реф по зоо. Гипотезы происхождения многоклеточных
 Скачать 186.25 Kb. Скачать 186.25 Kb.
|

|
Министерство сельского хозяйства ФГБОУ ВО «Санкт-Петербургский государственный университет ветеринарной медицины» Кафедра биологии, экологии и гистологии РЕФЕРАТ на тему: «Гипотезы происхождения многоклеточных» Работу выполнил: студент 1 курса 11 группы очного отделения Зубкова Татьяна Александровна Проверил: профессор, доктор биологических наук Чумасов Евгений Иванович Санкт-Петербург, 2022 г. СодержаниеВведение 2 Основная часть 4 Глава 1. Происхождение многоклеточных животных 4 Глава 2. Современные взгляды на происхождение многоклеточных животных 11 Заключение 18 Список литературы 20 ВведениеПроблема происхождения многоклеточных представляет интерес не только для зоологии, но и имеет большое общебиологическое значение. Многоклеточность является той морфоанатомической основой, на которой формируется огромное разнообразие структурных плоскостей, жизненных форм и эволюционных потенциалов. Поэтому знание путей и причин формирования многоклеточности у животных является ключом к пониманию многих важных общезоологических и биологических проблем. Ввиду своей исключительной важности проблема происхождения многоклеточных животных издавна привлекала внимание исследователей. На этот счет было выдвинуто много гипотез, большинство из которых сейчас представляют исторический интерес как любопытные примеры формирования зоологической мысли. Все эти гипотезы сгруппированы в четыре категории. Основная частьГлава 1. Происхождение многоклеточных животныхПервая группа состоит из гипотез, предполагающих независимое происхождение простейших и многоклеточных животных. К ним относятся представления С. Авернцева (1910) и А. А. Заварзина (1945). По мнению этих авторов, даже на заре существования жизни на Земле первичное живое вещество (первичная слизь), еще не имевшее клеточной структуры, в одном случае приобрело организацию простейших, в другом сразу же многоклеточная структура. Такие предположения противоречат как фундаментальному общему биологическому обобщению клеточной теории, так и сравнительным цитологическим данным, которые указывают на исключительное сходство тонких клеточных структур простейших и метазоа, которые вряд ли могли возникнуть независимо. Вторая группа представлена гипотезами, которые выводят многоклеточные непосредственно из отдельных простейших. В частности, Г. Иеринг (1877), А. А. Тихомиров (1887), И. Хаджи (1944), О. Стейнбок (1963) и другие авторы. Суть этих гипотез состоит в том, что многоклеточные животные эволюционировали от крупных, высокоразвитых и сложно организованных простейших путем так называемой целлюлизации, то есть одновременного разделения простейшего тела на множество специализированных клеток. Такое предположение, несмотря на всю его фантастичность с современной точки зрения, имеет определенные сравнительные анатомические и эмбриологические основания. Таким образом, некоторые инфузории с точки зрения сложности организации, по крайней мере, не уступают нижним многоклеточным, таким как некишечная турбеллярия. Гипотезы о клеточности основаны на том факте, что узкоспециализированные структуры инфузорий породили специализированные ткани и органы многоклеточных животных. Эмбриологическим основанием гипотезы о клеточности является поверхностное измельчение яиц членистоногих. При таком типе фрагментации деление ядра сначала не сопровождается делением цитоплазмы. Границы ячейки появляются одновременно и относительно поздно. Гипотезы о клеточности подвергли резкой критике В. А. Догеля, В. Н. Беклемишева, А. А. Захваткина, А. В. Иванова, О. М. Иванова-Казас и другие крупные зоологи. Короче говоря, суть этой критики заключается в следующем. Во-первых, его авторы указали на несостоятельность эмбриологического аргумента. Дело в том, что членистоногие это животные, которые далеки от происхождения многоклеточности и поэтому вряд ли могут сохранить примитивные формы развития. Сокращение членистоногого яйца без сомнения результат обширной специализации. Нижние многоклеточные имеют совершенно иной ход онтогенеза. Кроме того, исходя из гипотезы о клеточности, в онтогенезе многоклеточного животного все ткани должны были дифференцироваться сразу после фрагментации синцитиального и прямо на месте. В действительности в ходе индивидуального развития многоклеточных организмов (во время гаструляции и органогенеза) наблюдается последовательная дифференциация и значительные движения клеточных масс. Во-вторых, высокоорганизованные простейшие слишком специализированные существа, чтобы давать начало животным с организацией совершенно иного типа. Такое предположение противоречит одному из фундаментальных законов эволюции, который гласит, что источники эволюционно молодой группы организмов всегда не являются наиболее совершенным представителем эволюционных предшественников. В-третьих, гипотезы целлюлизации не имеют экологического обоснования. В связи с этим разделение тела простейших на клетки кажется необоснованным. Третья группа гипотез выводит многоклеточные из колониальных простейших. Среди них исторически первой была гипотеза желудка известного немецкого зоолога Эрнста Геккеля (1874), которая в течение длительного периода завоевала большую популярность среди специалистов. Э. Геккель обосновал эту гипотезу тем, что все многоклеточные животные в своем развитии обязательно проходят двухслойную стадию гаструл. (рисунок 1) Основываясь на биогенетическом законе Геккеля-Мюллера (онтогенез является кратким повторением филогенеза), Геккель предположил, что каждый этап индивидуального развития многоклеточного животного повторяет (повторяет) соответствующую стадию формы предков. Таким образом, стадия зиготы в филогенезе соответствует стадии одноклеточного организма, стадия morula1 (поздняя стадия дробления в виде плотного зачатка) соответствует морю колонии простейших в форме шаровидных кластеры, на стадии бластулы колония простейших в виде полого шара, похожего на современный Volvox взрыв. Индентирование (инвагинация) части стенки сферической колонии, по словам Геккеля, привело к образованию двухслойного животного гастреа, соответствующего стадии инвагинации гаструлы. Наружный слой клеток желудка (эктодерма) служил кожей, внутренний слой (энтодерма) играл роль кишечника. Бластопор действовал как отверстие рта. Среди современных Metazoa наиболее близки к гастриту Геккеля самые примитивные представители кишечнополостных, которым Геккель положил начало всему филогенетическому древу многоклеточных животных. 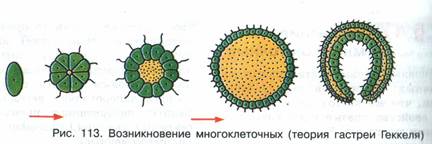 Рисунок 1. Происхождение многоклеточности по Геккелю Гипотеза о желудке Геккеля имела большое историческое значение, способствуя утверждению эволюционной идеи и краху «теории типов» Ж. Кювье. Однако она не была свободна от ряда фундаментальных недостатков. К ним относится, прежде всего, отсутствие какого-либо четкого экологического и физиологического обоснования процесса инвагинации. Гипотеза о желудке Геккеля не осталась одна. Повинуясь зову моды, многие ученые предложили оригинальные гипотезы о колониальном происхождении многоклеточных животных. Среди них я упомяну Ланкастера с его «теорией планулы» (1877 г.) и Бючли с «теорией плакулы» (1884 г.). В настоящее время эти представления представляют только узкоспециализированный исторический интерес, поэтому мы не будем останавливаться на них конкретно. Очень тщательная критика теории гастрита желудка была дана И. И. Мечниковым (1886). (рисунок 2) Таким образом, он убедительно продемонстрировал, что инвагинация не может быть исторически первым способом формирования двухслойной многоклеточной организации. Дело в том, что примитивному многоклеточному свойственен совершенно иной механизм гаструляции, а именно многополярная иммиграция. Инвагинация в филогенетическом аспекте появилась намного позже, в результате прогрессивной эволюции многоклеточного онтогенеза. Кроме того, данные сравнительной физиологии четко указывают на вторичную природу абдоминального пищеварения, которой предшествовало внутриклеточное. Поэтому, по мнению И. И. Мечникова, вряд ли первичная многоклеточная клетка, как и желудок Геккеля, могла бы иметь кишечник и отверстие рта. 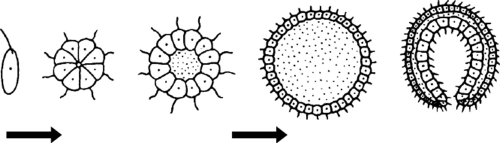 Рисунок 2. Происхождение многоклеточности по Мечникову В качестве альтернативы гипотезе Геккеля И. И. Мечников предложил оригинальную теорию, названную теорией фагоцителлы (1886). Он достаточно обоснован и в несколько модернизированном виде сохраняет свое значение в настоящее время. При разработке теории фагоцителлы И.И. Мечников исходил из следующих соображений. Предки многоклеточных животных могли быть одноклеточными с типом питания животных, то есть представителями царства простейших. Многие жгутики при определенных условиях (в частности, при захвате пищи) могут принимать амебоидную форму. Внутриклеточное пищеварение в процессе эволюции возникло раньше, чем брюшная, поэтому первичная многоклеточная едва имела кишечник, а также открывало рот. Наиболее примитивными методами гаструляции являются многополярная иммиграция и смешанное расслаивание; Филогенетические пути формирования бислойной организации должны были быть схожими. Начальным этапом развития многоклеточных животных Мечников считал сферическую колонию жгутиковых, все из которых располагались на его поверхности в один слой. Жгутики служили для плавания в колонии и способствовали улавливанию частиц пищи путем организации водоворотов (так называемый метод седиментации питания). Клетки, которые захватили частицу пищи, отбросили жгутик, приняли амебоидную форму и бросились в колонию, где они начали переваривать пищу. Закончив пищеварение и проголодавшись, они восстановили жгутик и вернулись на поверхность. Впоследствии, по мнению И. И. Мечникова, изначально однородные особи колонии были разделены на два слоя кинобласт с локомоторной функцией и фагоцитобласт с трофической функцией. Этот гипотетический организм напоминал нижние многоклеточные личинки, известные как паренхимы. Поэтому И. И. Мечников назвал это существо, согласно сложившейся зоологической традиции, паренхимеллой. Однако, учитывая тот факт, что паренхима является чисто установочной стадией и не питается сама по себе, она изменила свое мнение и предложила другое название фагоцителла (подчеркивая тем самым внутриклеточное пищеварение этого животного). Теория И. И. Мечникова, находясь в тени гипотезы Геккеля о гастрее, не получила должного признания, а затем была совершенно незаслуженно забыта. Только спустя полвека она была восстановлена в правах благодаря работам В. Н. Беклемишевой, А. В. Ивановой, А. А. Захваткиной и А. А. Заварзина. В настоящее время идеи И. И. Мечникова являются основой общепринятых представлений о происхождении многоклеточных животных, о которых пойдет речь ниже. Четвертая группа гипотез предполагает происхождение многоклеточных животных из многоклеточных растений. Франц (1919, 1924) и Харди (1953) рискнули сформулировать такие взгляды. Так, Франц предположил, что многоклеточные животные происходят от бурых водорослей, а именно от фукуса. Основное сходство, которому Франц придавал абсолютное филогенетическое значение, это сходство жизненных циклов и более или менее идентичный характер полового размножения. Концепция Харди (1953) заключается в следующем. По словам его автора, переход к многоклеточному состоянию у растений протекает легче, чем у животных, поскольку общий характер питания многоклеточного растения поглощение пищи всей поверхностью тела остается неизменным. У животного должен быть новый способ кормления, что крайне затрудняет переход в многоклеточное состояние. В противном случае многоклеточность не дает животному никаких преимуществ. Исходя из этих соображений, Харди предположил, что многоклеточные животные произошли от уже сформированных метафитов, которые преодолели вышеупомянутые трудности. Не имея минерального питания, они начали питаться небольшими организмами, как это делают современные насекомоядные растения. В результате этих соображений Харди разработал простой полипоидный метазун с везикулярной полостью и щупальцами. Гипотезы происхождения метазоя из метафитов настолько экзотичны, что нет смысла останавливаться на их критическом анализе. Глава 2. Современные взгляды на происхождение многоклеточных животныхСовременные представления о происхождении многоклеточных животных основаны на гипотезе фагоцитарлы И.И. Мечникова, несколько модернизированной и дополненной с учетом поздних открытий и идей. Специалистам хорошо известно, что любое развитие выходит на качественно новый уровень тогда и только тогда, когда возможности развития в рамках старого качества исчерпаны. Другими словами, когда развитие опирается на определенный «потолок», который невозможно преодолеть на основе предыдущей организации. Это означает, что одноклеточное существо имеет некоторые фундаментальные ограничения, которые препятствуют его улучшению. Анализ зоологического материала показал, что такие ограничения включают, прежде всего, некоторые аллометрические зависимости. Известно, что эволюция жизни на Земле идет по пути усложнения, одним из проявлений которого является так называемый филогенетический рост последовательное увеличение размеров организмов по мере их филогенетического роста. У одноклеточных организмов этот рост связан со многими факторами. Прежде всего, с необходимостью максимально простого перемещения относительно окружающей среды. Дело в том, что снабжение простейших кислородом и удаление продуктов их обмена происходит путем диффузии. В результате одноклеточное существо очень быстро создает вокруг себя «пустыню», помимо того, что загрязняется своими собственными выделениями. Поэтому для него жизненно важно изменить ситуацию, то есть перейти от поврежденной точки «А» к новой точке «В». Тем не менее, маленький организм имеет колоссальное отношение поверхности к объему и, вследствие этого, очень сильно зависит от трения о воду. Другими словами, при активном движении он испытывает чрезвычайно большое сопротивление окружающей среде. Причем это сопротивление пропорционально площади самого простого, а его двигательная сила объему. Таким образом, увеличение линейных размеров простейших, скажем, в два раза приведет к тому, что сопротивление плаванию увеличится в четыре раза, а мощность увеличится в восемь раз. В результате наблюдается тенденция к увеличению размеров простейших как эволюционный ответ на необходимость энергичного плавания. 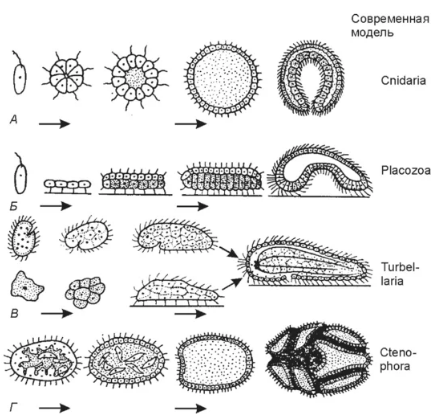 Рисунок 3. Современная гипотеза происхождения Еще одной причиной увеличения размеров является создание в вашем организме запаса питательных веществ и резервной биомассы, что делает их относительно независимыми от колебаний жизненных ресурсов. И, наконец, третья причина филогенетический рост простое следствие сложности организации. Когда есть много разных морфологических структур, им нужен более вместительный «контейнер». Таким образом, в ходе прогрессивной эволюции простейшие должны увеличиваться в размерах. Функционирование одноклеточного существа, как и любого другого живого организма, основано на соответствующих ответах на вызовы окружающей среде. В одноклеточных организмах такие реакции контролируются ядром. Например, вещество появилось в среде. Это вещество связывается с рецепторами, расположенными на внешней поверхности клеточной мембраны, и в результате этого взаимодействия рецептор посылает химический сигнал ядру в виде любой молекулы. Эта молекула достигает ядра и вызывает экспрессию желаемого гена. В результате клетка начинает синтезировать желаемое вещество: ответ состоялся. С увеличением размера самого простого расстояние между клеточной мембраной и ядром увеличивается. Время реакции организма на внешние сигналы также увеличивается, и оно, в конце концов, начинает безнадежно отставать, превращаясь в очень флегматичного и медленно движущегося гиганта, беззащитного в быстро меняющейся среде. Надо сказать, что дикая природа сталкивалась с этой проблемой не раз. Относительно недавний пример: появление на Земле крупных динозавров длиной от носа до кончика хвоста более 20 метров. Учитывая, что скорость нервного импульса у рептилий одного и того же порядка (30–40 метров в секунду), можно представить, как маленький, но наглый хищник соизволил поужинать с хвостом динозавра, прежде чем начал понимать, что у него что-то сзади, что-то происходит. Считается, что именно по этим причинам мозг гиганта был не больше теннисного мяча, тогда как основная масса нервной массы располагалась в сакральном отделе. Это «изобретение» уменьшило «плечо» рефлекторной дуги примерно вдвое, сократив время, необходимое динозавру, чтобы «осмыслить» события, происходящие с его хвостом. Выходом из этой ситуации была полиэнергетика: в самом простом было развито множество ядер, каждое из которых контролировало свою «провинцию» прилегающую область цитоплазмы. Однако это решение оказалось лишь полумерой, поскольку целостность полученного организма была небольшой. Простейшее было разделено на множество «автономий», и его скоординированное управление в целом было затруднено одинаковыми расстояниями между клеточной мембраной и глубинными частями клетки. В этом отношении самое простое было уподоблено огромной и неуклюжей Российской империи середины XIX века, когда заказы из столицы, которой в то время был Петербург, были переданы в отдаленные провинции с помощью конной эстафеты. Стоит ли удивляться, что в таком случае губернатор Камчатки признал Крымскую войну только через три месяца после ее начала, и даже эту новость он получил не из Санкт-Петербурга, а из английской эскадры, которая начала бомбардировать Петропавловск-Камчатский с морской налет. Таким образом, общий план простейшей структуры оказался чреват фундаментальными ограничениями, которые невозможно преодолеть в рамках одноклеточной организации. Тем не менее, основная линия эволюции простейших от первобытных неядерных диплоидных форм до полиплоидных и, далее, полиэнергетических представителей субдоминуса по праву интерпретируется зоологами как тенденция к многоклеточности. Как только возникают проблемы, которые не могут быть решены в рамках одноклеточной организации, остается только один путь сотрудничество одноклеточных лиц. Именно эти соображения лежат в основе современных представлений о возникновении многоклеточности у животных. Согласно этим представлениям, предки многоклеточных животных были довольно примитивными жгутиками, похожими на современных представителей хоанофлагеллята жгутиков воротничков. На их филогенетическую близость указывают сходства в ультратонкой структуре жгутика и кинетосом, митохондрий, составе резервных питательных веществ, а также наличие воротниковых клеток или клеток с зачатками воротника у некоторых многоклеточных животных. Кроме того, современные Choanophlagellata проявляют ярко выраженную тенденцию к образованию различных колоний. Первым шагом к многоклеточности было объединение жгутиконосцев с одним воротником в простую колонию типа Sphaeroeca сферический агрегат клеток, ориентированных наружу их жгутиками. Клетки колонии были полностью идентичны в морфологическом и функциональном отношении. Максимум, на что такая колония могла бы быть способна с точки зрения дифференцировки клеток, это появление морфологического градиента, как в случае современного Volvox на его функционально переднем полюсе клетки меньше и постепенно увеличиваются к функционально заднему полюсу. Поверхность сферической колонии и, следовательно, ее трение о воду намного меньше, чем общая поверхность составляющих ее особей, а локомоторная мощность колонии равна сумме двигательных способностей особей. Таким образом, сотрудничество простейших увеличило эффективность плавания. Сначала такая колония, по-видимому, размножалась только бесполым путем, распадаясь на отдельные ячейки, каждая из которых дала начало новой колонии (как в случае с современной Sphaeroeca). Чтобы колония развивалась в целом, должна была произойти первая дифференцировка клеток в генитальные и соматические. Точнее, в цикле развития наследственной формы должно было появиться новое поколение, представленное сексуальными особями, подобно тому, как это наблюдается в колониях современного Volvox или Proterospongia. Специализированные половые клетки спасли колонию от постоянного разрушения, поскольку они взяли на себя функцию размножения. Колония получила возможность развиваться как единое целое, ее интеграция могла бы усиливаться из поколения в поколение и подчиняться индивидуальности отдельных людей. Второй этап необязательная дифференциация колонии на функциональные группы клеток. Причиной этой стадии является продолжающееся увеличение размера колонии, благодаря которому составляющие ее клетки рассредоточены по периферии, и внутри желатиновой массы образуется свободное пространство. Поскольку жгутиковые клетки были расположены на поверхности колонии, локомоторная мощность стала пропорциональной площади, и дальнейшее улучшение локомоции из-за простого увеличения размера было невозможно в этом отношении эволюция зашла в тупик. Но члены колонии получили возможность чередовать разные фазы деятельности, оптимизируя выполнение определенной функции по очереди. Таким образом, расположенные снаружи клетки выполняли двигательную функцию в интересах всей колонии и питались одна за другой, отфильтровывая частицы пищи из воды, каждая для себя. «Загрузившись» в пищу, клетки потеряли жгутик, приобрели амебоидную форму и вошли в колонию, где сосредоточились на переваривании пищи. Снова проголодавшись, клетки вернулись на поверхность, восстановили жгутик, и все началось заново. Третий этап. Эта физиологическая изоляция клеточных слоев была важной предпосылкой для постоянной морфологической дифференциации колонии. В итоге клеточная масса колонии была разделена на два слоя, каждый из которых специализировался на выполнении определенных функций. Клетки внешнего слоя кинобласты взяли на себя функцию локомоции и, частично, захвата пищи (с последующим ее переносом в клетки внутреннего слоя). Клетки внутреннего слоя фагоцитобласт взяли на себя трофическую функцию захвата пищи с поверхности колонии и ее переваривания (с последующей передачей легкоусвояемых продуктов расщепления клеткам кинобласта). Таким образом, клетки больше не должны были заменять друг друга в своем движении от поверхности колонии внутрь и назад, изменяя при этом внешний вид жгутика на форму амебы и форму амебы на внешний вид жгутика. Таким образом, постоянная дифференциация членов колонии на два клеточных слоя позволила сэкономить время и жизненные ресурсы членов колонии, которые оказались эволюционно выгодными и укоренились генетически. Так возникло первое первичное многоклеточное животное (прометазоа), названное ранней фагоцителлой или фагоцителлой. Четвертый этап появление первого настоящего многоклеточного животного Eumetazoa. Его суть заключалась в эпителизации кинобластов, что повлекло за собой ряд важных эволюционных последствий. Сама эпителизация была вызвана, прежде всего, необходимостью увеличения силы межклеточных связей достаточно большого и активно плавающего существа. Таким образом, клетки кинобласта были надежно «сшиты» друг с другом и образовали самую первую ткань эктодерму. ЗаключениеЭто означало следующие преобразования. При эпителизации кинобластов часть клеток специализируется на сенсорной и стимулирующей функциях. Так появились первые чувствительные нервные элементы, образующие первичное нервное сплетение или нервную систему диффузного типа в эктодерме. Чувствительные элементы сосредоточены на аборальном полюсе, где они образуют теменную пластинку. Ведь фагоциты очень рано образовали очаг, из которого развился первичный мозг (возможно, связанный со статоцистой). Благодаря этому интеграция фагоцителлы по всему организму резко возросла. Клетки фагоцитобластов больше не могли совать свою псевдоподию между клетками жгутика, поэтому в эктодерме появилось отверстие бластопор или первичный рот, через которое клетки фагоцитобласта могли захватывать частицы пищи. Открытие рта появилось на функционально заднем полюсе, так как при плавании фагоцителлы, по гидродинамическим причинам, частицы пищи были сосредоточены там. Последнее доказано лабораторными экспериментами с личинками некоторых низкоклеточных животных, рекапитулирующих позднюю фагоцителлу: когда частицы туши добавляются в воду, все они появляются в области заднего полюса личинки, где они фагоцитируются клетками пищеварительная паренхима. С появлением орального отверстия функционально задний полюс стал морфологически задним и назван оральным (или вегетативным). В соответствии с этим противоположный полюс назывался аборальным (или животным), а ось, соединяющая их, называлась главной главной осью тела, очень важной координатой, с которой морфоанатомический анализ коррелирует с топографическим положением органов и частей всех многоклеточных животных. Таким образом, появилось существо с радиальной гетерополярной симметрией основной формой симметрии истинных многоклеточных организмов. Это существо было названо поздней фагоцителлой, или фагоцителлой. Именно оно лежит в основе филогенетического дерева всех настоящих многоклеточных животных Eumetazoa. Список литературы1. Левушкин С.И., Шилов И.А. - Общая зоология: Учебн. для студ. биол. спец. вузов -- М.: Высшая школа, 1994, 432 с. 2. Иорданский Н.Н. - Эволюция жизни -- М.: Академия, 2001, 425 с. 3. Догель В.А. - Зоология беспозвоночных: Учебник для ун-тов\ Под ред. Проф. Полянского Ю.И. - 7-е изд., перераб. и доп. - М.: Высшая школа, 1981, 606 с. 4. Иванов А.В. - Происхождение многоклеточных животных. Филогенетические очерки -- Л.: Наука, Ленингр. Отд., 1968, 287 с. 5. http\www.allbiology.ru |