщине мембраны, то потенциал покоя возможно будет рассчитать по уравнению Гольдмана:  , где , где P – проницаемость мембраны для данного иона. Условия возникновения и фазы потенциала действия. Раздражители – внешние или внутренние факторы, действующие на клетку. При действии раздражителей на клетку меняется электрическое состояние клеточной мембраны. Потенциал действия возникает лишь при действии раздражителя достаточной силы и длительности. Пороговая сила – минимальная сила раздражителя, необходимая для инициации потенциала действия. Раздражители большей силы – надпороговые; меньшей силы – подпороговые. Пороговая сила раздражителя находится в обратной зависимости от его длительности в определённых пределах. Если у раздражителя надпороговой или пороговой силы на участке раздражения возникает электрический импульс характерной формы, распространяющийся вдоль всей мембраны, то возникнет потенциал действия. Фазы потенциала действия:
 - потенциал цитоплазмы - потенциал цитоплазмыд – деполяризация р – реполяризация г – гиперполяризация Фаза деполяризации – быстрая перезарядка мембраны: внутри положительный заряд, снаружи – отрицательный. Фаза реполяризации – возвращение заряда и потенциала мембраны к исходному уровню. Фаза гиперполяризации – временное превышение уровня покоя, предшествующее восстановлению потенциала покоя. Амплитуда потенциала действия заметно превышает амплитуду потенциала покоя – «овершут» (перелёт). Механизм генерации потенциала действия. Потенциал действия – результат изменения ионной проницаемости мембраны. Проницаемость мембраны для ионов натрия – непосредственная функция мембранного потенциала. Если мембранный потенциал понижается, то натриевая проницаемость возрастает. Действие порогового раздражителя: уменьшение мембранного потенциала до критической величины (критическая деполяризация мембраны) → резкое повышение натриевой проницаемости → усиленный приток натрия в клетку по градиенту → дальнейшая деполяризация мембраны → процесс зацикливается → включается механизм положительной обратной связи. Усиленный приток натрия в клетку вызывает перезарядку мембраны и окончание фазы деполяризации. Положительный заряд на внутренней поверхности мембраны становится достаточным для уравновешивания градиента концентрации ионов натрия. Усиленное поступление натрия в клетку заканчивается, следовательно, заканчивается фаза деполяризации. PK : PNa : PCl в состоянии покоя 1 : 0,54 : 0,045, на высоте фазы деполяризации: 1 : 20 : 0,045. В процессе фазы деполяризации проницаемость мембраны для ионов калия и хлора не меняется, а для ионов натрия – возрастает в 500 раз. Фаза реполяризации: увеличивается проницаемость мембраны для ионов калия → усиленный выход ионов калия из клетки по градиенту концентрации → Уменьшение положительного заряда на внутренней поверхности мембраны, обратное изменение мембранного потенциала → уменьшение натриевой проницаемости → обратная перезарядка мембраны → уменьшение калиевой проницаемости, замедление оттока калия из клетки. К концу фазы реполяризации происходит восстановление потенциала покоя. Мембранный потенциал и проницаемость мембраны для ионов калия и натрия возвращается к уровню покоя. Фаза гиперполяризации: возникает, если проницаемость мембраны для ионов калия ещё повышена, а для ионов натрия уже вернулась к уровню покоя. Резюме: Потенциал действия формируется двумя потоками ионов через мембрану. Поток ионов натрия внутрь клетки → перезарядка мембраны. Поток ионов калия наружу → восстановление потенциала покоя. Потоки почти одинаковы по величине, но сдвинуты по времени.  Диффузия ионов через клеточную мембрану в процессе генерации потенциала действия осуществляется по каналам, которые являются высокоселективными, т.е. они обладают большей проницаемостью для данного иона (открытие для него дополнительных каналов). При генерации потенциала действия клетка приобретает определённое количество натрия и теряет определённое количество калия. Выравнивание концентраций этих ионов между клеткой и средой не происходит благодаря калий-натриевому насосу. Методы регистрации и экспериментального исследования биопотенциалов. 1. Внутриклеточное отведение. Один электрод погружают в межклеточную жидкость, другой (микроэлектрод) - вводится в цитоплазму. Между ними – измерительный прибор. Микроэлектрод представляет собой полую трубку, кончик которой оттянут до диаметра в доли микрона, а пипетка наполнена хлоридом калия. При введении микроэлектрода мембрана плотно охватывает кончик, и повреждения клетки почти не происходит. Для создания потенциала действия в эксперименте клетка стимулируется надпороговыми токами, т.е. ещё одна пара электродов связана с источником тока. На микроэлектрод подаётся положительный заряд. 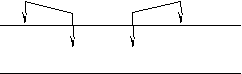 С их помощью можно регистрировать биопотенциалы как крупных, так и мелких клеток, а также биопотенциалы ядер. Но наиболее удобным, классическим объектом исследований, являются биопотенциалы крупных клеток. Например, Nitella ПП 120 мВ (120 * 103 В) Гигантский аксон кальмара ПП 60мВ Клетки миокарда человека ПП 90 мВ 2. Фиксация напряжения на мембране. В определённый момент развитие потенциала действия искусственно прерывается с помощью специальных электронных схем. При этом фиксируется значение мембранного потенциала и величины ионных потоков через мембрану в данный момент, следовательно, есть возможность их измерения. 3. Перфузия нервных волокон. Перфузия – замена оксоплазмы искусственными растворами различного ионного состава. Таким образом, можно определить роль конкретного иона в генерации биопотенциалов. Проведение возбуждения по нервным волокнам.
Роль потенциала действия в жизнедеятельности. Раздражимость – способность живых клеток под влиянием раздражителей (определённых факторов внешней или внутренней среды) переходить из состояния покоя в состояние активности. При этом всегда меняется электрическое состояние мембраны. Возбудимость – способность специализированных возбудимых клеток в ответ на действие раздражителя генерировать особую форму колебания мембранного потенциала – потенциал действия. В принципе возможно несколько видов ответов возбудимых клеток на раздражение, в частности – локальный ответ и потенциал действия. Потенциал действия возникает, если действует пороговый или надпороговый раздражитель. Он вызывает уменьшение мембранного потенциала до критического уровня. Тогда происходит открытие дополнительных натриевых каналов, резкое увеличение натриевой проницаемости и развитие процесса по механизму положительной обратной связи. Локальный ответ возникает, если действует подпороговый раздражитель, составляющий 50-70% от порогового. Деполяризация мембраны при этом меньше критической, наступает лишь кратковременное, небольшое увеличение натриевой проницаемости, механизм положительной обратной связи не включается, и потенциал быстро возвращается к исходному состоянию. В процессе развития потенциала действия возбудимость меняется. Снижение возбудимости – относительная рефрактерность. Полная утрата возбудимости – абсолютная рефрактерность. По мере приближения к уровню критической деполяризации возбудимость повышается, так как для достижения этого уровня и развития потенциала действия становится достаточно и небольшого изменения мембранного потенциала. Именно так меняется возбудимость в начале фазы деполяризации, а также при локальном ответе клетки на раздражение. При удалении мембранного потенциала от критической точки возбудимость снижается. На пике фазы деполяризации, когда клетка уже не может реагировать на раздражение открытием дополнительных натриевых каналов, наступает состояние абсолютной рефрактерности. По мере реполяризации абсолютная рефрактерность сменяется относительной; к концу фазы реполяризации возбудимость снова увеличена (состояние «супернормальности»). Во время фазы гиперполяризации возбудимость снова снижена. Возбуждение – ответ специализированных клеток на действие пороговых и надпороговых раздражителей – это сложный комплекс физико-химических и физиологических изменений, в основе которого лежит потенциал действия. Результат возбуждения зависит от того, в какой ткани оно развивалось (где возник потенциал действия). К специализированным возбудимым тканям относятся:
Потенциалы действия обеспечивают проведение возбуждения по нервным волокнам и инициируют процессы сокращения мышечных и секреции железистых клеток. Потенциал действия, возникающий в нервном волокне – нервный импульс. Об аксонах. Аксоны (нервные волокна) – длинные отростки нервных клеток (нейронов). Афферентные пути – от органов чувств к ЦНС Эфферентные пути – от ЦНС к мышцам. Протяжённость – метры. Диаметр в среднем от 1 до 100 мкм (у гигантского аксона кальмара – до 1 мм). По наличию или отсутствию миелиновой оболочки различают аксоны:
Миелиновая оболочка – окружающая аксон дополнительная многослойная (до 250 слоёв) мембрана, образующаяся при внедрении аксона в шванновскую клетку (леммоцит, олигодендроцит), и многократном наматывании мембраны этой клетки на аксон. Миелин – очень хороший изолятор. Через каждые 1-2 мм миелиновая оболочка прерывается перехватами Ранвье, протяжённостью около 1 мкм каждый.  Только в области перехватов возбудимая мембрана контактирует с внешней средой. Кабельная теория проведения. Аксон по ряду свойств подобен кабелю: это полая трубка, внутренне содержимое – аксоплазма – проводник (как и межклеточная жидкость), стенка – мембрана – изолятор. Механизм проведения возбуждения (распространения нервного импульса) включает 2 ступени:
Локальные токи циркулируют между возбужденным и невозбуждённым участками нервного волокна ввиду разной полярности мембраны на этих участках. 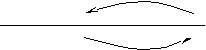 Внутри клетки они текут от возбуждённого участка к невозбуждённому. Снаружи – наоборот. Локальный ток вызывает сдвиг мембранного потенциала соседнего участка, и начинается распространение волны деполяризации по волокну, как тока по кабелю. Когда деполяризация очередного участка достигает критической величины, происходит открытие дополнительных натриевых, потом калиевых каналов, возникновение потенциала действия. В разных участках волокна потенциал действия формируется независимыми ионными потоками, перпендикулярными к направлению распространения. При этом на каждом участке происходит энергетическая подпитка процесса, так как градиенты ионов, идущих по каналам, создаются насосами, работа которых обеспечивается энергией гидролиза АТФ. Роль локальных токов – лишь инициация процесса путём деполяризации всё новых участков мембраны до критического уровня. Благодаря энергетической подпитке нервный импульс распространяется вдоль волокна без затухания (с неизменной амплитудой). Направление и скорость проведения. Одностороннее проведение нервного импульса обеспечивают:
Скорость проведения тем выше, чем более выражены кабельные свойства волокна. Для их оценки применяют константу длины нервного волокна: D – диаметр волокна bm – толщина мембраны Физический смысл константы: она численно равна расстоянию, на котором подпороговый потенциал уменьшился бы в e раз. С увеличением константы длины нервного волокна увеличивается и скорость проведения. Ионный состав аксоплазмы одинаков, следовательно, Беспозвоночные: увеличение диаметра волокна. Позвоночные: увеличение толщины и удельного сопротивления мембраны путём приобретения миелиновой оболочки, следовательно, происходит качественное изменение способа проведения. Непрерывное и сальтаторное проведение. Проведение возбуждения по амиелиновым волокнам – непрерывное: в своё время, последовательно, один за другим каждый участок волокна переходит в состояние возбуждения. Проведение по миелиновым волокнам – сальтаторное (скачкообразное).  Возбудимая мембрана мякотного аксона обнажена только перехватами Ранвье, только там открываются натриевые и калиевые каналы, только в них происходит генерация потенциала действия. При возбуждении одного перехвата локальные токи образуются между ним и соседним, и импульс «перескакивает» с одного перехвата на другой. По миелинизированному волокну импульс распространяется на порядок быстрее. Это обеспечивает позвоночным эволюционное преимущество. Электрический диполь – система, состоящая из двух равных, но противоположных по знаку точечных электрических зарядов, расположенных на некотором расстоянии друг от друга (плечо диполя l). Основная характеристика диполя – дипольный момент – вектор, равный произведению заряда на плечо диполя, направленный от отрицательного заряда к положительному. Обозначается 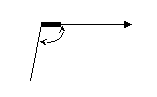 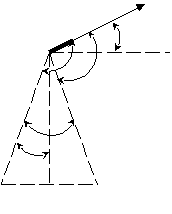  По Эйнтховену:      Кардиограмма представляет собой проекцию вектора дипольного момента сердца на направление соответствующего отведения, развёрнутую во времени. Теория ЭКГ предполагает электрический диполь рассматривать в среде безграничного однородного диэлектрика. А это не человек. Теория ЭКГ на сегодняшний день не завершена. Блочная схема ЭКГ: 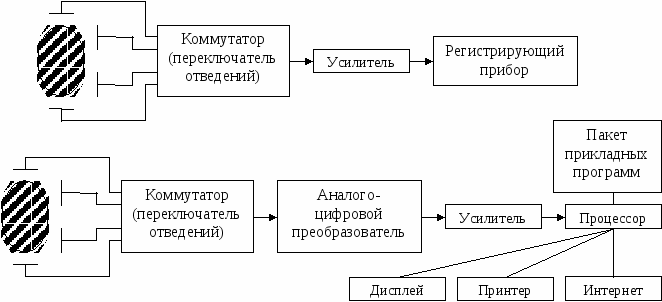 Модель эквивалентного токового генератора. Эквивалентная схема возникновения биотока:  r – сопротивление источника R – внешняя нагрузка Сильный источник, слабая внешняя нагрузка => система работает в состоянии короткого замыкания:  Токовый диполь:   - потенциал в точке униполя, - потенциал в точке униполя, Потенциал токового диполя: G можно пренебречь. Таких диполей на поверхности органа множество. Но можно выделить суммарный потенциал D0.  |