Сердюкова ЛС Структура и функции эритроцитарных клеток. Федеральное государственное бюджетное образовательное учреждение высшего образования воронежский государственный университет
 Скачать 289 Kb. Скачать 289 Kb.
|

|
| Обучающийся | | | Л.С. Сердюкова |
| Преподаватель | | канд. биол. наук | Ю.А. Лысенко |
Воронеж 2017
Оглавление
Оглавление…………………………………………………………………….2
Введение……………………………………………………………………….3
Глава 1. Структурные характеристики эритроцитарных клеток…………..4
. Особенности внешнего строения……………………………....4
. Строение мембраны эритроцита……………………………….7
. Внутреннее строение…………………………………………..14
. Метаболизм эритроцитарных клеток……………………...….16
. Эритропоэз……………………………………………………...17
Глава 2. Функции эритроцитарных клеток…………………………………..19
Заключение………………………………………………………………….….21
Список использованных источников……………………………………...….22
Введение
Кровь – важнейшая часть внутренней среды организма, выполняющая многообразные физиологические функции, состоит из жидкой среды — плазмы и взвешенных в ней клеток — форменных элементов: эритроцитов, лейкоцитов и тромбоцитов. Самые многочисленные из них – красные кровяные тельца – эритроциты (от греч. erythros – красный и kytos – вместилище, клетка). Именно они определяют реологическое поведение крови в сосудах. Суммарный объем эритроцитов в 50 раз больше объема лейкоцитов и тромбоцитов, а масса эритроцитов в 750 раз превышает массу лейкоцитов. У мужчин в 1 мкл крови их содержится в среднем 5,1 млн, а у женщин - 4,6 млн. У новорожденных число эритроцитов довольно высоко (5,5 млн/мкл крови), что обусловлено перемещением крови из плаценты в кровоток ребенка во время родов и значительной потерей воды в дальнейшем [30]. К году жизни их количество снижается до 3,6 – 4,9 млн/мкл, а в подростковом возрасте немного растёт (3,6 – 5,1 млн/мкл). Анемия – это снижение уровня эритроцитов, зависит от разных причин. Эритроцитоз – рост количества эритроцитарных клеток, возможен при значительном обезвоживании или при патологии крови, связанной с усиленным синтезом эритроцитов.
По изменению свойств эритроцитарных клеток (например, скорость оседания, деформируемость, форма, содержание гемоглобина и др.), их уровня в крови можно судить не только о состоянии здоровья человека, но и о наличии патологии в организме. Именно поэтому очень важно иметь представление о функциях, выполняемых эритроцитами, и о структурных особенностях этих клеток.
Глава 1. Структурные характеристики эритроцитарных клеток
1.1. Особенности внешнего строения
Эритроциты человека – маленькие эластичные клетки диаметром 7.2-7.9 мкм в высушенном препарате и 8.3-8.7 мкм во влажном (размеры нормального эритроцита изменчивы и зависят от методов определения и возраста клетки). Обнаруживаются в небольшом количестве и эритроциты диаметром от 5,5 до 6,5 мкм (микроциты), а также 8,5-9,0 мкм (макроциты). Появление в крови микроцитов происходит под влиянием неблагоприятных факторов (дефицит железа, белка, микроэлементов, витаминов, солнечного света). Увеличение диаметра эритроцитов происходит при так называемом стрессовом эритропоэзе, когда вызревание эритроцитов происходит, минуя несколько этапов деления эритроидных клеток костного мозга для быстрого пополнения периферической крови эритроцитами и улучшения снабжения тканей кислородом при неблагоприятных условиях. Неблагоприятные факторы могут приводить к истощению компенсаторных систем, в связи с чем в кровотоке появляются одновременно микро- и макроциты [7].
В нормальной крови человека основную массу (80-90%) составляют дискоциты – эритроциты двояковогнутой формы, с утолщениями по краям и центральной впадиной (пеллор), на которую приходится 35-55% поверхности. Поддержание дисковидной формы обусловлено отрицательным осмотическим давлением внутри клетки, состоянием мембраны и стромы эритроцита. При данной форме площадь поверхности на 20% больше, чем при сферической (минимальной для данного объема), отношение площади поверхности к объему самое высокое, любая молекула гемоглобина находится близко к поверхности, что обеспечивает максимальную скорость газообмена [6]. Кроме того, имеются планоциты (с плоской поверхностью) и стареющие формы эритроцитов – шиповидные эритроциты, или эхиноциты (6 %), куполообразные, или стоматоциты (1-3 %), и шаровидные, или сфероциты (1 %) (рис.1).
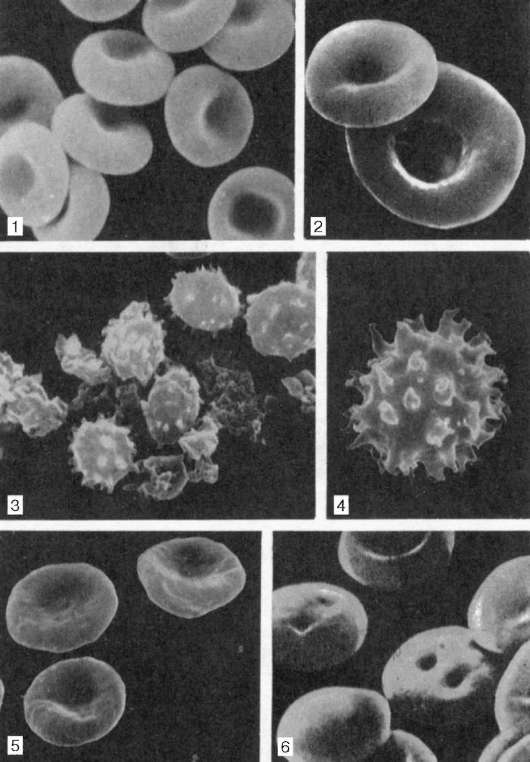
Рис. 1. Эритроциты различной формы в сканирующем электронном микроскопе, ув. 8000 (по Г.Н. Никитиной): 1 – дискоциты-нормоциты; 2 – дискоцит-макроцит; 3, 4 – эхиноциты; 5 – стоматоциты; 6 –сфероциты
При заболеваниях могут появляться аномальные формы эритроцитов, что чаще всего обусловлено изменением структуры гемоглобина (Нb). Замена даже одной аминокислоты в молекуле Нb может быть причиной изменения формы эритроцитов. Например, при серповидноклеточной анемии появление эритроцитов серповидной формы связано с генетическим повреждением гемоглобина, в молекуле которого вместо глутаминовой кислоты находится валин. Процесс нарушения формы эритроцитов при заболеваниях получил название пойкилоцитоз [30].
Толщина эритроцитарной клетки изменяется от 1.7-2.5 мкм в сухом до 1.5-1.7 мкм во влажном препарате. Площадь поверхности клетки составляет 128-160 мкм², а объём – 76-96 мкм³. Среднее содержание гемоглобина в эритроците составляет 26-34 пикограмм/эритроцит [8]. В норме размеры эритроцитов человека и содержание в них гемоглобина различаются в зависимости от возраста, пола, климатогеографических условий проживания, времени суток, а также от места забора крови для анализа. У женщин размеры и количество эритроцитов несколько меньше, чем у мужчин. Кровь мужчин содержит больше клеток в единице объема, в ней больше гемоглобина, более высокий гематокрит и объем эритроцита [27].
Среди факторов, определяющих форму эритроцита, можно выделить следующие:
цитоскелет (система мембранных белков);
химический состав липидной компоненты мембраны, возможная неоднородность его вдоль мембраны;
концентрация ионов, АТФ;
электростатические факторы: поверхностный заряд мембраны и состояние ионизации белков цитоскелета;
cостояние молекул гемоглобина;
cостояние внутриклеточных структур.
Особенности цитоскелета и клеточной мембраны позволяют эритроцитам претерпевать значительные изменения и восстанавливать форму. Это свойство эритроцитарных клеток получило название деформируемость. При воздействии сдвиговых напряжений на эритроцит или при его прохождении через капилляр, диаметр которого меньше диаметра эритроцита, он может принимать форму чаши, груши, колокольчика, песочных часов, а после снятия воздействия возвращаться к исходным параметрам. Эритроциты человека диаметром 8 мкм проходят через капилляры диаметром 2-5 мкм. Способность эритроцитов к деформации определяется внутренней (цитоплазматической) вязкостью, вязкостно-эластическими свойствами мембраны и отношением площади клетки к её объёму. Для исследования деформируемости эритроцитов используют различные методы: фильтрацию, зондовую сканирующую микроскопию, лазерную дифрактометрию с компьютерным анализом изображений и другие [1, 18].
Строение мембраны эритроцита
Массовая доля мембраны составляет 10% от клеточной. В ее составе 90% белков и только 10% липидов. Это делает эритроциты особенными клетками организма, так как почти во всех остальных мембранах липиды преобладают над белками. Именно плазмалемма определяет гомеостаз и функциональное состояние эритроцита. От нее зависят процессы взаимодействия эритроцита с окружающей средой, активность мембранассоциированных ферментов, транспорта ионов и газообмена, а также длительность нахождения эритроцита в периферическом кровотоке. Мембрана эритроцитов содержит набор ферментов гликолиза, пентозофосфатного цикла, системы глутатиона, адениловой системы и других реакций обмена для реализации анаэробного пути усвоения энергии и являются важнейшим компонентом антиоксидантной системы организма. Структура мембран эритроцитов лабильна, и под воздействием ряда факторов возможна ее деструкция [6]. Перенос веществ через мембрану совершается в зависимости от их химических свойств разными способами: путем диффузии (вещества в виде раствора проходят через заполненные водой мембранные поры), путем проникновения через липидные участки (если вещества растворимы в жирах). Некоторые вещества могут взаимодействовать со встроенными в мембрану молекулами-переносчиками, образуя с ними легкообратимую связь, и в дальнейшем проходить через мембрану либо пассивно, либо в результате так называемого активного транспорта [30].
Несмотря на видовую специфичность эритроцитов, плазмалемме эритроцитов человека и животных присущи общие принципы молекулярной организации. Выделяют следующие структурные элементы:
1. Двойной слой липидов. Он обеспечивает барьер между окружающей средой и цитоплазмой эритроцита. На его долю приходится 50-60% от массы эритроцитарной мембраны. При этом следует учесть, что синтез фосфолипидов и холестерина в зрелых эритроцитах не возможен. Липиды мембран эритроцитов делят на 3 класса: нейтральные, гликолипиды и фосфолипиды.
Нейтральные липиды, составляют до 30% массы всех липидов и представлены глицеридами, холестерином и его эфирами [6]. Многочисленные экспериментальные данные подтверждают зависимость устойчивости мембраны эритроцитов к воздействию различных повреждающих агентов от содержания в них холестерина [25, 28, 36]. Искусственное снижение концентрации холестерина вызывает дестабилизацию мембраны, усиление её повреждений после инкубации с окислителем, что проявляется в снижении осмотической резистентности и усилении гемолиза эритроцитов. Избыточное накопление холестерина сопряжено с изменением физико-химических свойств эритроцитарной мембраны: повышением её микровязкости, ухудшением деформируемости клеток, способности к прохождению в микроциркуляторном русле [21].
Гликолипиды составляют около 10% мембранных липидов и являются гликосфинголипидами (нейтральными и кислыми). Все они представляют собой амфипатические молекулы, т. е. у них есть гидрофильный и гидрофобный концы.
Фосфолипиды – это наиболее широко представленный класс липидов в эритроцитарной мембране (около 60%). Они являются производными либо сфингозалина (сфинголипиды: сфингомиелин), либо глицерина (глицерофосфолипиды: фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, фосфатидилинозитол, фосфатидная кислота и 35 видов жирных кислот).
В состав мембраны также входят свободные жирные кислоты, триглицериды, лизофосфатидилэтаноламин, фосфатидилинозит (6%). Распределение липидов в эритроцитарной мембране ассиметрично: на ее внешней стороне концентрируются сфингомиелин (26% от всех липидов) и фосфатидилхолин (28%), на внутренней – фосфатидилсерин (13%), фосфатидилэтаноламин (27%) и аминофосфолипиды. Поддержание ассиметричного расположения липидов в бислое контролируется рядом ферментов. Липиды эритроцитарной мембраны регулируют подвижность внутримембранных белков, обеспечивая нормальное функционирование мембранассоциированных ферментов и рецепторов, а также чувствительность рецепторов к лигандам, регулируют трансмембранный транспорт веществ, иммунный ответ эритроцита и функционирование вторичных мессенджеров межклеточного и внутриклеточного взаимодействия, в том числе инозитол-1,4,5-трифосфата и диацилглицерола, который активирует Са2+-фосфолипидзависимую протеинкиназу С и регулирует работу Са2+-АТФ-азы [6].
2. Белки. Они обеспечивают поддержание формы эритроцита, определяют его механические свойства, осуществляют связь между скелетом мембраны и липидным слоем, трансмембранный транспорт молекул, реализуют ферментативную активность, содержат антигены узнавания для аутологических антител к стареющим и аномальным эритроцитам, ответственны за анионный транспорт в эритроците, участвуют в газообмене. Методом электрофореза в мембране эритроцитов обнаружено около 15 основных мембранных белков с молекулярной массой 15-250 кДа. Условно они разделяются на интегральные (встроены в липидный слой) и периферические (цитоплазматические, находятся на наружной или внутренней поверхности клеточной мембраны и нековалентно связаны с интегральными мембранными белками). Многие мембранные белки пронизывают бислой насквозь (трансмембранные белки), обладают амфипатическими свойствами: у них есть гидрофобные участки, проходящие через мембрану и взаимодействующие с гидрофобными хвостами липидных молекул внутри бислоя, и гидрофильные участки, взаимодействующие с фосфатными группами фосфолипидов. По функциональному значению делятся на формирующие мембранный скелет (спектрины, анкирин, актин и др.) и на белки, обеспечивающие метаболизм и ионный гомеостаз клетки (анионный канал, гликофорины, аддуцин, ацетилхолинэстераза, Nа+, К+-АТФ-аза, Са2+-АТФ-аза, карбоангидраза). Регуляторная функция белков осуществляется путем их фосфорилирования и дефосфорилирования.
Белок спектрин является основным белком, составляющим структуру цитоскелета (25% массы всех белков мембраны), участвует в поддержании двояковогнутой формы эритроцита, регулирует подвижность белков и удерживает в равновесии двойной слой липидов. Молекулы спектрина имеют вид палочек диаметром 2 нм и длиной около 100 нм, образуют сеть, которая фиксируется на цитоплазматической стороне плазмалеммы с помощью анкирина и белка полосы 3 или актина, белка полосы 4.1 и гликофорина.
Белок анкирин – крупный белок, его молекула состоит из 1881 аминокислоты, связывает спектриновый цитоскелет с плазмалеммой.
Гликофорин А – трансмембранный белок, пронизывает плазмалемму в виде одиночной спирали, является основным сиалогликопротеидом мембраны эритроцита (75% от всех сиалогликопротеидов), содержит большинство сиаловых кислот, обладает антигенной активностью, несет рецепторы к вирусу гриппа и фитогемагглютинину [6].
Белок полосы 3 – основной интегральный трансмембранный гликопротеид-переносчик ионов Cl- и HCO3-, назван так по расположению этой белковой фракции на электрофореграмме относительно других белков. Связан со спектрином посредством анкирина [5] (рис.2).
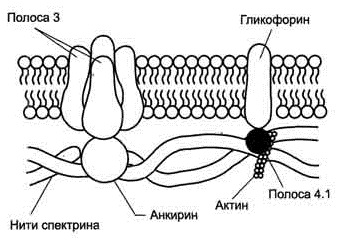
Рис. 2. Строение околомембранного белкового комплекса
За перенос различных полярных молекул, таких, как сахара, аминокислоты, нуклеотиды и многие другие метаболиты, через клеточные мембраны ответственны специфические белки, называемые мембранными транспортными белками. Эти белки обеспечивают перенос специфических веществ через мембрану без непосредственного контакта с гидрофобной внутренностью липидного бислоя, формируя в нем сквозные проходы. Существуют два основных класса мембранных транспортных белков: белки-переносчики и каналообразующие белки. Белки-переносчики связывают молекулу переносимого вещества, что приводит к их конформационным изменениям и как результат к переносу этой молекулы через мембрану. Каналообразующие белки формируют заполненные водой поры, пронизывающие липидный бислой. Когда эти поры открыты, молекулы специфических веществ (обычно неорганические ионы подходящего размера и заряда) проходят сквозь них и, следовательно, через мембрану.
Все каналообразующие белки и многие белки-переносчики позволяют растворенным веществам проходить через мембраны только пассивно (пассивный транспорт, облегченная диффузия). Среди белков, обеспечивающих ионный гомеостаз эритроцита, наиболее важными является белок, образующий анионный канал. Плазмалемма каждого эритроцита содержит около 600000 анионных каналов, на которые приходится 10% поверхности мембраны. Через данный канал в обе стороны (по градиенту концентрации) проходят анионы (Сl-, НСО3-, ОН-) и глюкоза. Благодаря анионным каналам реализуется эффект Гиббса-Доннана: отрицательно заряженный гемоглобин выталкивает анионы из клетки, отчего их концентрация в эритроцитах значительно ниже, чем в плазме, а pH содержимого эритроцита (7,22) меньше pH плазмы (7,40). Катионы (Nа+, К+) не проходят через мембрану эритроцита по градиенту концентрации, так как мембрана эритроцита не содержит катионные каналы, то есть она не имеет специальных систем, предназначенных для пассивного транспорта катионов. Но она содержит систему белков, выполняющих функцию катионных насосов, т.е. переносящих катионы (Nа+, К+, Н+, Са2+) против градиента концентрации. В частности, Na+, К+-АТФ-аза, используя энергию АТФ, переносит Nа+ и К+ против градиента концентрации: ионы Nа+ из эритроцита, а ионы К+ – в эритроцит. Отсутствие К+-каналов (выход К+ из клетки по градиенту концентрации) способствует сохранению отрицательного заряда на внешней поверхности мембраны, что принципиально отличает эритроциты от клеток возбудимых тканей – нейронов, миокардиоцигов, миоцитов и мышечных волокон. При снижении активности Na+, K+-АТФ-азы концентрация Na+ в клетке повышается, что приводит к увеличению осмотического давления, увеличению поступления воды в эритроцит и к его гибели в результате гемолиза. Ca2+-АТФ-аза осуществляет выведение ионов кальция из эритроцитов.
3. Цитоскелет – соединение белков друг с другом и с мембраной, расположенное вдоль внутренней (обращённой к цитоплазме) стороны плазмалеммы. Делает прочной основную мембрану, обеспечивая единство ее липидного слоя и в то же время придавая ей внутреннюю подвижность и гибкость [6] (рис.3).
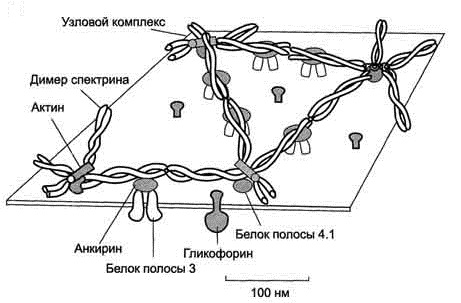
Рис. 3. Строение цитоскелета эритроцита
4. Гликокалликс. Сформирован олигосахаридами, гликолипидами и гликопротеидами, расположенными на наружной поверхности мембраны. Они определяют антигенные свойства эритроцитов. При связывании этих антигенов (агглютиногенов А и В) с соответствующими антителами (агглютинины α и β) происходит агглютинация – склеивание эритроцитов. В норме в плазме крови нет агглютининов к собственным эритроцитам, в противном случае возникает аутоиммунное разрушение эритроцитов [5].
Плазмалемма эритроцита несёт на себе резус-фактор, рецепторы инсулина, соматотропного гормона, а также рецепторы к лектинам и другим биологически активным веществам. Взаимодействие с указанными веществами приводит к метаболическим изменениям, трансформации эритроцита, к дефосфолирированию клеточных белков, что в конечном итоге влияет на функции эритроцитарной клетки [6]. Также на поверхности мембраны происходит адсорбция веществ. В норме преимущественно переносятся атерогенные липиды. К ним относятся липопротеиды низкой и очень низкой плотности. По количеству молекул липиды нередко занимают первое место среди органических веществ, адсорбированных на эритроцитах. Адсорбция белка влияет на деформируемость и реологию эритроцитов. Увеличение адсорбции на эритроцитах глюкозы или белков с повышенной химической активностью ведёт к частичному «сбросу» в плазму липидов, включая атерогенные. Считается, что поступлением в стенку сосудов атерогенных липидов обусловлен атеросклероз. О риске его возникновения, а также о начальных нарушениях липидного обмена можно судить по показателям транспорта липидов, белка и глюкозы на поверхности форменных элементов крови [1, 10].
Исследование морфометрических показателей, определение характера и глубины нарушения проницаемости мембран эритроцитов имеет большое значение для выявления патогенеза [35]. Доказана взаимосвязь между изменениями свойств эритроцитов и клеточных мембран внутренних органов, поэтому эритроциты наиболее удобны для исследования проницаемости всех биологических мембран и оценки резистентности эритроцитов при патологии и в различных физиологических состояниях. Установлено изменение осмотической резистентности эритроцитов при различных заболеваниях, выявлено её снижение при онкологической патологии, воздействии вредных производственных факторов, травме головного мозга. Нарушение проницаемости мембран эритроцитов было выявлено у больных ангинами и паратонзиллитами [16, 22, 32, 33].
1.3. Внутреннее строение
Эритроцит состоит из воды (70%), гемоглобина (25%), а также липидов, сахаров, солей, ферментных белков, в целом на долю которых приходится остальные 5%. Содержимое эритроцитов – это идеальная ньютоновская жидкость, вязкость которой зависит от концентрации гемоглобина [6].
В зрелых эритроцитарных клетках млекопитающих отсутствует клеточное ядро и большинство органелл, что повышает содержание гемоглобина [26]. У остальных позвоночных эритроциты имеют ядро. Зрелые эритроциты лишены ещё и рибосом, поэтому нового гемоглобина не накапливают, а лишь транспортируют и используют уже имеющийся в течение отведённого им срока жизни. Они также лишены поверхностных рецепторов к трансферрину (то есть не способны более захватывать и усваивать дополнительное железо) [34].
Цитоплазма эритроцита состоит из воды (60%) и сухого остатка (40%), содержащего, в основном, гемоглобин (от др.-греч. – кровь и лат. globus – шар) – белок, состоящий из 4-х полипептидных цепей глобина и небелковой части – гема В (комплексного соединения порфирина с двухвалентным железом). 5% сухого остатка стромы эритроцитарной клетки приходятся на органические (глюкоза и промежуточные продукты её катаболизма) и неорганические вещества. Из ферментов в цитоплазме присутствуют ферменты гликолиза, антиоксидантной защиты и метгемоглобинредуктазной системы, карбоангидраза [5].
Гемоглобин обладает высокой способностью связывать кислород (O2), диоксид углерода (CO2), угарный газ (CO). При высоком парциальном давлении кислорода гемоглобин в лёгких соединяется с ним, образуется оксигемоглобин ярко-алого цвета. Гемоглобин, отдавший кислород тканям, носит название дезоксигемоглобин, или восстановленный (HbH). Отдав кислород тканям, гемоглобин присоединяет к себе ионы водорода и углекислый газ, образуется карбоксигемоглобин (HbCO2), имеет тёмно-вишнёвую пигментацию. При небольшой нехватке железа эритроциты продуцируются с той же скоростью, что и раньше, но при этом содержат в себе меньше гемоглобина. Сначала при небольшом дефиците железа начинает уменьшаться объём эритроцитов и концентрация гемоглобина в них, при нарастании дефицита железа начинает уменьшаться количество эритроцитов [3]. Гемоглобин высокотоксичен при попадании значительного его количества из эритроцитов в плазму крови, что происходит при патологических состояниях. Его токсичность проявляется тканевой гипоксией – ухудшением кислородного снабжения тканей.
1.4. Метаболизм эритроцитарных клеток
Обмен веществ зрелых безъядерных эритроцитов направлен на обеспечение их функции как переносчиков кислорода, а также их участия в переносе диоксида углерода. В связи с этим метаболизм эритроцитов отличается от метаболизма других клеток. Он должен, прежде всего, поддерживать способность эритроцита обратимо связывать кислород, для чего необходимо восстановление иона железа в составе гема. Двухвалентное железо в нем постоянно переходит в трехвалентное вследствие спонтанного окисления и, для того чтобы могло происходить связывание кислорода, Fe(III) должно быть восстановлено в Fe(II) [30].
Зрелые эритроциты лишены митохондрий, поэтому в качестве энергетического материала для гликолиза они могут использовать только глюкозу. В эритроцитах катаболизм глюкозы обеспечивает сохранение структуры и функции гемоглобина, целостность мембран и образование энергии для работы ионных насосов. Глюкоза поступает в эритроцитарные клетки путём облегченной диффузии [5]. В цитоплазме эритроцитов содержатся ферменты анаэробного гликолиза, с помощью которых синтезируются АТФ и НАДН, обеспечивающие энергией главные процессы, связанные с переносом О2 и СО2, а также поддержание осмотического давления и перенос ионов через плазмалемму эритроцита. Энергия гликолиза обеспечивает активный транспорт катионов через плазмалемму, поддержание оптимального соотношения концентрации К+ и Na+ в эритроцитах и плазме крови, сохранение формы и целостности мембраны эритроцита [30].
1.5. Эритропоэз
Жизненный цикл эритроцита в кровотоке в среднем составляет 100-120 дней. Старение эритроцитов связано со снижением активности ферментных систем. Начиная с 60-го дня после выхода эритроцита в периферическую кровь, в нем прогрессивно снижается активность ферментов, что приводит к уменьшению энергетического потенциала клетки, нарушению способности эритроцитов поддерживать градиент натрия и калия, к росту концентрации ионов Са2+ внутри эритроцита, к повышению содержания метгемоглобина. Кроме того, при старении уменьшается содержание фосфолипидов и холестерина, меняется их соотношение при неизменном сохранении содержания мембранных белков. По мере старения эритроциты принимают сферическую форму, что препятствует их прохождению через внутриэндотелиальные синусы селезенки; это способствует их осмотическому лизису и фагоцитированию. Ежедневно в норме разрушается около 200,0x10^9 эритроцитов/л (0,8% или 1 столовая ложка) и столько же выходит в периферическую кровь [27]. Утилизация старых или поврежденных эритроцитов производится макрофагами главным образом в селезенке, а также в печени и костном мозге, при этом гемоглобин распадается, а высвобождающееся из гема железо используется для образования новых эритроцитов [5].
После кровопотери и при патологическом укорочении жизни эритроцитов скорость эритропоэза может возрастать в несколько раз. Мощным стимулятором эритропоэза служит снижение парциального давления О2 (т.е. несоответствие между потребностью ткани в кислороде и его поступлением). При этом возрастает содержание в плазме особого вещества – эритропоэтина – термостабильного гликопротеина, который стимулирует дифференцировку и ускоряет размножение предшественников эритроцитов в костном мозгу [30].
Эритроциты образуются в кроветворных тканях: желточном мешке у эмбриона, печени и селезенке у плода и красном костном мозгу плоских костей у взрослого [30]. В процессе созревания эритроцитарная клетка проходит несколько последовательных стадий дифференциации:
Гемангиобласт (первичная стволовая клетка)
Гемоцитобласт (плюрипотентная гемопоэтическая стволовая клетка)
СFU-GEMM (мультипотентная гемопоэтическая клетка)
СFU-Е (унипотентная гемопоэтическая клетка)
Пронормобласт (проэритробласт, рубрибласт)
Базофильный, ранний нормобласт (эритробласт, прорубрицит)
Полихроматофильный, промежуточный нормобласт (эритробласт, рубрицит)
Ортохроматический, поздний нормобласт (эритробласт, метарубрицит). На этой стадии клетка утрачивает способность к синтезу ДНК и делению, а потом и ядро. После его потери остаются следовые количества мРНК и небольшое количество рибосом, которые сохраняются еще в течение 2 — 3 суток во время превращения нормобласта в ретикулоцит. Наряду с этими процессами растет количество гемоглобина в клетке.
Ретикулоцит, «юный» эритроцит [34]. Содержит митохондрии, рибосомы, цитоплазматическую сеть, осуществляет расщепление жирных кислот, активный транспорт катионов, сохраняет способность к синтезу белка (глобина), гема, включению железа в гемоглобин. После созревания ретикулоциты выходят из костного мозга в общее кровеносное русло. После 1-2 дней пребывания в системном кровотоке заканчивают созревание и превращаются в зрелые эритроциты (этот процесс продолжается 1-3 суток). При этом происходит полная деградация рибосом, частичное нарушение клеточной мембраны и избирательный распад ключевых ферментов, находящихся в органеллах цитоплазмы. Прекращается большая часть синтетических процессов, нарушается способность к синтезу белка, образованию гема, синтезу липидов; инактивируются флавиновые ферменты и цитохромоксидазы [6].
Глава 2. Функции эритроцитарных клеток
Среди основных функций эритроцитарных клеток можно выделить следующие:
1. Газотранспортная. Эритроцит переносит кислород, не потребляя его и не расходуя при этом энергии. Перенос кислорода от альвеол легких к тканям и углекислого газа от тканей к легким реализуется с участием гемоглобина и карбоангидразы. Эритроциты участвуют также в транспорте аминокислот, антител, токсинов и ряда лекарственных веществ, адсорбируя их на поверхности плазмалеммы [9, 14, 19, 25, 31].
2. Буферная. Осуществляется за счет гемоглобина - одного из самых мощных компонентов буферной системы крови [9, 14, 19, 25]. При нормальных пределах pH оксигемоглобин является более сильной кислотой, чем дезоксигемоглобин. Гемоглобин, освобождаясь в тканях от кислорода, приобретает способность к связыванию ионов H+, а в венозной крови в результате выделения CO2 тканями происходит накопление в крови этих ионов [27].
3. Питательная. Эритроциты переносят на своей поверхности аминокислоты, холестерин, глюкозу, витамины В1, В2, В6, С от органов пищеварения к клеткам организма [11, 24, 25].
4. Защитная. Реализуется за счет адсорбции на поверхности эритроцитов токсических веществ, ряда вирусов и микробов [11, 25], разрушения медиаторов типа ацетилхолина ацетилхолинэстеразой [25], а также наличия компонентов антиоксидантной системы [14, 17, 20].
5. Гуморальная. Регулируют адаптационные процессы в норме и при патологии [14, 25].
6. Иммунная [15, 29]. Эритроциты взаимодействуют с антителами, циркулирующими иммунными комплексами, образующимися при взаимодействии растворимых антигенов и антител, благодаря наличию на мембране Fc-рецепторов к иммуноглобулинам, компонентам комплемента С3b и С4b и большого числа поверхностных антигенов.
7. Детоксикационная. Участие в утилизации эндогенных альдегидов, образующихся в процессе перекисного окисления липидов, реакциях гликозилирования и при окислении радикалов некоторых свободных аминокислот. При этом в разных метаболических путях образуются разные по структуре и свойствам альдегиды. В этих процессах принимают участие альдегиддегидрогеназа, альдегидредуктаза и глутатион-S-трансфераза [13].
8. Участие в гемостазе [4, 12, 29]. Основными факторами, влияющими на реологические свойства крови, являются клеточный и плазменный. Клеточный фактор — это количество и состояние эритроцитов. С учётом того, что по сравнению с тромбоцитами и лейкоцитами эритроцитов гораздо больше, именно они определяют величину гематокрита и вязкость крови [6].
9. Участие в метаболизме катехоламинов, ацетилхолина, иммунных комплексов и ряда лекарственных веществ [23].
10. Регуляция сосудистого тонуса [37].
11. Эритроциты несут в себе групповые признаки крови (агглютиногены) и детерминанты Rh [2].
Заключение
Подводя итоги изложенному, можно сделать вывод, что эритроцитарные клетки, выполняя множество разнообразных функций, имеют важное значение для поддержания жизни. Во многом это обеспечивается структурой и различными свойствами красных кровяных телец. Знание строения эритроцита (в норме и при патологии) позволяет понять механизмы клеточных взаимодействий, происходящих в организме, а также улучшить диагностику заболеваний и разработать оптимальный комплекс лечебных мероприятий.
Список использованных источников
1. Авторегуляция неспецифической проницаемости мембраны эритроцита / М.В. Фок, А.Р. Зарицкий, Г.А. Зарицкая, Е. В. Переведенцева. – М.: Наука, 1999. – 76 с.
2. Антигены эритроцитов человека / Оловникова Н.И., Николаева Т.Л. // Гематология и трансфузиология. – 2001. - №5. – С. 37-45.
3. Ашихмин Я. Анализ крови / Ашихмин Я. – 2016. – (https://postnauka.ru/faq/69175)
4. Ашкинази И.Я. Эритроцит и внутреннее тромбопластинообразование / И.Я. Ашкинази. – Л.: Наука, 1977. – 155 с.
5. Биохимия. Учебник для вузов, Под ред. Е.С. Северина. – ГЭОТАР-МЕД, 2003. – С. 659-660.
6. Боровская М.К. Структурно-функциональная характеристика мембраны эритроцита и её изменения при патологиях разного генеза / Боровская М.К., Кузнецова Э.Э., Горохова В.Г., Корякина Л.Б., Курильская Т.Е., Пивоваров Ю.И. // Бюллетень ВСНЦ СО РАМН. – 2010. - №3 (73). – С. 334-351.
7. Быкова И.А. Морфологические особенности эритроцитов периферической крови в норме и патологии (световая микроскопия) / И.А. Быкова // Вопросы охраны материнства и детства. – 1991. – Т 36, №6. – С.28-30.
8. Воробьев А.И. Клинико-диагностическое значение лабораторных показателей в гематологии. / А.И. Воробьев. – М.: Медицина, 2001. – С. 2.
9. Воробьев А.И. Руководство по гематологии. В 3-х томах / А.И. Воробьев. – М.: Ньюдиамед, 2005. – Т.3. – 416 с.
10. Гареев Р.А. Фундаментальные и прикладные аспекты адсорбционно-транспортной функции эритроцитов / Р.А. Гареев // 5 съезд физиологов Казахстана, Караганда, 2003 г.: тез. докл. – С. 75-79.
11. Гареев Р.А., Смагулова З.Ш., Макарушко С.Г. и др. Адсорбция белка, глюкозы и холестерина на эритроцитах при действии адаптивных гормонов // Научные труды I съезда физиологов СНГ. – Москва. – 2005. – С.15.
12. Григорьев Г.И. Свертывающая активность и кислотная резистентность повреждённых и неповреждённых эритроцитов при различных видах внутрисосудистого свертывания крови / Г.И. Григорьев // Вопросы охраны материнства и детства. – 1991. – Т.36, №4. – С. 13-15.
13. Давыдов В. В. Метаболизм эндогенных альдегидов: участие в реализации повреждающего действия оксидативного стресса и его возрастные аспекты / В. В. Давыдов, А. И. Божков // Биомед. хим. - 2003. - Т.49, н 4. - С. 374-387.
14. Клетки костного мозга и периферической крови / Гаврилов О.К., Козинец Г.И., Черняк Н.Б. – М.: Медицина, 1985. – 286 с.
15. Козинец Г.И. Клетки крови и костного мозга / Г.И. Козинец. – М.: МИА, 2004. – 203 с.
16. Кулапина О.И. Показатель проницаемости мембран эритроцитов при тонзиллярной патологии / Кулапина О.И., Киричук В.Ф., Зайцева И.А., Утц И.А. // Клиническая лабораторная диагностика. – 2006. - № 6. – С. 53-55.
17. Молчанова Т.П. Основы молекулярной организации белков мембраны эритроцитов и их дефекты, приводящие к гемолитическим анемиям / Т.П. Молчанова // Гематология и трансфузиология. – 1989. - №7. – С.32-41.
18. Муравьёв А.В. Методы изучения деформируемости эритроцитов в эксперименте и клинике / Муравьёв А.В., Тихомирова И.А., Муравьёв А.А., Булаева С.В., Маймистова А.А. // Клиническая лабораторная диагностика. – 2010. – №1. – C. 28.
19. Реология крови / Левтов В.А., Регирер С.А., Шадрина Н.Х. – М.: Медицина, 1982. – 270 с.
20. Ройтман Е.В. Изменение реологических свойств крови и осмотической резистентности эритроцитов при активации свободнорадикальных процессов / Ройтман Е.В., Дементьева И.И., Азизова О.А. и др. // Клиническая лабораторная диагностика. – 2001. - №3. – С.42-43.
21. Роль холестерина в структурной организации мембран эритроцитов / Шевченко О.Г. // Вестник Института биологии Коми НЦ УрО РАН / №6 (152). – 2010. – С. 12.
22. Рыскельдиев Н.А. Показатели абсорбционно-транспортной функции эритроцитов в остром периоде черепно-мозговой травмы / Рыскельдиев Н.А. // Нейрохирургия и неврология Казахстана. – 2011. - № 1(22). – С. 6-7.
23. Связь пропранолола с белками плазмы и эритроцитами / Е. Т. Гнеушев, Т. Ш. Мамедов, И. А. Гнеушева и др. // Фарм. и токсик. - 1991. - Т. 51 - №1. - С. 55-57.
24. Смирнов И.Ю., Чирикова О.А. Роль структурных элементов мембран эритроцитов в процессах адсорбции белков плазмы // Мат. междунар. конф. «Гемореалогия в микро- и макроциркуляции». – Ярославль. – 2005. – С. 202.
25. Структура и функции эритроцитарных мембран / Черницкий Е.А., Воробей А.В. – Минск: Наука и техника, 1981. – 216 с.
26. Транспорт кислорода через мембрану эритроцитов / Евстифеев А.И., Котов Н.В. – Казань: Изд. Казан. фед. ун-та, 2014. – С. 8.
27. Трошкина Н.А. Эритроцит: строение и функции его мембраны / Трошкина Н.А., Циркин В.И., Дворянский С.А. // Вятский медицинский вестник. – 2007. - №2-3. - С. 32-37.
28. Физиология и патофизиология эритроцита / Новицкий В.В., Рязанцева Н.В., Степовая Е.А. – Томск: Изд-во Том. ун-та, 2004. – 202 с.
29. Физиология системы гемостаза / Балуда В.П., Балуда М.В., Деянов Н.И. с соавт. – М.: Медицина, 1995. – 243 с.
30. Физиология человека. В 3-х томах. / Р. Шмидт, Г. Тевс. – М.: Мир, 1996. – С. 423-425.
31. Функциональная система транспорта кислорода: фундаментальные и клинические аспекты / Зинчук В.В., Максимович Н.А., Борисюк М.В. – Гродно: ГГМУ, 2003. – 236 с.
32. Шакиров Д.Ф. Исследование кислотной и осмотической резистентности эритроцитов у рабочих нефтехимического производства / Шакиров Д.Ф., Самсонов В.М., Кудрявцев В.П., Гильманов А. Ж. // Клиническая лабораторная диагностика. – 2003. - № 7. – С. 21.
33. Шмаров Д.А. Соотношение цитометрических показателей эритроцитов периферической крови и крови плода при нормальной беременности и железодефицитной анемии (ЖДА) / Шмаров Д.А, Точенов А.В., Балбулян А.К., Козинец Г.И. // Клиническая лабораторная диагностика. – 2010. - № 10. – С. 17.
34. Эритропоэз (https://ru.wikipedia.org/wiki/Эритропоэз)
35. Эритроциты крови как объект исследования метаболизма / Ю.С. Краморенко, О.В. Машнина. – (www.eyeinst.kz/docs/journal/12_39_21.doc)
36. Amount of cholesterol in host membrane affects erythrocyte invasion and replication by Babesia bovis / K. Okubo, N. Yokoyama, N. Takabatake et al. // Parasitol., 2007. – Vol. 134, №5. – P. 625-630.
37. The erythrocyte as regulator of vascular tone / M. Ellswort, T. Forrester, C. Ellis, H. Dietrich // Amer. J. Physiol. - 1995. - Vol. 269, N 6, Pt. 2. - P. 312-324.