Синапсы. Httpsmedfsh ruteoriyateoriyaponormalnoyfiziologiilektsiiponormalnojfiziologiisinapsfiziologiyamyshechnyhvolokon Физиология химических синапсов что такое синапс, его определение, виды синапсов, механизмы функционирования химических синапсов
 Скачать 1.08 Mb. Скачать 1.08 Mb.
|

|
https://medfsh.ru/teoriya/teoriya-po-normalnoy-fiziologii/lektsii-po-normalnoj-fiziologii/sinaps-fiziologiya-myshechnyh-volokon Физиология химических синапсов: что такое синапс, его определение, виды синапсов, механизмы функционирования химических синапсов. Эти механизмы начинаются с экзоцитоза, далее идёт выброс нейромедиаторов, и заканчиваются механизмы рецепторами (они могут быть ионотропными и метаботропными). + на лекции он приводил 2 примера рецепторов: - глутаматергические рецепторы; - ГАМК - рецепторы (их тоже разобрать). 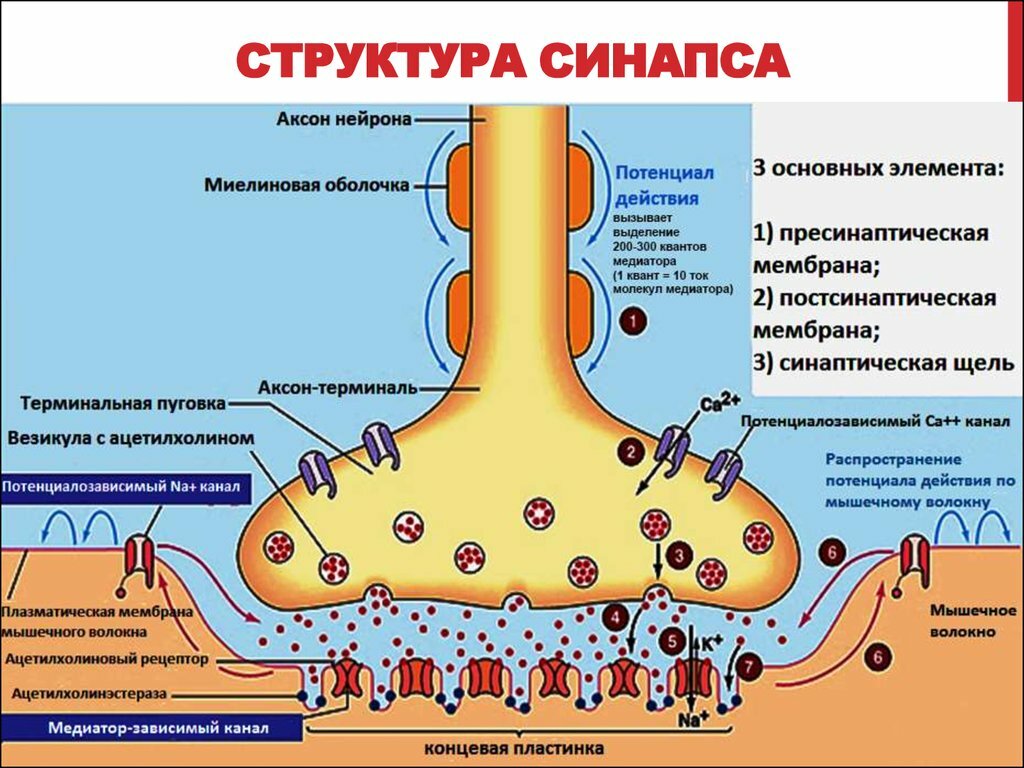 Синапс - это специализированная структура, которая обеспечивает передачу возбуждения с одной возбудимой структуры на другую. Термин «синапс» введён Ч. Шеррингтоном и означает «сведение», «соединение», «застёжка». Синапс - это структурно-функциональное образование, обеспечивающее переход возбуждения или торможения с окончания нервного волокна на иннервирующую клетку. Cтруктура синапса: 1) пресинаптическая мембрана (электрогенная мембрана в терминале аксона, образует синапс на мышечной клетке); 2) постсинаптическая мембрана (электрогенная мембрана иннервируемой клетки, на которой образован синапс); 3) синаптическая щель (пространство между пресинаптической и постсинаптической мембраной, заполнена жидкостью, которая по составу напоминает плазму крови). Существует несколько классификаций синапсов. 1. По локализации: 1) центральные синапсы; 2) периферические синапсы. Центральные синапсы лежат в пределах центральной нервной системы, а также находятся в ганглиях вегетативной нервной системы. Центральные синапсы – это контакты между двумя нервными клетками, причём эти контакты неоднородны и в зависимости от того, на какой структуре первый нейрон образует синапс со вторым нейроном, различают: 1) аксосоматический, образованный аксоном одного нейрона и телом другого нейрона; 2) аксодендритный, образованный аксоном одного нейрона и дендритом другого; 3) аксоаксональный (аксон первого нейрона образует синапс на аксоне второго нейрона); 4) дендродентритный (дендрит первого нейрона образует синапс на дендрите второго нейрона). Различают несколько видов периферических синапсов: 1) мионевральный (нервно-мышечный), образованный аксоном мотонейрона и мышечной клеткой; 2) нервно-эпителиальный, образованный аксоном нейрона и секреторной клеткой. 2. Функциональная классификация синапсов: 1) возбуждающие синапсы; 2) тормозящие синапсы. 3. По механизмам передачи возбуждения в синапсах: 1) химические; 2) электрические. Особенность химических синапсов заключается в том, что передача возбуждения осуществляется при помощи особой группы химических веществ – медиаторов. Различают несколько видов химических синапсов: 1) холинэргические. В них происходит передача возбуждения при помощи ацетилхолина; 2) адренэргические. В них происходит передача возбуждения при помощи трёх катехоламинов; 3) дофаминэргические. В них происходит передача возбуждения при помощи дофамина; 4) гистаминэргические. В них происходит передача возбуждения при помощи гистамина; 5) ГАМКэргические. В них происходит передача возбуждения при помощи гаммааминомасляной кислоты, т. е. развивается процесс торможения. Особенность электрических синапсов заключается в том, что передача возбуждения осуществляется при помощи электрического тока. Таких синапсов в организме обнаружено мало. Синапсы имеют ряд физиологических свойств: 1) клапанное свойство синапсов, т. е. способность передавать возбуждение только в одном направлении с пресинаптической мембраны на постсинаптическую; 2) свойство синаптической задержки, связанное с тем, что скорость передачи возбуждения снижается; 3) свойство потенциации (каждый последующий импульс будет проводиться с меньшей постсинаптической задержкой). Это связано с тем, что на пресинаптической и постсинаптической мембране остаётся медиатор от проведения предыдущего импульса; 4) низкая лабильность синапса (100–150 имульсов в секунду). Механизм передачи возбуждения в химических возбуждающих синапсах. В синапсах с химической передачей возбуждение передается с помощью медиаторов (посредников). Медиаторы - это химические вещества, которые обеспечивают передачу возбуждения в синапсах. Медиаторы в зависимости от их природы делятся на несколько групп: * моноамины (ацетилхолин, дофамин, норадреналин, серотонин и др.); * аминокислоты (гамма-аминомаслянная кислота - ГАМК, глутаминовая кислота, глицин и др.); * нейропептиды (вещество Р, эндорфины, нейротензин, АКТГ, ангиотензин, вазопрессин, соматостатин и др.). Медиатор в молекулярном виде находится в пузырьках пресинаптического утолщения (синаптической бляшке), куда он поступает: * из околоядерной области нейрона с помощью быстрого аксонального транспорта (аксотока); * за счет синтеза медиатора, протекающего в синаптических терминалях из продуктов его расщепления; * за счет обратного захвата медиатора из синаптической щели в неизменном виде. Когда по аксону к его терминалям приходит возбуждение, пресинаптическаямембрана деполяризуется, что сопровождается поступлением ионов кальция из внеклеточной жидкости внутрь нервного окончания. Поступившие ионы кальция активируют перемещение синаптических пузырьков к пресинаптичской мембране, их соприкосновение и разрушение (лизис) их мембран с выходом медиатора в синаптическую щель. В ней медиатор диффундирует к субсинаптической мембране, на которой находятся его рецепторы. Взаимодействие медиатора с рецепторами приводит к открытию преимущественно каналов для ионов натрия. Это приводит к деполяризации субсинаптической мембраны и возникновению так называемого возбуждающего постсинаптического потенциала (ВПСП). В нервно-мышечном синапсе ВПСП называется потенциалом концевой пластинки (ПКП). Между деполяризованной субсинаптической мембраной и соседними с ней участками постсинаптическои мембраны возникают местные токи, которые деполяризуют мембрану. Когда они деполяризуют мембрану до критического уровня, в постсинаптической мембране мышечного волокна возникает потенциал действия, который распространяется по мембранам мышечного волокна и вызывает его сокращение. Химические тормозные синапсы. Эти синапсы по механизму передачи возбуждения сходны с синапсами возбуждающего действия. В тормозных синапсах медиатор (например, глицин) взаимодействует с рецепторами субсинаптической мембраны и открывает в ней хлорные каналы, это приводит к движению ионов хлора по концентрационному градиенту внутрь клетки и развитию гиперполяризации на субсинаптической мембране. Возникает так называемый тормозной постсинаптический Потенциал (ТПСП). Ранее полагали, что каждому медиатору соответствует специфическая реакция постсинаптической клетки - возбуждение или торможение в той или иной форме. В настоящее время установлено, что одному медиатору чаше всего соответствует .не один, а несколько, различных рецепторов. Например, ацетилхолин в нервно-мышечных синапсах скелетных мышц действует на Н-холинорецепторы (чувствительные к никотину), которые открывают широкие каналы для натрия (и калия), что порождает ВПСП (ПКП). В ваго-сердечных синапсах тот же ацетилхолин действует на М-холинорецепторы (чувствительные к мускарину), открывающие селективные каналы для ионов калия, поэтому здесь генерируется тормозной постсинаптический потенциал (ТПСП). Следовательно, возбуждающий или тормозной характер действия медиатора определяется свойствами субсинаптической мембраны (точнее, видом рецептора), а не самого медиатора. Физиологические свойства химических синапсов. Синапсы с химической передачей возбуждения обладают рядом общих свойств: * Возбуждение через синапсы проводится только в одном направлении (односторонне). Это обусловлено строением синапса: медиатор выделяется только из пресинаптического утолщения и взаимодействует с рецепторами субсинаптической мембраны; * передача возбуждения через синапсы осуществляется медленнее, чем по -нервному волокну - синаптическая задержка; * передача возбуждения осуществляется с помощью специальных химических посредников - медиаторов; * в синапсах происходит трансформация ритма возбуждения; * синапсы обладают низкой лабильностью; * синапсы обладают высокой утомляемостью; * синапсы обладают высокой чувствительностью к химическим (в том числе и фармакологическим) веществам. Электрические синапсы возбуждающего действия. Кроме синапсов с химической передачей возбуждения преимущественно в центральной нервной системе (ЦНС) встречаются синапсы с электрической передачей. Возбуждающим электрическим синапсам свойственны очень узкая синаптическая щель и очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран, что обеспечивает эффективное прохождение локальных электрических токов. Низкое сопротивление, как правило, связано с наличием поперечных каналов, пересекающих обе мембраны, т. е. идущих из клетки в клетку (щелевой контакт). Каналы образуются белковыми молекулами (полумолекулами) каждой из контактирующих мембран, которые соединяются комплементарно. Эта структура легко проходима для электрического тока. Схема передачи возбуждения в электрическом синапсе: ток, вызванный пресинаптическим потенциалом действия, раздражает постсинаптическую мембрану, где возникает ВПСП и потенциал действия. Поперечные каналы объединяют клетки не только электрически, но и химически, так как они проходимы для многих низкомолекулярных соединений. Поэтому возбуждающие электрические синапсы с поперечными каналами формируются, как правило, между клетками одного типа (например, между клетками сердечной мышцы). Общими свойствами возбуждающих электрических синапсов являются: * быстродействие, (значительно превосходит таковое в химических синапсах); * слабость следовых эффектов при передаче возбуждения (в результате этого в них практически невозможна суммация последовательных сигналов); * высокая надёжность - передачи возбуждения. Возбуждающие электрические синапсы могут возникать при благоприятных условиях и исчезать при неблагоприятных. Например, при повреждении одной из контактирующих клеток её электрические синапсы с другими клетками ликвидируются. Это свойство называется пластичностью. Электрические синапсы могут быть с односторонней и двусторонней передачей возбуждения. Электрический тормозной синапс. Наряду с электрическими синапсами возбуждающего действия могут встречаться электрические тормозные синапсы. Примером такого синапса может служить синапс, который образует нервное окончание на выходном сегменте маутнеровского нейрона у рыб. Тормозящее влияние возникает за счёт действия тока, вызванного потенциалом действия пресинаптической мембраны. Пресинаптический потенциал вызывает значительную гиперполяризацию сегмента и гиперполяризующий ток мгновенно тормозит генерацию потенциала действия в начальном сегменте аксона. +В смешанных синапсах пресинаптический потенциал действия создаёт ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны неплотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом. Экзоцитоз Экзоцитоз (экзо — наружу) — процесс, обратный эндоцитозу. Благодаря ему клетка выводит внутриклеточные продукты или непереваренные остатки, заключённые в вакуоли или пузырьки. Пузырёк подходит к цитоплазматической мембране, сливается с ней, а его содержимое выделяется в окружающую среду. Так выводятся пищеварительные ферменты, гормоны, гемицеллюлоза и др.  Таким образом, биологические мембраны как основные структурные элементы клетки служат не просто физическими границами, а представляют собой динамичные функциональные поверхности. На мембранах органелл осуществляются многочисленные биохимические процессы, такие как активное поглощение веществ, преобразование энергии, синтез АТФ и др.  Общая характеристика и классификация нейромедиаторов. Нейромодуляторы Для непрерывной передачи импульсов нужно быстро удалять медиатор из белка-рецептора, чтобы следующий квант посредника мог с ним взаимодействовать. Механизмы удаления медиатора из синаптической щели могут быть различные: или путём разрушения медиатора специфическим ферментом, находящимся в синаптической щели, или путём обратного захвата медиатора специальным переносчиком и поступления его назад в пресинаптическое нервное окончание или в глиальную клетку, или же путём разрушения медиатора под действием специфического фермента в синаптической щели, или в постсинаптическом нейроне, или глиальной клетке. Для выделения везикул с медиатором необходимо определённое время, вследствие чего задержка передачи сигнала составляет 0,2-0,5 мс. Поэтому лабильность синапса составляет не более 100-150 имп/с. При большей частоте импульсов происходит блокировка проведения сигнала. Быстрое утомление синапсов связано с исчерпанием запасов везикул с медиатором в пресинаптическом нервном окончании и необходимости определённого времени для восстановления их запасов.  Таким образом, нейромедиаторы – это вещества, образующиеся в пресинаптических нервных окончаниях, хранящиеся там в особых везикулах, выделяющиеся из нервных окончаний под действием нервного импульса в синапс, связывающиеся со специфическим рецептором на постсинаптической мембране и имеющие механизмы для быстрого удаления медиаторов из синаптической щели. В роли медиаторов выступают ацетилхолин, дофамин, норадреналин, серотонин, ГАМК, глицин и некоторые другие вещества. В зависимости от эффектов на постсинаптической мембране нейромедиаторы делят на возбуждающие и тормозные. К возбуждающим нейромедиаторам относят ацетилхолин, дофамин, серотонин, глутаминовую кислоту, норадреналин. В некоторых синапсах в качестве возбуждающего нейромедиатора могут выделяться пурины, АТФ, некоторые нейропептиды. К тормозным нейромедиаторам относят гамма-аминомасляную кислоту (ГАМК) и глицин. Критерии нейромедиаторов: 1) вещество должно синтезироваться и накапливаться в пресинаптических нервных окончаниях, из которых должно выделяться в ответ на деполяризацию; 2) выделившийся нейромедиатор должен оказывать постсинаптическое действие путём взаимодействия со специфическим постсинаптическим рецептором; 3) вещество должно или разрушаться в синаптической щели, или удаляться из неё с помощью механизма обратного захвата; 4) введение этого вещества в нервную ткань должно оказывать такое же действие, как и эндогенное вещество. Кроме медиаторов, в синапсе могут выделяться также нейромодуляторы. Нейромодуляторы – это вещества, которую прямо не изменяют состояние синаптических мембран, но влияют на интенсивность и продолжительность действия классических нейромедиаторов и таким образом влияют на активность синаптической передачи сигнала. Чаще всего в роли нейромодуляторов выступают нейропептиды. Нейропептиды – это обширная группа короткоцепочечных пептидов, которые могут выступать и как нейромедиаторы, и как нейромодуляторы. Например, энкефалины и эндорфины связываются со специфическими рецепторами (с которыми также взаимодействует морфин), при этом происходит подавление чувства боли. Другой пептид – вещество Р, подавляет ощущение боли, а также вызывает сокращение гладких мышц. Ангиотензин II – гормон местного действия, сильно влияет на кровеносные сосуды и работу ЦНС. Так же действует и вазоактивный кишечный пептид (ВИП). Простагландины действуют как локальные химические агенты, так как они очень быстро инактивируются. Модулируя синаптическую передачу, они влияют на многие физиологические процессы, например, меняют секрецию медиаторов, работу аденилатциклаз. Рецепторы к нейромедиаторам располагаются на мембранах нейронов или клеток-мишеней (мышечные или железистые клетки). Их локализация может быть и на постсинаптических, и на пресинаптических мембранах. На пресинаптических мембранах чаще располагаются так называемые ауторецепторы, которые регулируют выделение этого же медиатора из пресинаптического окончания. Но есть и гетероауторецепторы, которые также регулируют выделение медиатора, но в этих рецепторах выделение одного медиатора регулирует другой медиатор или нейромодулятор. Большинство рецепторов – это мембраносвязанные олигомерные белки, которые связывают лиганд (нейромедиатор) с высоким сродством и высокой селективностью. В результате этого взаимодействия запускается каскад внутриклеточных изменений. Рецепторы характеризуются сродством к лиганду, количеством, насыщаемостью и способностью к диссоциации рецептор-лигандного комплекса. У некоторых рецепторов обнаружены изоформы, которые различаются сродством к определённым лигандам. Эти изоформы могут находиться в одной и той же ткани. Лиганды - это вещества, избирательно взаимодействующие с данным рецептором. Если фармакологическое вещество активирует данный рецептор, оно является агонистом для него, а если снижает его активность – то антагонистом. Связывание лиганда с рецептором приводит к изменению конформации рецептора, вследствие чего или открываются ионные каналы, или запускается каскад реакций, приводящих к изменениям метаболизма. Выделяют ионотропные и метаботропные рецепторы. Ионотропные рецепторы. Вследствие образования постсинаптического потенциала происходит открытие соответствующего ионного канала или сразу при действии медиатора, или через активацию G-белка. При этом рецептор или сам образует ионный канал, или связан с ним. После присоединения лиганда и активации рецептора происходит открытие канала для соответствующего иона. В результате на мембране образуется постсинаптический потенциал. Ионотропные рецепторы – это путь быстрой передачи сигнала и образования ПСП без изменения процессов метаболизма в клетке. Метаботропные рецепторы. Это более сложный путь передачи сигнала. При этом после связывания лиганда с рецептором происходит активация каскада фосфорилирование-дефосфорилирование. Это осуществляется или прямо, или через вторичные посредники, например, через тирозинкиназу, или через цАМФ, или цГМФ, или инозитолтрифосфат, или диацилглицерол, или за счёт увеличения внутриклеточного кальция, что в результате приводит к активации протеинкиназ. Фосфорилирование чаще всего включает в себя активацию цАМФ-зависимой или диацилглицерол-зависимой протеинкиназы. Эти эффекты развиваются более медленно и длятся более долго. Сродство рецептора к соответствующему нейромедиатору может меняться так же, как и к гормонам, например, за счёт аллостерических изменений рецептора или других механизмов. Поэтому сейчас рецепторы обозначают как мобильные и легко изменяемые структуры. Входя в состав мембраны, белки-рецепторы могут взаимодействовать с другими мембранными белками (так называемая интернализация рецепторов). Нейромодуляторы, как и нейромедиаторы, могут влиять на число и чувствительность рецепторов. Длительное присутствие больших количеств нейромедиатора или нейромодулятора может снижать их чувствительность (даун-регуляция), а недостаток лигандов повышать их чувствительность (ап-регуляция).   N-холинорецепторы электрических органов и скелетных мышц представляют собой пентамерные структуры, образованные четырьмя типами субъединиц (α; β; γ либо ε; δ) в соотношении α2βγδ (в эмбриональных или денервированных мышцах) или α2βεδ (в иннервированных мышцах взрослых особей). Степень гомологии аминокислотных последовательностей между разными типами субъединиц достигает 40%, что позволяет предположить их общее происхождение (Numa etal., 1983). N-холинорецепторы стали классической моделью для изучения ионотропных рецепторов (хемочувствительных каналов, рецепторов-каналов). К этой группе принадлежат также рецепторы тормозных аминокислотных медиаторов (ГАМК и глицина), 5-НТ3-рецепторы и некоторые другие. Все они представляют собой пентамеры, состоящие из гомологичных субъединиц с молекулярной массой порядка 40 000—60 000 каждая. Внеклеточные домены каждой субъединицы состоят из 210 N-конце-вых аминокислот. Затем следуют 4 трансмембранных домена, причем на участок между 3-м и 4-м доменами приходится почти вся внутриклеточная часть субъединицы (рис. 9.1). 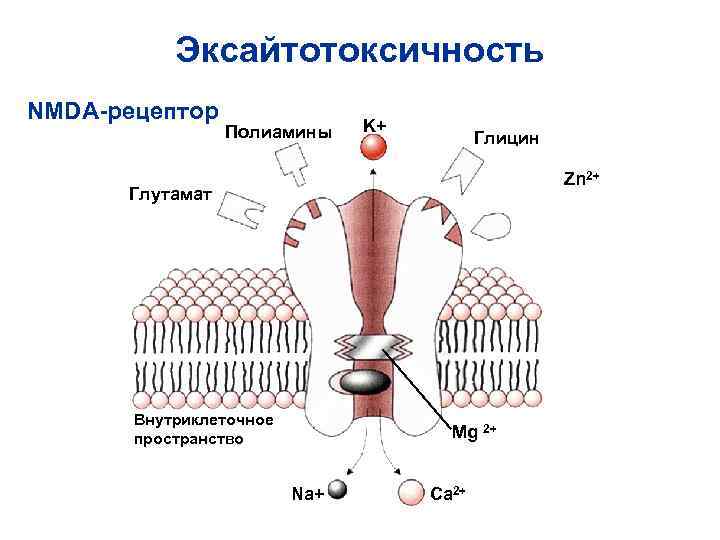 На этом рисунке представлена структура NMDA рецептора, соответствующая имеющимся знаниям.  У NMDA рецептора существует семь различных мест [сайтов] связывания. Три из них находятся снаружи поверхности клетки, два - внутри, одно внутри канала, и еще одно (место иона магния) на наружной и внутренней поверхностях. Два сайта для агонистов, обозначенных EAA (ВАК) и Gly; они соответствуют возбуждающей аминокислоте (глютамат и аспартат) и глицину. Оба сайта должны быть заняты перед тем, как канал сможет открыться и пропустить ион. Третий сайт - мишень иона цинка (Zn2+), который блокирует канал своим присутствием. Снаружи клетки в канале имеется сайт магниевого иона. Точно такой же сайт существует и внутри клетки (он может находится и внутри самого канала). Обычно ион магния занимает наружный сайт; внутренний сайт вероятно пустует из-за биологиеских условий. Внутриклеточная часть канал содержит два места связывания. Одно связывается с полиаминами (спермином и спермидином), его функции пока остаются неизвестными. Другое, непоказанное на рисунке, - это сайт фосфориляции. Ферменты могут связываться с этим сайтом увеличивая или уменьшая активность рецептора. Наконец, внутри самого канала находится сайт связывания PCP1, где связываются PCP, кетамин, MK-801 (dizocilpine), DXM и декстрорфан. Канал должен быть полностью открыт для того, чтобы эти наркотики могли войти; как только они связываются с сайтом, канал "забивается". NMDA рецепторы уникальны по нескольким причинам. В отличие от большинства других рецепторов, они требуют присутствия двух агонистов (глютамата или аспартата, и глицина) для открытия канала. Эти два агониста (Glu and Gly на рисунке) связываются в двух различных сайтах NMDA рецептора. После того, как оба агониста связались с каналом, он открывается достаточно широко для прохода калия, и рецептор работает сходным с AMPA и каинатным рецепторами образом. Это показано на Рисунке 9.  Но самая важная и уникальная характеристика NMDA рецептора - это то, что случается далее (Рисунок 10). Обычно ион магния связывается в специальном сайте у входа в канал; этот ион позволяет проходить калию, но не пропускает кальций, видимо из-за различия размеров. Эта связь образуется электростатическими силами.  Как только клетка становится достаточно активной - то есть клеточный потенциал увеличивается, и ион магния больше не может находиться на месте. Теперь в клетку может войти (и выйти, хотя этого не происходит) кальций через полностью открытый NMDA канал. Войдя внутрь, кальций вызывает серию импульсов, увеличивающих силу действия синапса. Ну и в чем тут смысл? Ну, если нейрон слегка активен, то NMDA канал может открыться частично, и ион магния не сможет покинуть своего места. Однако, если нейрон будет быстро или сильно активизирован, ион магния уйдет, в клетку войдет кальций и увеличит активацию нейрона и синаптическую силу. Этот процесс увеличения синаптической силы называется Долговременной потенциацией (Long-Term Potentiation) (ДВП (LTP)), и является одним из механизмов, с помощью которых нейроны могут изменять свою работу и "учиться". ДВП в гиппокампе вероятно отвечает за кратковременную память. Способность к обучению на самом деле может прямо зависеть от числа NMDA рецепторов в гиппокампе (где, как полагают, хранятся кратковременные воспоминания). ДВП обратима, и договременная (постоянная) память, скорее всего, хранится с помощью более постоянных изменений в экспрессии генов синаптической формы. Другой интересный аспект двойного действия NMDA рецептора - его работа как "детектора совпадений". Кальций может войти в клетку, только когда потенциал ее мембраны достаточно низок и синапс активирован; таким образом он определяет момент совпадения синаптического и межклеточного сигналов. В ассоциативных сетях (каким мы и полагаем гиппокамп) NMDA рецептор идеально сконструирован, чтобы помочь нейрону "обучиться" ассоциировать два входных сигнала. +Существует, по крайней мере, три типа NMDA рецепторов (у крысы, что, возможно, распространяется и на человека). Один вид находится в мозжечке, другой в таламусе и третий в коре головного мозга. Они слегка различаются, но возможно, что DXM оказывает другой спектр эффектов на эти виды рецепторов, чем другие NMDA антагонисты (такие как кетамин, PCP). Есть некоторые предположения, что ионный канал NMDA рецептора может (по неизвестной причине) стать "отделенным" от самого рецептора. Неконкурентный антагонизм NMDA рецепторов со стороны блокаторов открытого канала известен своим воздействием на весь мозг. NMDA блоакда вызывает усиленную секрецию дофамина в промежуточном мозге и префронтальной коре. NMDA блокада вызывает и активацию 5HT систем специфично нацеленную на 5HT1A рецептор. |