Вопрос 6
Стабильность градиента ионов достигается посредством активного транспорта: мембранные белки переносят ионы через мембрану против электрического и (или) концентрационного градиентов, потребляя для этого метаболическую энергию. Наиболее важный процесс активного транспорта - это работа Na/K-насоса, существующего практически во всех клетках; насос выкачивает ионы натрия из клетки, одновременно накачивая ионы калия внутрь клетки. Таким образом обеспечивается низкая внутриклеточная концентрация ионов натрия и высокая – калия. Градиент концентрации ионов натрия на мембране имеет специфические функции, связанные с передачей информации в виде электрических импульсов, а также с поддержанием других активных транспортных механизмов и регулирования объема клетки (см. ниже). Поэтому неудивительно, что более 1/3 энергии, потребляемой клеткой, расходуется на Na/К-насос, а в некоторых наиболее активных клетках на его работу расходуется до 70% энергии
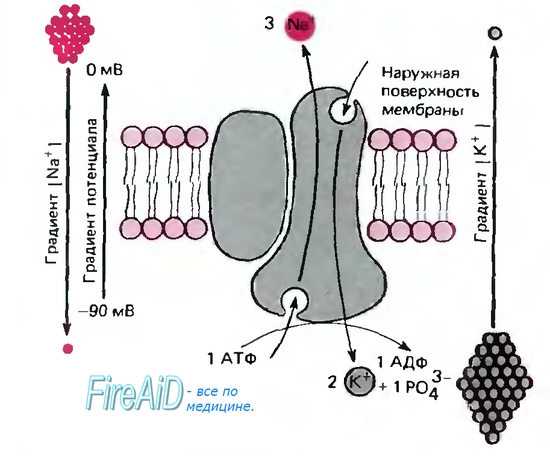
Рис 1.6. Схема Na/K-насоса АТФазы (погруженной в липидный бислой плазматической мембраны), которая за один цикл выносит из клетки три иона Na+ против градиентов потенциала и концентрации и приносит в клетку два иона К+. В ходе этого процесса одна молекула АТФ расщепляется на АДФ и фосфат. На схеме АТФаза показана как димер, состоящий из большой (функциональной) и малой субъединиц; в мембране она существует как тетрамер, образованный двумя большими и двумя малыми субъединицами
Na/K-транспортный белок представляет собой АТФазу. На внутренней поверхности мембраны она расщепляет АТФ на АДФ и фосфат (рис. 1.6). На транспортировку трех ионов натрия из клетки и одновременно двух ионов калия в клетку используется энергия одной молекулы АТФ, т. е. суммарно за один цикл из клетки удаляется один положительный заряд. Таким образом, Na/К-насос является электрогенным (создает электрический ток через мембрану), что приводит к увеличению электроотрицательности мембранного потенциала приблизительно на 10 мВ. Транспортный белок выполняет эту операцию с высокой скоростью: от 150 до 600 ионов натрия в секунду. Аминокислотная последовательность транспортного белка известна, однако еще не ясен механизм этого сложного обменного транспорта. Данный процесс описывают с использованием энергетических профилей переноса белками ионов натрия или калия (рис. 1.5,-6). По характеру изменения этих профилей, связанных с постоянными изменениями конформации транспортного белка (процесс, требующий затраты энергии), можно судить о стехиометрии обмена: два иона калия обмениваются на три иона натрия.
Na/К-насос, как и изолированная Na+/К-зависимая мембранная АТФаза, специфически ингибируется сердечным гликозидом уабаином (строфантином). Поскольку работа Na/K-насоса представляет собой многоступенчатую химическую реакцию, она, подобно всем химическим реакциям, в значительной степени зависит от температуры, что продемонстрировано на рис. 1.7. Здесь поток ионов натрия из мышечных клеток показан относительно времени; практически это эквивалентно потоку ионов натрия, опосредованному работой Na/K-насоса, потому что пассивный поток ионов натрия против градиентов концентрации и потенциала крайне мал. Если препарат охладить примерно на 18 С, то поток ионов натрия из клетки быстро уменьшится в 15 раз, а сразу после нагревания восстановится до исходного уровня. Такое уменьшение потока ионов натрия из клетки в несколько раз больше, чем то, которое бы соответствовало температурной зависимости процесса диффузии или простой химической реакции. Сходный эффект наблюдается, когда запас метаболической энергии истощается в результате отравления динитрофенолом (ДНФ) (рис. 1.7, Б). Следовательно, поток ионов натрия из клетки обеспечивается энергозависимой реакцией - активным насосом. Другой характеристикой насоса наряду со значительной температурной и энергетической зависимостью является наличие уровня насыщения (как и у всех других химических реакций); это означает, что скорость работы насоса не может возрастать бесконечно при повышении концентрации транспортируемых ионов (рис. 1.8). В отличие от этого поток пассивно диффундирующего вещества растет пропорционально разности концентраций в соответствии с законом диффузии

Рис. 1.7. А, Б Активный транспорт Na+. Ось ординат поток радиоактивного 24Na+ из клетки (имг.-мин-1). Ось абсцисс: время с начала эксперимента. А. Клетка охлаждена с 18,3c до 0,5 С; поток Na+ из клетки в этот период заторможен.
Б. Подавление потока Na+ из клетки динитрофенолом (ДНФ) в концентрации 0-2 ммоль/л (по [13] с изменениями)
Помимо Na/K-насоса плазматическая мембрана содержит по крайней мере еще один насос—кальциевый; это насос откачивает ионы кальция (Са2+) из клетки и участвует в поддержании их внутриклеточной концентрации на крайне низком уровне. Кальциевый насос присутствует с очень высокой плотностью в саркоплазматическом ретикулуме мышечных клеток, которые накапливают ионы кальция в результате расщепления молекул АТФ
Вопрос 7
Мембранный потенциал - разность электрических потенциалов между наружной и внутренней поверхностями биологической мембраны, обусловленная неодинаковой концентрацией ионов, гл. обр. натрия, калия и хлора.
Биоэлектрические потенциалы — электрические потенциалы, возникающие в живых клетках и тканях; показатель биоэлектрической активности, определяемой разностью электрических потенциалов между двумя точками живой ткани.
Основными видами Б. п. являются мембранный потенциал (или потенциал покоя), потенциал действия, постсинаптические потенциалы. Другие виды Б. п. различных органов и тканей (рецепторные, секреторные, потенциалы сердца, головного мозга и др.) являются аналогами или производными вышеперечисленных Б. п. Мембранный потенциал (потенциал покоя) регистрируется между наружной и внутренней сторонами мембраны живой клетки. Его наличие обусловлено неравномерным распределением ионов (в первую очередь ионов натрия и калия) между внутренним содержанием клетки (ее цитоплазмой) и окружающей клетку средой (см. Мембраны биологические). Внутренняя сторона мембраны заряжена отрицательно по отношению к наружной (рис. 1). Величина мембранного потенциала различна у разных клеток: для нервной клетки она составляет 60—80 мВ, для поперечнополосатых мышечных волокон — 80—90 мВ, для волокон сердечной мышцы — 90—95 мВ. При неизменном функциональном состоянии клетки величина потенциала покоя не изменяется; поддержание постоянной его величины обеспечивается нормальным протеканием клеточного метаболизма. Под влиянием различных факторов (раздражителей) физической или химической природы величина мембранного потенциала может изменяться. Увеличение разности потенциалов между клеткой и окружающей средой называется гиперполяризацией, уменьшение — деполяризацией.
При уменьшении потенциала покоя до определенной критической величины (порог возбуждения) возникает кратковременное колебание, получившее название потенциала действия. Если потенциал покоя присущ всем живым клеткам без исключения, то потенциал действия характерен в основном для специализированных возбудимых образований, является показателем развития процесса возбуждения. Вслед за потенциалом действия (пиковый потенциал, или спайк) возникает следовая деполяризация мембраны (отрицательный следовой потенциал) и последующая ее гиперполяризация (положительный следовой потенциал). Амплитуда потенциала действия у большинства нервных клеток млекопитающих составляет 100—110 мВ, у скелетных и сердечных мышечных волокон — 110—120 мВ. Длительность потенциалов действия у нервных клеток 1—2 мс, у скелетных мышечных волокон 3—5 мс, у сердечных мышечных волокон — 50—600 мс. Следовые потенциалы по своей длительности намного превышают потенциал действия. Потенциал действия обеспечивает распространение возбуждения от рецепторов к нервным клеткам, от нервных клеток к мышцам, железам, тканям. В мышечном волокне потенциал действия способствует осуществлению цепи физико-химических и ферментативных реакций, лежащих в основе механизма сокращения мышц.
Постсинаптические потенциалы (возбуждающий и тормозящий) возникают на небольших участках клеточной мембраны (постсинаптической мембране), входящих в состав синапса. Величина постсинаптических потенциалов составляет несколько милливольт, длительность — 10—15 мс. Возбуждающий постсинаптический потенциал (ВПСП) связан с деполяризацией клеточной мембраны. При достижении критической точки деполяризации возникает распространяющийся потенциал действия (рис. 2). Тормозящий постсинаптический потенциал (ТПСП), связанный с гиперполяризацией клеточной мембраны, препятствует возникновению потенциала действия.
Механизм возникновения Б. п. связан с наличием определенных физико-химических градиентов между отдельными тканями организма, между жидкостью, окружающей клетку, и ее цитоплазмой, между отдельными клеточными элементами. Во всех случаях местом возникновения градиентов являются мембраны, различающиеся не только по своей структуре, но и по ионообменным свойствам. Возникновение Б. п. в живых клетках обусловлено неравномерной концентрацией ионов натрия, калия, кальция и хлора на внутренней и наружной поверхности клеточной мембраны и ее различной проницаемостью для них. Величина мембранного потенциала покоя определяется соотношением концентраций, проникающих через мембрану ионов. Высокие концентрационные градиенты ионов калия и натрия поддерживаются благодаря существованию в клеточной мембране так называемого калиево-натриевого насоса, который обеспечивает выделение из цитоплазмы проникающих в нее ионов натрия и введение в цитоплазму ионов К+. Подобный насос работает против их концентрационных градиентов и требует для этого энергии. Источником энергии является аденозинтрифосфорная кислота (АТФ). Энергия, выделяемая при расщеплении локализованной в мембране АТФ-азой одной молекулы АТФ, обеспечивает выделение из клетки трех ионов натрия взамен на два иона калия, поступающих в клетку.
Механизм возникновения потенциала действия обусловлен последовательно изменяющейся во времени проницаемостью мембраны для ионов. Восходящая фаза потенциала действия связана с повышением проницаемости для ионов натрия благодаря все увеличивающемуся количеству открываемых натриевых каналов. Последующая смена активации натриевых каналов на их инактивацию приводит к снижению проницаемости для ионов натрия и возрастанию проницаемости для ионов калия, что приводит к реполяризации мембраны и появлению ее потенциала покоя. В гладких мышцах в отличие от нервных клеток и скелетных мышц в генезе восходящей фазы потенциала действия ведущая роль отводится повышению проницаемости для ионов кальция. В мышце сердца сохранение потенциала действия на определенном уровне (плато потенциала действия) также обусловлено повышением проницаемости мембраны для ионов кальция.
На мембранах секреторных клеток формируются секреторные потенциалы. Их величина прямо связана с характером секреторной деятельности, что дает возможность оценивать функциональное состояние секреторных клеток. В тканях или органах может происходить суммация биоэлектрической активности отдельных клеток, работающих синхронно или асинхронно. Суммарная биоэлектрическая активность также отражает функциональное состояние того или иного органа или ткани.
Исследование Б. п. нашло широкое применение в медико-биологических лабораториях, в клинической практике при диагностике различных заболеваний ц.н.с., сердечно-сосудистой и мышечной систем. При отведении суммарных Б. п. от нервных стволов, мышц, головного мозга, сердца и других органов применяют поверхностные макроэлектроды (см. Электрокардиография, Электромиография, Электроэнцефалография). В некоторых случаях используют внутриполостные электроды или вводимые непосредственно в ткань (например, игольчатые). Для регистрации и измерения Б. п. отдельных клеток чаще всего пользуются внутриклеточными и точечно-внеклеточными микроэлектродами. Электроды соединяют с усилителями переменного или постоянного тока, входящими в комплект серийно выпускаемых медицинских приборов. Усилитель может быть связан с устройством автоматизированной обработки биоэлектрических сигналов.
Вопрос 8
Уравнение Нернста — уравнение, связывающее окислительно-восстановительный потенциал системы с активностями веществ, входящих в электрохимическое уравнение, и стандартными потенциалами окислительно-восстановительных пар.

,
где E - электродный потенциал, E0 - стандартный электродный потенциал, измеряется в вольтах;
R — универсальная газовая постоянная, равная 8.31 Дж/(моль·K);
T— абсолютная температура;
F — число Фарадея, равное 96485,35 Кл/моль;
n— число моль электронов, участвующих в процессе;
И 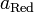 — активности соответственно окисленной и восстановленной форм вещества, участвующего в полуреакции. — активности соответственно окисленной и восстановленной форм вещества, участвующего в полуреакции.
Если в формулу Нернста подставить числовые значения констант R и F и перейти от натуральных логарифмов к десятичным, то при T = 298K получим
МОДЕЛЬ ЭЛЕКТРОГЕНЕЗА ЖИВЫХ ТКАНЕЙ ГОЛЬДМАНА [ Goldman's model of bioelectrogenesis, Goldman-Hodgkin-Katz equation ]
Модель электрогенеза живых тканей Голдмана, или модель Голдмана-Ходжкина-Катца, или уравнение Голдмана-Ходжкина-Катца - одна из концептуальных физико-химических моделей, описывающих механизмы существования электромагнитных явлений в живой ткани.
Непосредственным источником электромагнитной энергии в любой клетке организма является динамический концентрационный элемент, образованный цитоплазматической мембраной и растворами ионов, которые неравновесно распределены между цитоплазмой и межклеточной жидкостью. Цитоплазматическая мембрана обладает неодинаковой проницаемостью для катионов и анионов и может изменяться в зависимости от разных причин. Процессы электрогенеза, протекающие на мембране являются стохастическими процессами.
Простейшей материальной физико-химической моделью, демонстрирующей механизмы электрогенеза в живых тканях является модель Нернста, а соответствующей простейшей математической моделью - уравнение Нернста. Эта простейшая модель рассматривает раствор только одной соли. Живые ткани содержат значительное число разных электролитов. Математической моделью, аналогичной модели Нернста, но описывающей механизмы электрогенеза с учетом наличия многих электролитов, является модель Гольдмана-Ходжкина-Катца: ЭДС=(RT:F)·ln((pК1CК1e+pК2CК2e+…+pA1CA1i…):(pК1CК1i+pК2CК2i+…+pA1CA1e…)). Здесь, как и в модели Нернста: R - универсальная газовая постоянная, Т - абсолютная температура растворов, F - число Фарадея, CK1 - концентрация катиона 1 (например, калия), CK2 - концентрация катиона 2 (например, натрия), CA1 - концентрация аниона 1 (например, хлора), pK - проницаемость мембраны для катиона, pA - проницаемость мембраны для аниона, индексы при символах CK1,…, CA1: i - на внутренней поверхности мембраны, e - на наружной поверхности мембраны.
NB: Модель Голдмана, как и модель Нернста, основана на допущении, что мембрана обладает нерегулируемой проницаемостью для ионов, обусловленной размером пор мембраны. Авторы модели не делали допущений относительно сущности процессов на мембране. На основании того, что эта модель аналитическая (но не вероятностная), можно полагать, что авторы считали само собой разумеющимся, что процессы на мембране по своей сущности жестко детерминированы. Очевидно, что в действительности, процессы на мембране являются вероятностными. Полное их описание должно включать по крайней мере две функции: зависимость математического ожидания от концентраций и проводимостей как случайных переменных и зависимость дисперсии от тех же переменных. Помимо того, процессы на мембране не являются пассивными. Это активные управляемые процессы. В частности, движение ионов через плазмалемму клетки может осуществляться не только путем простой диффузии, но и путем активного транспорта (натриевый насос). Можно предположить, что те ученые, которые признают справедливость положений вероятностной методологии (Трифонов Е.В., 1978, …, 2011, …).Трифонов Е.В., 1978, …, 2002, ….), смогут первыми подтвердить эти новые идеи относительно вероятностной природы электрогенеза.
Более жестким возражением против принятия детерминистских моделей электрогенеза, как истины последней инстанции, является то, что эти модели основаны на детерминистских предпосылках современной физики и химии. А эти предпосылки ложны. Мир есть иерархия не детерминистских, но вероятностных сущностей и явлений! Следовательно и предпосылки должны быть соответствующими реальности.
Еще более жестким возражением против принятия детерминистских физико-химических моделей электрогенеза, как истины последней инстанции, является то, что физико-химические сущности и явления не эквивалентны живым сущностям и явлениям. Некоторые физиологи еще в позапрошлом веке (19 в) понимали это. Вспомним аргументированную конструктивную критику И.М. Сеченова (1862 г.) «теории» биоэлектрогенеза отца электрофизиологии Э. Дюбуа-Реймона. Еще тогда И.М.Сеченов говорил: «Электрические явления мышц и нервов суть продукты их жизни» (курсив мой, см. статью Электрофизиология). Сколько же еще столетий понадобится ученым, чтобы осознать разницу между жизнью и физикой и химией?!
И, наконец, еще более жестким возражением против принятия детерминистских моделей электрогенеза, как истины последней инстанции, является то, что эти модели основаны на исключительно материалистических предпосылках современной физики и химии. Одной из таких предпосылок является запрет на все, что лежит за пределами материализма, то есть запрет метафизики и метанауки вообще. Простите, но это просто глупо, и непрактично отрицать то, чем человек пользуется со времени своего возникновения! Впрочем, это уже выходит за рамки обозначенной темы, но относится не более и не менее как ко всей современной науке.
вопрос 9
Потенциа́л де́йствия — волна возбуждения, перемещающаяся по мембране живой клетки в процессе передачи нервного сигнала. По сути своей представляет электрический разряд — быстрое кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона, мышечного волокна или железистой клетки), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного или мышечного импульса, играющего сигнальную (регуляторную) роль.
Потенциалы действия могут различаться по своим параметрам в зависимости от типа клетки и даже на различных участках мембраны одной и той же клетки. Наиболее характерный пример различий: потенциал действия сердечной мышцы и потенциал действия большинства нейронов. Тем не менее, в основе любого потенциала действия лежат следующие явления:
Мембрана живой клетки поляризована — её внутренняя поверхность заряжена отрицательно по отношению к внешней благодаря тому, что в растворе возле её внешней поверхности находится бо́льшее количество положительно заряженных частиц (катионов), а возле внутренней поверхности — бо́льшее количество отрицательно заряженных частиц (анионов).
Мембрана обладает избирательной проницаемостью — её проницаемость для различных частиц (атомов или молекул) зависит от их размеров, электрического заряда и химических свойств.
Мембрана возбудимой клетки способна быстро менять свою проницаемостъ для определённого вида катионов, вызывая переход положительного заряда с внешней стороны на внутреннюю.
Первые два свойства характерны для всех живых клеток. Третье же является особенностью клеток возбудимых тканей и причиной, по которой их мембраны способны генерировать и проводить потенциалы действия.
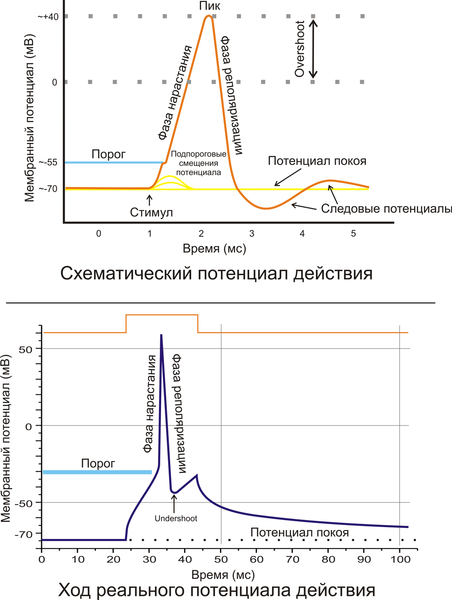
Фазы потенциала действия
Предспайк — процесс медленной деполяризации мембраны до критического уровня деполяризации (местное возбуждение, локальный ответ).
Пиковый потенциал, или спайк, состоящий из восходящей части (деполяризация мембраны) и нисходящей части (реполяризация мембраны).
Отрицательный следовой потенциал — от критического уровня деполяризации до исходного уровня поляризации мембраны (следовая деполяризация).
Положительный следовой потенциал — увеличение мембранного потенциала и постепенное возвращение его к исходной величине (следовая гиперполяризация).
Общие положения
Поляризация мембраны живой клетки обусловлена отличием ионного состава с её внутренней и наружной стороны. Когда клетка находится в спокойном (невозбуждённом) состоянии, ионы по разные стороны мембраны создают относительно стабильную разность потенциалов, называемую потенциалом покоя. Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя, он будет иметь отрицательное значение (порядка −70 — −90 мВ). Это объясняется тем, что суммарный заряд на внутренней стороне мембраны существенно меньше, чем на внешней, хотя с обеих сторон содержатся и катионы, и анионы. Снаружи — на порядок больше ионов натрия, кальция и хлора, внутри — ионов калия и отрицательно заряженных белковых молекул, аминокислот, органических кислот, фосфатов, сульфатов. Надо понимать, что речь идёт именно о заряде поверхности мембраны — в целом среда и внутри, и снаружи клетки заряжена нейтрально.
Потенциал мембраны может изменяться под действием различных стимулов. Искусственным стимулом может служить электрический ток, подаваемый на внешнюю или внутреннюю сторону мембраны через электрод. В естественных условиях стимулом часто служит химический сигнал от соседних клеток, поступающий через синапс или путём диффузной передачи через межклеточную среду. Смещение мембранного потенциала может происходить в отрицательную (гиперполяризация) или положительную (деполяризация) сторону.
В нервной ткани потенциал действия, как правило, возникает при деполяризации — если деполяризация мембраны нейрона достигает некоторого порогового уровня или превышает его, клетка возбуждается, и от её тела к аксонам и дендритам распространяется волна электрического сигнала. (В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы, которые сильно отличаются от потенциала действия по своей природе — например, они не подчиняются принципу «всё или ничего». Эти потенциалы преобразуются в потенциал действия на особом участке мембраны — аксонном холмике, так что потенциал действия не распространяется на дендриты).
Рис. 3. Простейшая схема, демонстрирующая мембрану с двумя натриевыми каналами в открытом и закрытом состоянии, соответственно
Это обусловлено тем, что на мембране клетки находятся ионные каналы — белковые молекулы, образующие в мембране поры, через которые ионы могут проходить с внутренней стороны мембраны на наружную и наоборот. Большинство каналов ионоспецифичны — натриевый канал пропускает практически только ионы натрия и не пропускает другие (это явление называют селективностью). Мембрана клеток возбудимых тканей (нервной и мышечной) содержит большое количество потенциал-зависимых ионных каналов, способных быстро реагировать на смещение мембранного потенциала. Деполяризация мембраны в первую очередь вызывает открытие потенциал-зависимых натриевых каналов. Когда одновременно открывается достаточно много натриевых каналов, положительно заряженные ионы натрия устремляются через них на внутреннюю сторону мембраны. Движущая сила в данном случае обеспечивается градиентом концентрации (с внешней стороны мембраны находится намного больше положительно заряженных ионов натрия, чем внутри клетки) и отрицательным зарядом внутренней стороны мембраны (см. Рис. 2). Поток ионов натрия вызывает ещё бо́льшее и очень быстрое изменение мембранного потенциала, которое и называют потенциалом действия (в специальной литературе обозначается ПД).
Согласно закону «всё-или-ничего» мембрана клетки возбудимой ткани либо не отвечает на стимул совсем, либо отвечает с максимально возможной для неё на данный момент силой. То есть, если стимул слишком слаб и порог не достигнут, потенциал действия не возникает совсем; в то же время, пороговый стимул вызовет потенциал действия такой же амплитуды, как и стимул, превышающий пороговый. Это отнюдь не означает, что амплитуда потенциала действия всегда одинакова — один и тот же участок мембраны, находясь в разных состояниях, может генерировать потенциалы действия разной амплитуды.
После возбуждения нейрон на некоторое время оказывается в состоянии абсолютной рефрактерности, когда никакие сигналы не могут его возбудить снова, затем входит в фазу относительной рефрактерности, когда его могут возбудить исключительно сильные сигналы (при этом амплитуда ПД будет ниже, чем обычно). Рефрактерный период возникает из-за инактивации быстрого натриевого тока, то есть инактивации натриевых каналов (см. ниже).
Распространение потенциала действия
Распространение потенциала действия по немиелинизированным волокнам
По немиелинизированному волокну ПД распространяется непрерывно. Проведение нервного импульса начинается с распространением электрического поля. Возникший ПД за счет электрического поля способен деполяризовать мембрану соседнего участка до критического уровня, в результате чего на соседнем участке генерируются новые ПД. Сами ПД не перемещаются, они исчезают там же, где возникают. Главную роль в возникновении нового ПД играет предыдущий.
Если внутриклеточным электродом раздражать аксон посередине, то ПД будет распространяться в обоих направлениях. Обычно же ПД распространяется по аксону в одном направлении (от тела нейрона к нервным окончаниям), хотя деполяризация мембраны происходит по обе стороны от участка, где в данный момент возник ПД. Одностороннее проведение ПД обеспечивается свойствами натриевых каналов — после открывания они на некоторое время инактивируются и не могут открыться ни при каких значениях мембранного потенциала (свойство рефрактерности). Поэтому на ближнем к телу клетки участке, где до этого уже «прошел» ПД, он не возникает.
При прочих равных условиях распространение ПД по аксону происходит тем быстрее, чем больше диаметр волокна. По гигантским аксонам кальмара ПД может распространяться почти с такой же скоростью, как и по миелинизированным волокнам позвоночных (около 100 м/c).
Распространение потенциала действия по миелинизированным волокнам
По миелинизированному волокну ПД распространяется скачкообразно (сальтаторное проведение). Для миелинизированных волокон характерна концентрация потенциалзависимых ионных каналов только в областях перехватов Ранвье; здесь их плотность в 100 раз больше, чем в мембранах безмиелиновых волокон. В области миелиновых муфт потенциалзависимых каналов почти нет. ПД, возникший в одном перехвате Ранвье, за счет электрического поля деполяризует мембрану соседних перехватов до критического уровня, что приводит к возникновению в них новых ПД, то есть возбуждение переходит скачкообразно, от одного перехвата к другому. В случае повреждения одного перехвата Ранвье ПД возбуждает 2-ой, 3-ий, 4-ый и даже 5-ый, поскольку электроизоляция, создаваемая миелиновыми муфтами, уменьшает рассеивание электрического поля. Это увеличивает скорость распространения ПД по миелинизированным волокнам по сравнению с немиелинизированными. Кроме того, миелинизированные волокна толще, а электрическое сопротивление более толстых волокон меньше, что тоже увеличивает скорость проведения импульса по миелинизированным волокнам. Другим преимуществом сальтаторного проведения является его экономичность в энергетическом плане, так как возбуждаются только перехваты Ранвье, площадь которых меньше 1 % мембраны, и, следовательно, необходимо значительно меньше энергии для восстановления трансмембранных градиентов Na+ и K+, расходующихся в результате возникновения ПД, что может иметь значение при высокой частоте разрядов, идущих по нервному волокну.
Чтобы представить, насколько эффективно может быть увеличена скорость проведения за счёт миелиновой оболочки, достаточно сравнить скорость распространения импульса по немиелинизированным и миелинизированным участкам нервной системы человека. При диаметре волокна около 2 µм и отсутствии миелиновой оболочки скорость проведения будет составлять 1 м/с, а при наличии даже слабой миелинизации при том же диаметре волокна — 15-20 м/с. В волокнах большего диаметра, обладающих толстой миелинововой оболочкой, скорость проведения может достигать 120 м/с.
Скорость распространения потенциала действия по мембране отдельно взятого нервного волокна отнюдь не является постоянной величиной — в зависимости от различных условий, эта скорость может очень значительно уменьшаться и, соответственно, увеличиваться, возвращаясь к некоему исходному уровню.
Активные свойства мембраны
Схема строения мембраны клетки.
Активные свойства мембраны, обеспечивающие возникновение потенциала действия, основываются главным образом на поведении потенциалзависимых натриевых (Na+) и калиевых (K+) каналов. Начальная фаза ПД формируется входящим натриевым током, позже открываются калиевые каналы и выходящий K+-ток возвращает потенциал мембраны к исходному уровню. Исходную концентрацию ионов затем восстанавливает натрий-калиевый насос.
По ходу ПД каналы переходят из состояния в состояние: у Na+ каналов основных состояний три — закрытое, открытое и инактивированное (в реальности дело сложнее, но этих трёх достаточно для описания), у K+ каналов два — закрытое и открытое.
Поведение каналов, участвующих в формировании ПД, описывается через проводимость и высчиляется через коэффициенты переноса (трансфера).
Коэффициенты переноса были выведены Ходжкиным и Хаксли.
Кривая ПД
Вопрос 10
Механические колебания – это повторяющееся движение, при котором тело
многократно проходит одно и то же положение в пространстве. Различают
периодические и непериодические колебания. Периодическими называют
колебания, при которых координата и другие характеристики тела описываются
периодическими функциями времени.
Примерами механических колебаний могут служить движение шара на пружине,
на нити, движение ножек звучащего камертона или молекул воздуха вблизи
него (рис. 1). В физике рассматривают и другие колебания – процессы, обладающие
той или иной степенью повторяемости во времени (например, электромагнитные
колебания.)
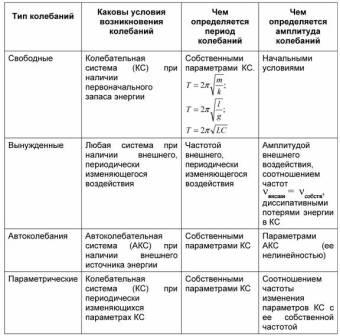
Колебания можно классифицировать по условиям возникновения (свободные,
вынужденные, автоколебания) и по характеру изменения во времени кинематических
характеристик (пилообразные, гармонические, затухающие)
Классификация колебаний :
Для описания кинематических характеристик используют аналитическую
зависимость характеристики, например координаты или скорости от
времени u и графическое представление этой функции (рис. 2, а сложной формы,
б прямоугольные, в пилообразные, г гармонические, д затухающие, е нарастающие). – – – – – –(t)
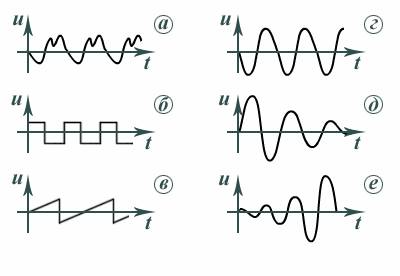
Рис. 2
Наиболее общими характеристиками колебаний являются следующие физические
величины: амплитуда колебаний А наибольшее отклонение колеблющегося
тела от положения равновесия (отклонение величины от ее среднего значения);
период колебаний Т время, через которое движение тела полностью
повторяется (повторяются все кинематические характеристики колебаний),
т.е. совершается одно полное колебание; частота колебаний v – величина,
показывающая число колебаний, совершаемых за 1 с. Вместо частоты v чаще
пользуются понятием циклической частоты w. Циклическая частота w – это
число колебаний, совершаемых за 2p секунд. Частота обратно пропорциональна
периоду: – –
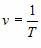
И
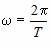
В СИ период Т выражается в секундах (c), частота v в герцах (Гц), циклическая
частота w – в обратных секундах (с–1). –
Единица амплитуды колебаний зависит от того, какая колеблющаяся физическая
величина рассматривается.
Для сравнения колебаний, происходящих с одной частотой, но различающихся по
тому, какую стадию полного колебания проходит тело, вводят понятие фазы
колебаний. Если два шарика на нитях одинаковой длины отвести от положения
равновесия вправо и отпустить, то они будут колебаться в фазе (синфазно,
синхронно), если их развести в разные стороны, то колебания будут происходить
в противофазе.
При описании колебаний с помощью функции изменения кинематической величины
во времени фазой j называют аргумент функции, описывающей колебательный
процесс.
Уравнение гармонических колебаний
Гармонические колебания – колебания, при которых физическая величина,
характеризующая эти колебания, изменяется во времени по синусоидальому
закону
x = A sin (wt + j0),
где x значение колеблющейся величины в момент времени t, A амплитуда колебаний,
w – циклическая (или круговая) частота, (wt + j0) – фаза гармонических колебаний,
j0
– начальная фаза. – –
Графиком гармонических колебаний является синусоида (рис. 3)

Рис. 3
Выбор начальной фазы позволяет при описании гармонических колебаний перейти
от функции синуса к функции косинуса.
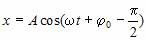
,
Гармонические колебания :

Если координата материальной точки меняется по гармоническому закону
x(t) = A sin (wt + j0),
то скорость, импульс и ускорение тоже меняются по гармоническому закону
(t) = Aw cos (wt + j0),a(t) = –Aw2 sin (wt + j0).
Сравнивая выражения для x и a получаем соотношение(t),(t)
a + w2x = 0,
которое принято считать определением гармонических колебаний в динамике.
В случае гармонических колебаний справедливо также соотношение
2 + w2x2 = const,
которое удобно использовать для получения периода гармонических колебаний
в различных колебательных системах. |
 Скачать 0.71 Mb.
Скачать 0.71 Mb.