Молекулярная организация хромосом
 Скачать 94.67 Kb. Скачать 94.67 Kb.
|

|
МОЛЕКУЛЯРНАЯ ОРГАНИЗАЦИЯ ХРОМОСОМ Термин хромосома был предложен В.Вальдейером в 1888 году для обозначения интенсивно окрашенных структур, наблюдаемых в ядре во время деления клетки. В интерфазе хромосомы деконденсируются и представлены в виде хроматина. В хроматине выделяют два типа участков: эухроматин, который транскрибируется и является генетически активным, и гетерохроматин, который не может быть транскрибирован и, таким образом, генетически неактивен. В метафазе клеточного деления хромосомы достигают максимальной конденсации, что позволяет точно идентифицировать хромосомы и изучить их морфологию. Каждая хромосома состоит из одной или двух молекул ДНК (в зависимости от периода клеточного цикла), которые, связываясь с гистоновыми и негистоновыми белками, образуют нуклепротеиновые комплексы с разным уровнем конденсации: - нуклеосомный уровень- молекула ДНК с диаметром 2 нм взаимодействует с гистонами, формируя первичную нуклеосомную нить, единицей организации которой является гистоновый октамер из восьми молекул гистоновых белков (2Н2А, 2Н2В, 2НЗ и 2Н4) и ДНК; на каждый октамер приходится около двух витков спирали ДНК из «160 п.н.: в результате образуется нуклеопротеиновая полинуклеосоная нить диаметром 11 нм, а длина молекулы ДНК в результате упаковки сокращается в 6 раз; - соленоид - второй уровень упаковкигенетического материала; образуется посредством конденсации нуклеосомной нити при участий гистона HI; Диаметр нуклеосомной нити на этом уровне составляет около 30 нм, а уровень укладки достигает 40 раз; - третий уровень конденсации - петли, которые образуются следующим образом: соленоидная нить фиксируется к специальной продольной оси., образованной белками ядерного матрикса (scaffold); соленоид взаимодействует с осью в специфических сайтах (SAR) посредством специальных белков хромосомной нити (SAP); на этом уровне генетический материала конденсируется в 600 - 1000 раз; - четвертый уровень конденсации - метафазная хромосома- является результатом спирализации петель вокруг хромосомной оси после утери контакта scaffold / ядерная оболочка; один виток спирали состоит из примерно 30-ти петель, при этом обеспечивается максимальный (10 000 раз) уровень упаковки генетического материала. ОБЩАЯ ХАРАКТЕРИСТИКА ХРОМОСОМ Хромосомы представляют собой надмолекулярный уровеньорганизации генетического материала и морфологический субстрат наследственности. Основным компонентом хромосомы является ДНК, которая обеспечивает хранение, передачу и реализацию генетической информации, представленной в виде специфических полинуклеотидных последовательностей - генов. Одна хромосома содержит от нескольких сотен до нескольких тысяч структурных генов и: - представляет одну группу сцепления. - обеспечивает упорядоченное расположение генов в пространстве и времени; - обеспечивает сцепленное (совместное) наследование генов. Хромосомы удваиваются путем полуконсервативной репликации ДНК в фазе S клеточного цикла. Репликация различных участков хромосомы происходит асинхронно, но к концу фазы S полностью завершается, и все хромосомы становятся двухроматидными. Хромосомы являются очень динамичными образованиями, меняя свою форму и активность в зависимости от периода клеточного цикла. Кроме того, они отличаются гетерогенностью, благодаря чередованию различных последовательностей ДНК, среди которых: - кодирующие и некодирующие последовательности; - уникальные и повторяющиеся последовательности; - эухроматиновые и гетерохроматиновые сегменты; - участки, отличающиеся по уровню закручивания хроматиновой нити (размеры петель); - последовательности, богатые парами А=Т и G=С; - участки с разным содержанием ассоциированных белков. Все это обусловливает полиморфизм хромосом, их дифференциальную окраску и специфическое чередование полос (бэндов). Каждый индивид, каждая клетка содержит диплоидный набор хромосом, который представлен парами гомологичных хромосом. 23 пары хромосом соматической клетки человека составляют ее кариотип и обеспечивают, как количественно, так и качественно фенотип клетки и организма человека. Изменение числа или структуры хромосом приводит к серьезным нарушениям развития -хромосомным синдромам (например: 47, XX(XY),+21 - синдром Дауна; 46,XX(XY),5p- - синдром „cri-du-chat") или вызывают трансформацию анеуплоидных клеточных клонов. В настоящее время ученые-цитогенетики располагают разнообразными методами кариотипирования для выявления численных и структурных хромосомных аномалий с целью: диагностики хромосомных болезней, пренатальной диагностики и предупреждения рождения детей с хромосомными болезнями, изучения хромосомных нарушений в опухолевых клетках. МОРФОЛОГИЯ МЕТАФАЗНЫХ ХРОМОСОМ Хромосомы лучше всего изучать во время метафазы митоза, т.к. в этой фазе они: - располагаются в центре клетки, образуя метафазную пластинку; - максимально конденсированы и легко различимы с использованием световой микроскопии; - являются двухроматидными, а сестринские хроматиды соединены между собой в области центромеры, что позволяет различить их морфологию. 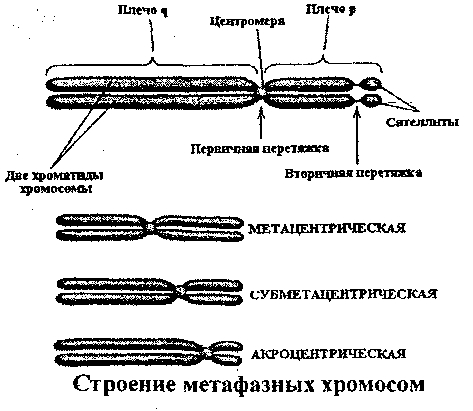 Морфологическими элементами метафазной хромосомы являются: - 2 хроматиды; - центромера; - теломеры; - вторичная перетяжка; - спутник (сателлит); - ломкие (фрагильные) участки. Хроматида представлена одной линейной молекулой ДНК, ассоциированной с гистоновыми и негистоновыми белками и максимально конденсированной. Метафазная хромосома состоит из двух сестринских хроматид, являющихся результатом репликации ДНК в фазе S и, таким образом, генетически идентичных. Хроматиды одной хромосомы соединены в области центромеры и остаются в таком состоянии до анафазы. Центромера, или первичная перетяжка, представляет собой специфический участок хромосомы из ДНК и специальных центромерных белков (CENP-A,B,C,D,E). Центромерная ДНК состоит из высокоповторяющихся последовательностей (сателлитная ДНК), практически одинаковых для всех хромосом. Положение центромеры в хромосоме постоянно и специфично для каждой хромосомы/пары гомологичных хромосом. Центромера делит хромосому на два плеча: р (проксимальное) и q (дистальное). По положению центромеры хромосомы делятся на: - метацентрические - центромера расположена посередине и плечи равные; - субметацентрические - центромера несколько смещена к одному из концов, а плечи имеют разную длину; - акроцентрические - центромера значительно смещена к концу хромосомы, из-за чего одно плечо намного короче другого. Центромеры выполняют следующие функции: - созревание кинетохоров для прикрепления хромосом к нитям веретена деления; - продольное расщепление и разделение сестринских хроматид с образованием из одной двухроматидной хромосомы двух однохроматидных хромосом; - точное и равное распределение генетического материала во время митоза, точная передача генетической информации от клетки к клетке. Теломеры представлены специфическими последовательностями ДНК на концах хромосом в комплексе со специальными белками. В состав теломерной ДНК входят: (а) тандемно и многократно повторяющиеся короткие последовательности (TTAGGG), одинаковые у всех хромосом, (b) специфические для каждой хромосомы последовательности ДНК. Теломеры выполняют следующие функции: - защищают концы хромосом от действия нуклеаз; - предотвращают слипание концов хромосом; - обеспечивают репликацию всей ДНК; - предотвращают укорачивание хромосом благодаря активности теломеразы; - контролируют процессы старения клеток и многоклеточного организма; - регулируют фиксацию хроматина к ядерной мембране в интерфазе, обеспечивая тем самым нормальную архитектуру интерфазных хромосом; - обеспечивают правильную конъюгацию гомологичных хромосом в мейозе. Вторичные перетяжки (h) представляют собой деспирализованные и слабо окрашенные участки повторяющейся ДНК; в норме они могут быть как в проксимальных плечах (р) акроцентрических хромосом 13, 14, 15, 21, 22, так и в дистальных плечах хромосом 1, 9, 16, реже 4, 6, 10 и Y. Вторичные перетяжки акроцентрических хромосом образуют область ядрышкового организатора. Длина вторичной перетяжки может варьировать в пределах нормального индивидуального полиморфизма. Сателлиты - это терминальные участки коротких плеч акроцентрических хромосом 13, 14, 15, 21, 22, отделенные вторичной перетяжкой и состоящие из конститутивного гетерохроматина; число и размеры сателлитов варьируют от индивида к индивиду. Ломкие (фрагильные) участки представляют собой деконденсированные сегменты хромосом, отличающиеся повышенной чувствительностью к действию мутагенных факторов, под влиянием которых в них легко происходят разрывы и, в результате этого, хромосомные перестройки. Фрагильные участки: - являются маркерами нормального индивидуального полиморфизма; - ассоциированы с некоторыми моногенными синдромами (например, FRAXA и семейная умственная отсталость); - могут участвовать в опухолевой прогрессии (путем инактивации генов-супрессоров). КЛАССИФИКАЦИЯ ХРОМОСОМ ЧЕЛОВЕКА Каждая соматическая клетка организма человека содержит диплоидный набор хромосом (2п=46), или 23 пары хромосом: - 22 пары, идентичные у мужчин и женщин, - аутосомы; - 1 пару отличающуюся у разных полов хромосом (XX - у женщин, XY - у мужчин) -гоносомы. Идентичные по морфологии (размеры и форма) и содержанию генов, но различные по родительскому происхождению хромосомы, называются гомологичными. В зрелых половых клетках - гаметах - содержится по одному гаплоидному набору хромосом (n=23): в яйцеклетках 22+Х, а в сперматозоидах 22+Х или 22+Y. Для идентификации хромосом используют морфологические критерии, данные авторадиографического анализа и выявляемые методами дифференциальной окраски бэнды. Морфологические критерииотражают размеры и конфигурацию хромосомы. Различают количественные (длина хромосомы, центромерный индекс) и качественные (наличие вторичной перетяжки и сателлитов) критерии классификации хромосом человека. Длина хромосомы- абсолютная длина (в микронах) или относительная длина, которая вычисляется по следующей формуле: В зависимости от длины хромосомы классифицируют на: большие, средние, мелкие. Положение центромеры. Для характеристики положения центромеры на хромосоме используют центромерный индекс, который определяют по формуле: Исходя из положения центромеры и величины центромерного индекса хромосомы человека делят на:  На основании морфологических количественных (длина и положение центромеры) и качественных (сателлиты и вторичные перетяжки) критериев хромосомы человека классифицируют на 7 групп, которые обозначают буквами латинского алфавита от А до G: - группа А (пары 1-3) - большие метацентрические хромосомы; хромосома 1 может иметь вторичную перетяжку (lqh), хромосома 2 слабо субметацентрическая; - группа В (пары 4-5) - большие субметацентрические хромосомы; - группа С (пары 6-12 и хромосома X) - субметацентрические хромосомы средних размеров; в этой группе у женщин 16 хромосом, у мужчин - 15; хромосомы 8, 9, 10 и 12 более субметацентрические, в то время как хромосомы 6, 7, 11 и X менее субметацентрические; хромосома 9 может иметь вторичную перетяжку на дистальном плече (9qh); - группа D (пары 13-15) - средние акроцентрические хромосомы; все хромосомы этой группы имеют вторичную перетяжку и сателлит на проксимальном плече; - группа Е (пары 16-18) - хромосома 16 средняя метацентрическая, может иметь вторичную перетяжку на дистальном конце; хромосомы 17 и 18 мелкие и субметацентрические; - группа F (пары 19-20)- мелкие метацентрические хромосомы; - группа G (пары 21-22 и хромосома Y) - мелкие акроцентрические хромосомы; хромосомы 21 и 22 могут иметь вторичную перетяжку и сателлит на проксимальном плече; хромосома Y не имеет сателлита; хромосомы группы G используют для определения пола: в этой группе у женщин 4 акроцентрические хромосомы (2 хр. 21 + 2 хр. 22), а у мужчин - 5 акроцентрических хромосом (2 хр. 21 +2хр. 22+Y). ИЗУЧЕНИЕ ХРОМОСОМ ЧЕЛОВЕКА Изучение хромосом предполагает определение количества, формы и структуры хромосом, идентификацию специальных показателей - реперов и анализ индивидуального полиморфизма. Современная цитогенетика человека располагает разнообразными методами, позволяющими идентифицировать численные и структурные хромосомные аномалии, выявлять хромосомные маркеры в норме и патологии. Более того, некоторые методы, основанные на технологии рекомбинантной ДНК, позволяют выявить изменения на уровне последовательностей ДНК. В зависимости от поставленной цели, хромосомы можно изучать как в интерфазе, так и во время клеточного деления. Это позволяет идентифицировать изменения в строении и числе хромосом (хромосомные аберрации, анеуплоидии), а также некоторые небольшие по размерам изменения в молекуле ДНК (микроделеции, микродупликаций). 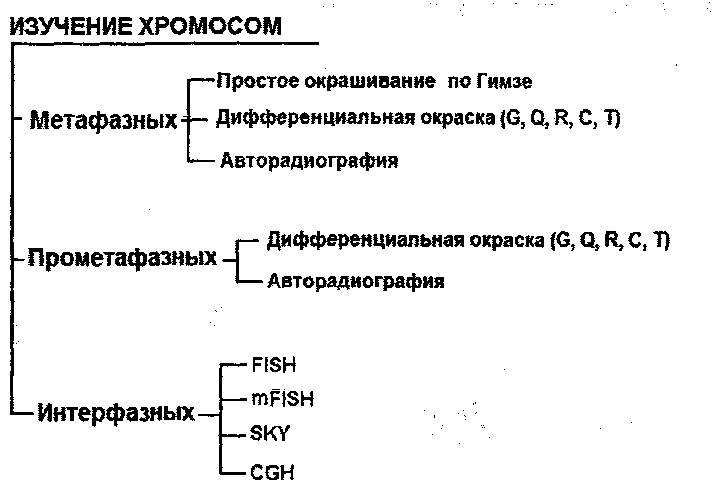 Изучение метафазных хромосом Оптимальным этапом для изучения хромосом является стадия метафазы, когда хромосомы достигают максимальной конденсации и располагаются в одной плоскости, что позволяет их идентифицировать с высокой точностью. Для изучения кариотипа требуется выполнение нескольких условий: - стимуляция клеточных делений для получения максимального количества делящихся клеток, - блокирование клеточного деления в метафазе; - гипотонизацш клеток и приготовление препарата хромосом для дальнейшего исследования под микроскопом. Для изучения хромосом можно использовать клетки из активно пролиферирующих тканей (клетки костного мозга, стенок семенников, опухолей) или клеточные культуры, которые получают путём культивирования в контролируемых условиях на специальных питательных средах клеток, выделенных из организма (клетки периферической крови*, лимфоциты Т, клетки красного костного мозга, фибробласты разного происхождения, клетки хориона, опухолевые клетки) * Техника получения хромосомных препаратов из лимфоцитов периферической крови, культивируемых в изолированных условиях является наиболее простым методом и состоит из следующих этапов: - забор венозной крови в асептических условиях; - добавление гепарина для предотвращения свертывания крови; - перенос материала во флаконы со специальной питательной средой; - стимуляция клеточных делений добавлением фитогемагглютинина; - инкубация культуры в течение 72 часов при температуре 370С. Блокирование клеточного деления на стадии метафазы достигается введением в среду колхицина или колцемида – веществ - цитостатиков, разрушающих веретено деления. Получение препаратов для микроскопического анализа включает следующие этапы: - гипотонизацю клеток, которая достигается добавлением гипотонического раствора хлорида калия; это приводит к набуханию клетки, разрыву ядерной оболочки и дисперсии хромосом; - фиксацию клеток для остановки жизнедеятельности клетки с сохранением структуры хромосом; для этого используются специальные фиксаторы, например, смесь этилового спирта и уксусной кислоты; - окрашивание препарата по Гимзе или использование других способов окрашивания; - анализ под микроскопом с целью выявления численных нарушений (гомогенных или в мозаике) и структурных аберраций; - фотографирование и вырезание хромосом; - идентификацию хромосом и составление кариограммы (идиограммы). Этапы кариотипирования Дифференциальная окраска хромосом 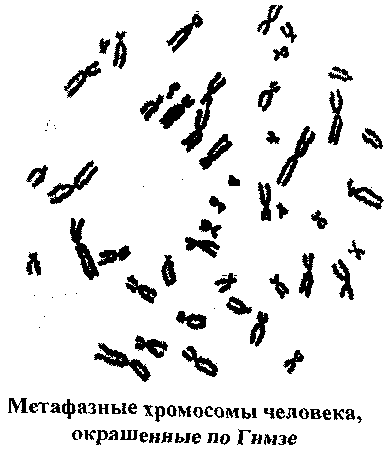 В настоящее время наряду с рутинными методами изучения кариотипа используются методы дифференциальной окраски, позволяющие выявить в хроматидах чередование окрашенных и неокрашенных полос. Они называются бэндами и имеют специфическое и точное распределение, обусловленное особенностями внутренней организации хромосомы Методы дифференциальной окраски были разработаны в начале 70-х годов ХХ-го века и стали важной вехой в развитии цитогенетики человека. Они имеют широкое практическое применение, т.к.: - чередование полос не носит случайный характер, а отражает внутреннюю структуру хромосом, например распределение эухроматиновых и гетерохроматиновых участков, богатых AT или GC последовательностями ДНК, участков хроматина с разной концентрацией гистонов и негистонов; - распределение бэндов идентично для всех клеток одного организма и всех организмов данного вида, что используется для точной идентификации вида; - метод позволяет точно идентифицировать гомологичные хромосомы, которые являются одинаковыми с генетической точки зрения и имеют сходное распределение бэндов; - метод обеспечивает точную идентификацию каждой хромосомы, т.к. разные хромосомы имеют разное распределение бэндов; - дифференциальная окраска позволяет выявить многие структурные нарушения хромосом (делеции, инверсии), которые с трудом обнаруживаются методами простой окраски. В зависимости от способа предобработки хромосом и техники окрашивания различают несколько методов дифференциальной окраски (G,Q,R,T,C). Используя их, можно получить чередование окрашенных и неокрашенных полос - бэндов, стабильных и специфичных для каждой хромосомы. 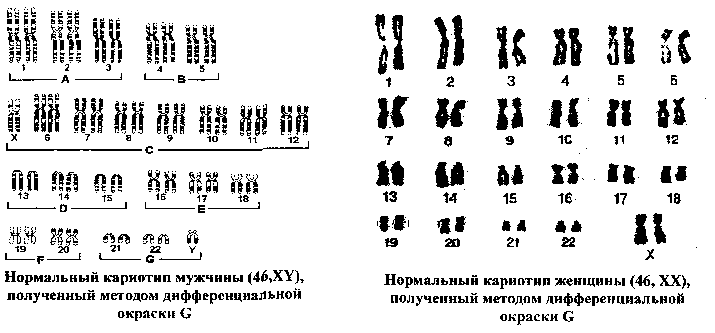  Характеристика различных методов дифференциальной окраски хромосом
Молекулярно-цитогенетические методы На основе методов молекулярного анализа нуклеиновых кислот была разработана серия новых техник изучения интерфазных хромосом, характеризующихся высокой специфичностью и точностью - методы гибридизации insitu (FISH, CGH, SKY). Метод FISH- флуоресцентная гибридизация insitu- включает следующие этапы: - создание зондов - однонитевых фрагментов ДНК, к которым присоединяют биотин или дигоксигенин; зонды могут быть комплементарны хромосоме или ее конкретному участку; - щелочная обработка препаратов insituс целью денатурации хромосомной ДНК за счет разрыва водородных связей между двумя цепями ДНК; - гибридизация хромосомной ДНК с зондом путем комплементарного связывания зонда со специфической последовательностью хромосомы; - обработка препаратов веществами, которые избирательно связываются с биотином (и дигоксегенином; для биотина таким веществом является стрептовидин, для дигоксегенина - антидигоксигениновое антитело; к этим веществам могут быть присоединены флуоресцентные красители (родамин или флуоресцеинт изотиоцианат); - визуализация хромосом с помощью люминесцентного микроскопа на фоне неокрашенных хромосом. Метод FISH используется для выявления: - численных хромосомных аномалий в интерфазном ядре; - структурных аномалий; - сложных межхромосомных перестроек; - учета симметричных аберраций у облученных лиц; - локализации гена. Этот достаточно простой, точный, быстрый и экономичный метод находит все более Широкое применение в диагностике врожденных аномалий, моногенных синдромов, в том числе, в пренатальной диагностике. CGH- метод сравнительной геномной гибридизации, используется в онкологической цитогенетике для выявления делетированных (выпавших) или амплифицированных участков хромосом при некоторых типах опухолей. Делетированные участки, как правило, содержат гены-фупрессоры опухолей, а амплифицированные участки - онкогены. Это позволяет использовать метод для картирования и клонирования генов, вовлеченных в канцерогенез. Метод CGHзаключается в следующем: ДНК, выделенную из опухоли и из нормальной ткани, метят разными флуорохромами. Меченую ДНК из опухоли и нормальной ткани гибридизуют с хромосомным препаратом, и по интенсивности свечения метки определяют участки, содержащие делеции и амплификации. Для обработки данных используют специальные программы компьютерного анализа. SKY - спектроскопический анализ хромосом. Метод основан на использовании набора зондов и флуоресцентных красителей, имеющих сродство к определенным участкам хромосом. Каждая пара хромосом, таким образом, имеет уникальные спектральные характеристики. Используя интерферометр, аналог аппарата для измерения спектра астрономических объектов, можно определить незначительные вариации в спектральном составе хромосом, неразличимые человеческим глазом. Данные подвергаются компьютерной обработке по специальной программе, в результате чего каждая пара хромосом приобретает определенный цвет. Преимущество метода состоит в более точной идентификации гомологичных хромосом, которые имеют один и тот же цвет, и возможности выявления некоторых транслокаций, которые другими цитогенетическими методами не определяются. Метод используется в онкоцитогенетике для выявления в кариотипе опухолевых клеток даже очень незначительных по величине хромосомных аберраций. НОМЕНКЛАТУРА ХРОМОСОМ ЧЕЛОВЕКА Номенклатура хромосом человека основана на международной системе стандартизации, которая позволяет обозначить нормальный или патологический кариотипы с использованием специальных символов и знаков, отражающих число хромосом и тип нарушения - недостаток, избыток или перераспределение хромосомного материала. Символы, используемые для описания кариотипа (международные стандарты)
В случае нормального кариотипа человеказаписывается общее число хромосом, за тем, после запятой, следует обозначение половых хромосом: нормальный кариотип женщины - 46ДХ нормальный кариотип мужчины - 46,XY. Аномальные кариотипымогут быть связаны с численными и структурными нарушениями, например: - аутосомные аномалии - в этом случае записывается общее число хромосом, запятая, половые хромосомы, запятая, + (лишняя хромосома) или - (отсутствие хромосомы), номер вовлеченной в нарушение хромосомы (например: 47, XX, +21 (трисомия 21); 47,XY,+13 (трисомия 13); 45,ХХ,-8 (моносомия 8); и.т.д.; - гоносомные аномалии - записывается хромосомное число, и после запятой указываются соответствующие половые хромосомы (например: 45,Х (моносомия X); 47,XXY (дисомия X); 47,ХХХ (трисомия X)). Используя метод дифференциальной окраски, в каждой хромосоме можно выявить реперы - важные элементы для идентификации хромосомы (рис. 6): - чередование бэндов, - центромера, - теломеры. Хромосомные реперы ограничивают районы хромосом, каждый из которых содержит несколько бэндов, а бэнды, в свою очередь, состоят из суббэндов. В метафазных хромосомах различают около 400-500 бэндов, в то время как в хромосомах ранней профазы можно выделить до 1800-2000 бэндов. Таким образом, метод изучения профазных хромосом отличается более высокой разрешающей способностью. 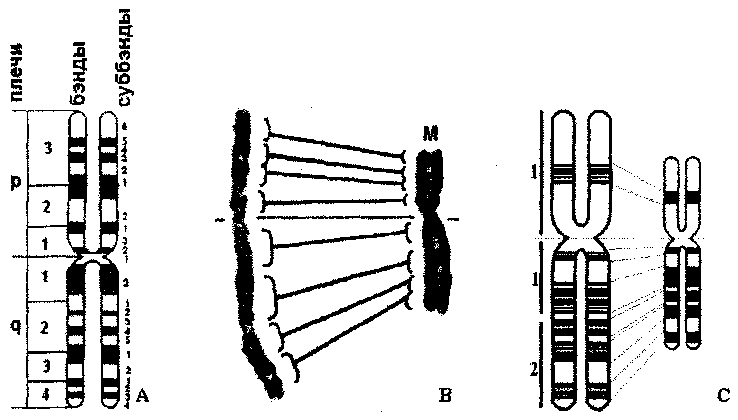 Номенклатура бэндов: участки и бэнды нумеруются от центромеры к теломеру для каждого плеча отдельно; например: 7ql2 - хромосома (7), дистальное плечо (q), участок (1), бэнд (2). При обозначении структурных хромосомных нарушений указывают тип перестройки, точки разрывов, бэнд и участок, вовлеченные в нарушение. Нарушение: - 46,XX,del(1)(q2 lq31) - деления в хромосоме 1, от района 2, бэнда 1 до района 3, бэнда 1. - 46,XY,r(2)(p21q31) - кольцевая хромосома 2; точки разрыва находятся в проксимальном плече: район 2 бэнд 1 и в дистальном плече: район 3 бэнд 1. - 46,XX,inv(2)(p21q31) - перицентрическая инверсия фрагмента от района 2 бэнда 1 проксимального плеча до района 3 бэнда 1 дистального плеча хромосомы 2. - 46,X,i(Xq) - изохромосома по дистальному плечу хромосомы X. ВАРИАЦИИ КАРИОТИПА В ПРЕДЕЛАХ НОРМАЛЬНОГО ФЕНОТИПА Существует ряд отклонений от нормального кариотипа, которые определяют незначительные вариации в пределах нормального фенотипа. К ним относятся: а) численные изменения: - у женщин - в возрасте после 60-ти лет до 7% соматических клеток могут терять хромосому X, в результате кариотип становится 45,Х; - у мужчин- в возрасте более 70-ти лет до 2% соматических клеток утрачивают хромосому Y, кариотип становится 45,Х. в) структурные изменения: - длина и форма гомологичных хромосомможет незначительно варьировать; изменения затрагивают чаще короткие плечи хромосом групп D, G (особенно гетерохроматиновые участки - например, хромосома Y может быть метацентрической); - спутники (сателлиты)обычно располагаются в акроцентрических хромосомах, за исключением хромосомы Y, но иногда могут наблюдаться и в других хромосомах - например, хромосомах 17, 18; при это они значительно варьируют по форме и размерам. - вторичные перетяжкипредставлены обычно в хромосомах 1, 9, 16 и Y, иногда перетяжка несколько увеличена. Хромосомный полиморфизм Полиморфизм бэндов (Q, G и С) отражает вариации по размерам и аспекту некоторых участков хромосом; чаще всего они затрагивают область центромеры, короткие плечи и спутники хромосом групп D и G, область вторичной перетяжки в длинном плече Y.  Значение полиморфизма: полиморфизм наследуется по доминантному типу, не меняя фенотипического проявления, т.к. обычно ограничен только гетерохроматиновыми участками, неактивными с генетической точки зрения (изменения касаются количества повторяющейся ДНК). Полиморфизм хромосом используют: - в качестве маркера передачи некоторых признаков от родителей детям (например, установление отцовства); - для установления родительского происхождения лишней хромосомы в случае анеуплоидий (например, происхождение дополнительных хромосом при трисомиях); - для идентификации хромосом, содержащих маркерный ген какой-то моногенной болезни; - для установления групп сцепления генов; - для анализа частоты полиморфизма хромосом при некоторых формах лейкемии, а также при наличии врожденных аномалий у детей. |