моделирование. компьютерное моделирование в экологии
 Скачать 177.21 Kb. Скачать 177.21 Kb.
|

|
Информатика А.В.Могилев, Н.И.Пак, Е.К.Хённер § 4. КОМПЬЮТЕРНОЕ МОДЕЛИРОВАНИЕ В ЭКОЛОГИИ4.1. ЭКОЛОГИЯ И МОДЕЛИРОВАНИЕЭкология - одно из слов, появившихся сравнительно недавно у всех на устах и на страницах газет и журналов. Еще в 60-х годах нашего столетия почти никто, кроме узких специалистов, его не знал, да и большинство из тех, кто знал, использовал в таком смысле, который вряд ли способен заинтересовать широкую общественность. А между тем, термину более 120 лет. В 1869 г. немецкий естествоиспытатель Эрнст Геккель предложил составной термин «экология» («эко» - дом, жилище, местопребывание и «логос» - наука, знание) как название раздела биологии, ставшего самостоятельным. Классическая экология - наука о взаимодействии организмов и окружающей среды. Сегодня, говоря об экологии, чаще всего имеют в виду не классическую, а, так называемую, социальную экологию, оформившуюся как научное направление и направление общественно-политической деятельности на 100 лет позднее, и занимающуюся проблемами охраны окружающей среды, взаимодействием с ней человеческого сообщества. В данной главе мы ограничимся некоторыми классическими моделями «старой» экологии, что обусловлено следующими причинами. Во-первых, они достаточно просты и изучены, постановка их вполне очевидна и в познавательном плане интересна и полезна. Во-вторых, модели распространения загрязнений окружающей среды требуют использования весьма сложного математического аппарата, да и сами еще не вполне устоялись. Проблемы охраны окружающей среды чрезвычайно важны, но их обсуждение выходит за пределы нашего курса. Однако, для того, чтобы дать представление о задачах, стоящих перед современными исследователями в этой области, в следующем параграфе приведено описание одной из глобальных моделей, пытающихся выяснить пути взаимодействия экосистемы планеты с индустриальной и экономической системами современного общества. Остановимся на некоторых понятиях, которые будут встречаться в этой главе. Под особью понимается отдельный индивидуум, отдельный организм. Популяция -это совокупность особей одного вида, существующих в одно и то же время и занимающих определенную территорию. И, наконец, сообщество - это совокупность совместно сосуществующих популяций. В классической экологии рассматриваются взаимодействия нескольких типов: • взаимодействие организма и окружающей среды; • взаимодействие особей внутри популяции; • взаимодействие между особями разных видов (между популяциями). Математические модели в экологии используются практически с момента возникновения этой науки. И, хотя поведение организмов в живой природе гораздо труднее адекватно описать средствами математики, чем самые сложные физические процессы, модели помогают установить некоторые закономерности и общие тенденции развития отдельных популяций, а также сообществ. Кажется удивительным, что люди, занимающиеся живой природой, воссоздают ее в искусственной математической форме, но есть веские причины, которые стимулируют эти занятия. Вот некоторые цели создания математических моделей в классической экологии. 1. Модели помогают выделить суть или объединить и выразить с помощью нескольких параметров важные разрозненные свойства большого числа уникальных наблюдений, что облегчает экологу анализ рассматриваемого процесса или проблемы. 2. Модели выступают в качестве «общего языка», с помощью которого может быть описано каждое уникальное явление, и относительные свойства таких явлений становятся более понятными. 3. Модель может служить образцом «идеального объекта» или идеализированного поведения, при сравнении с которым можно оценивать и измерять реальные объекты и процессы. 4. Модели действительно могут пролить свет на реальный мир, несовершенными имитациями которого они являются. При построении моделей в математической экологии используется опыт математического моделирования механических и физических систем, однако с учетом специфических особенностей биологических систем: • сложности внутреннего строения каждой особи; • зависимости условий жизнедеятельности организмов от многих факторов внешней среды; • незамкнутости экологических систем; • огромного диапазона внешних характеристик, при которых сохраняется жизнеспособность систем. Привлечение компьютеров существенно раздвинуло границы моделирования экологических процессов. С одной стороны, появилась возможность всесторонней реализации сложных математических моделей, не допускающих аналитического исследования, с другой - возникли принципиально новые направления, и прежде всего - имитационное моделирование. 4.2. МОДЕЛИ ВНУТРИВИДОВОЙ КОНКУРЕНЦИИРассмотрим простейшую из указанных моделей для вида с дискретными периодами размножения, в которой численность популяции в момент времени t равна N, и изменяется во времени пропорционально величине основной чистой скорости воспроизводства R. Такими видами являются, например, большая часть растений, некоторые виды насекомых, у которых разные поколения четко разнесены во времени. Коэффициент R характеризует количество особей, которое воспроизводится в расчете на одну существующую, а также выживание уже существующих. Данная модель может быть выражена уравнением решение которого имеет вид где N0- начальная численность популяции. Эта модель, однако, описывает популяцию, в которой отсутствует конкуренция и в которой R является константой; если R>1, то численность популяции будет бесконечно увеличиваться. В реальности в какой-то момент начинают работать механизмы сдерживания роста популяции. В литературе приводится немало интересных примеров быстрого роста численности популяций, если бы для их размножения существовали идеальные условия. Особенно это относится к насекомым, растениям и микроорганизмам, которые могли бы покрыть земной шар толстым слоем, если им создать благоприятные условия для размножения. Но в действительности такого роста популяций, когда их численность увеличивается в геометрической прогрессии, на сколько-нибудь длительных промежутках времени не наблюдается. Следовательно, в первую очередь необходимо изменить уравнение (7.60) таким образом, чтобы чистая скорость воспроизводства зависела от внутривидовой конкуренции. Конкуренцию можно определить как использование некоего ресурса (пищи, воды, света, пространства) каким-либо организмом, который тем самым уменьшает доступность этого ресурса для других организмов. Если конкурирующие организмы принадлежат к одному виду, то взаимоотношения между ними называют внутривидовой конкуренцией, если же они относятся к разным видам, то их взаимоотношения называют межвидовой конкуренцией. 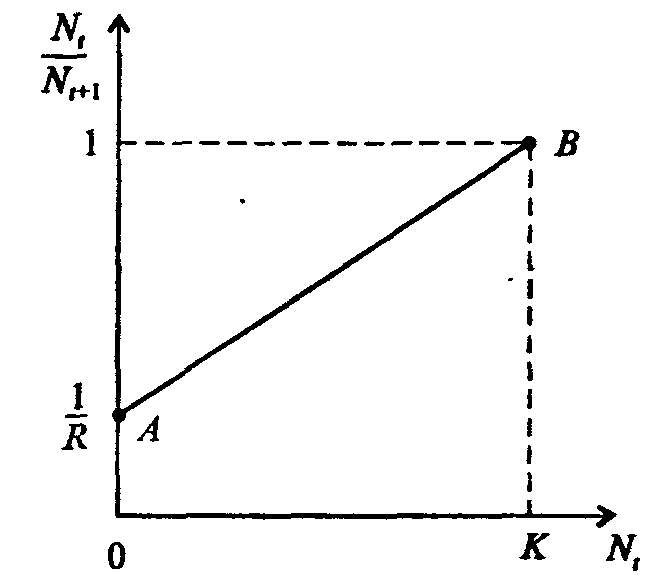 Рис. 7.37. К вопросу об ограничении скорости роста популяции На рис. 7.37 показана простейшая возможная зависимость скорости воспроизводства от численности популяции. Точка А отражает ситуацию, в которой численность популяции близка к нулю, конкуренция при этом практически отсутствует, и фактическую скорость воспроизводства вполне можно описывать параметром R в его первоначальном виде. Следовательно, при низкой плотности популяции уравнение (7.60) вполне справедливо. В преобразованном виде оно выглядит так: Точка В, напротив, отражает ситуацию, в которой численность популяции высока, и в значительной степени проявляется внутривидовая конкуренция. Фактическая скорость воспроизводства в результате конкуренции настолько снижена, что популяция в целом может не более чем восстанавливать в каждом поколении свою численность, потому что количество родившихся особей уравновешивается количеством погибших. Гипотезе, отраженной на рис. 7.37, соответствует уравнение где  . Это уравнение представляет собой модель роста популяции, ограниченного внутривидовой конкуренцией. Суть этой модели в том, что константа R в уравнении (7.60) заменена на фактическую скорость воспроизводства, т е. . Это уравнение представляет собой модель роста популяции, ограниченного внутривидовой конкуренцией. Суть этой модели в том, что константа R в уравнении (7.60) заменена на фактическую скорость воспроизводства, т е.  , которая уменьшается по мере роста численности популяции Nt. Достоинство полученного уравнения заключается в его простоте. Такой тип конкуренции приводит к саморегуляции численности популяции, изображенной на рис. 7.38 (для некоторого набора параметров модели; численное решение). , которая уменьшается по мере роста численности популяции Nt. Достоинство полученного уравнения заключается в его простоте. Такой тип конкуренции приводит к саморегуляции численности популяции, изображенной на рис. 7.38 (для некоторого набора параметров модели; численное решение). Рис. 7.38. Изменение численности популяции согласно уравнению (7.62) при R = 2, К = 200, N0 = 20 После несложного изменения в уравнении (7.62) может быть получена гораздо более общая модель, учитывающая интенсивность конкуренции. Простейшая из возможных зависимостей падения скорости роста популяции от ее численности, изображенная на рис. 7,37, является не законом природы, а всего лишь удобной гипотезой. Далеко не всегда реальная динамика численности популяции, определяемая внутривидовой конкуренцией, даже качественно согласуется с изображенной на рис. 7.38. Более общая гипотеза о законе падения скорости роста популяции в зависимости от ее численности приводит к следующему уравнению: Общность данной модели в отличие от уравнения (7.62) обусловлена введением в модель параметра b, который определяет тип зависимости падения скорости роста популяции от ее численности. Набор величин a, b, R можно использовать для сравнения и противопоставления сильно различающихся ситуаций. Другим положительным качеством уравнения (7.63) является его способность освещать новые стороны реального мира. Путем анализа кривых динамики популяций, полученных с помощью уравнения, можно прийти к предварительным выводам относительно динамики природных популяций. На рис. 7.39, а, б, в и г, построенных с помощью численного моделирования, показаны различные варианты динамики численности популяций, полученные с помощью уравнения (7.63) при разном сочетании параметров b и R. 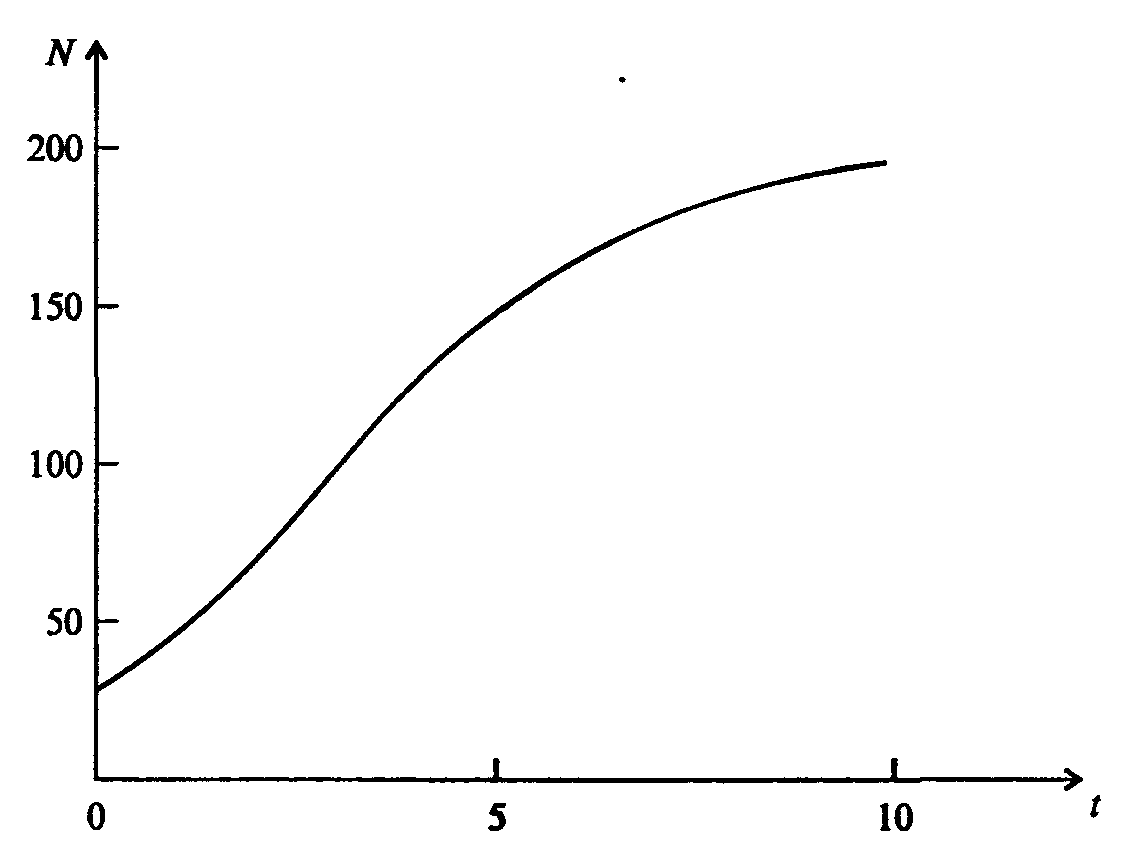 Рис. 7.39, а. Монотонное установление стационарной численности популяции при b =1,4, R =2 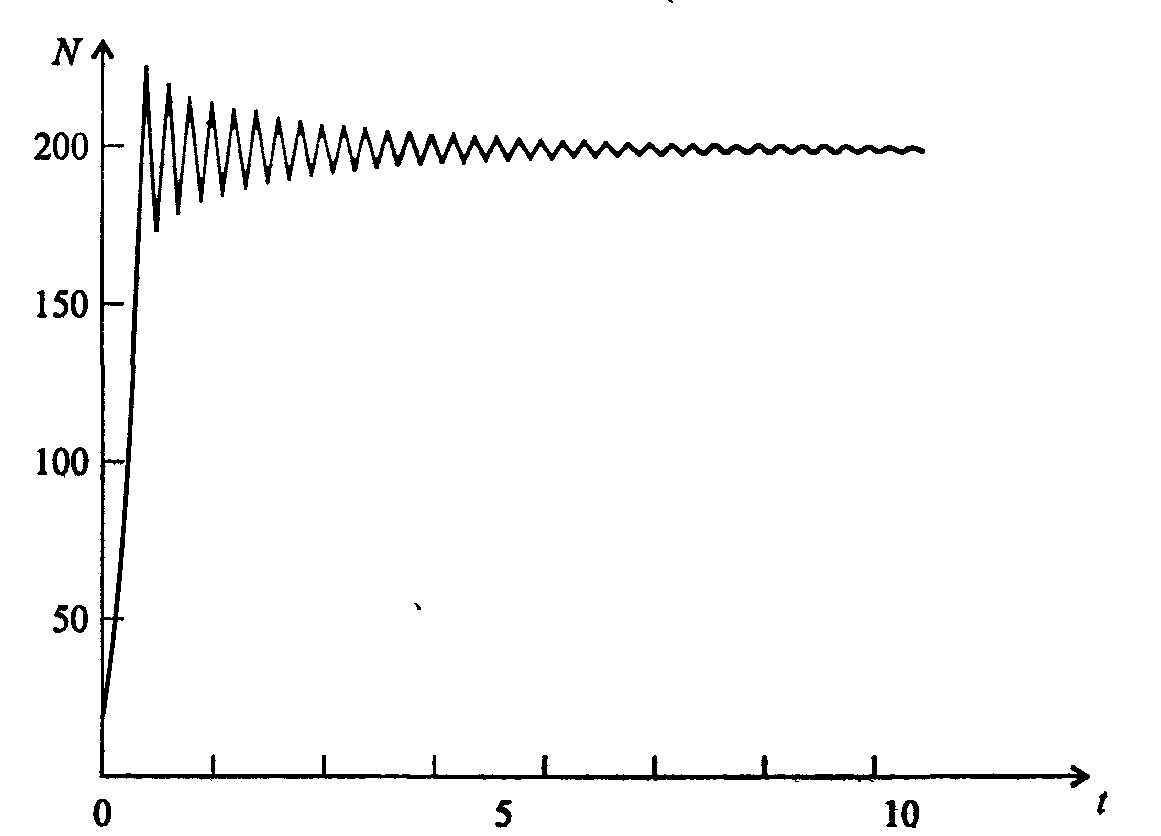 Рис. 7.39, б. Колебательное установление стационарной численности популяции при b =3,9, R =2 Важной частью исследования, связанного с моделью (7.63), является построение на фазовой плоскости (b, R) границ, которые разделяют монотонное затухание, затухающие колебания, устойчивые предельные циклы и случайные (хаотические) изменения, рис. 7.40 Для этого надо задаться значениями а и N0 и производить расчеты, изменяя параметры b, R. Различить каждый из возможных режимов можно попытаться визуально, выполняя построение на экране компьютера графиков изменения численности популяции и запоминая соответствующие значения параметров b, R при переходе от одного режима к другому. Следует, однако, понимать,  Рис. 7.39, в. Устойчивые предельные циклы изменения численности популяции при b = 3,6, R = 4 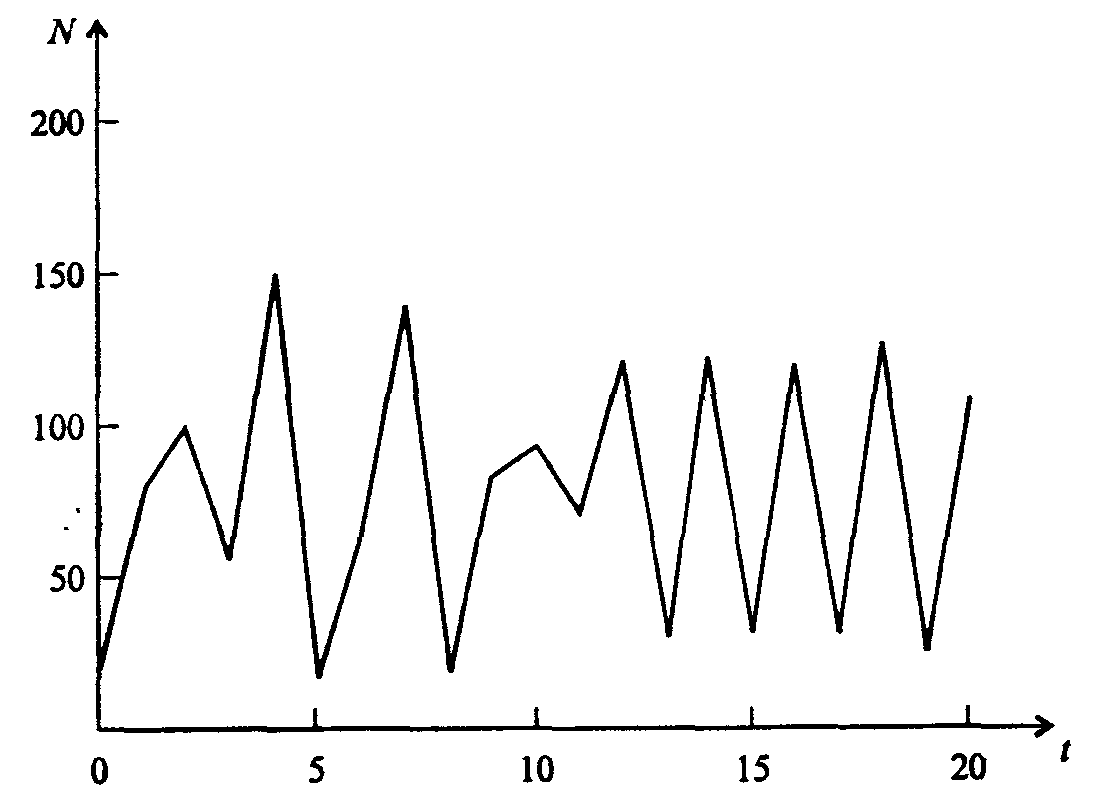 Рис. 7.39, г. Случайные изменения численности популяции при b = 4,4, R = 4 что установление различии между квазипериодическими, апериодическими и хаотическими движениями - сложная математическая задача. Для ее решения используются методы преобразований Фурье и другие, рассмотрение которых выходит за пределы данной книги. В связи с уравнениями (7.62), (7.63) сделаем замечание общего характера, справедливое для всех моделей в экологии. В отличие от физики, где «модель» часто является точно установленным законом природы, в экологии модель гораздо более условна. Ее адекватность реальности (в той мере, в какой моделирование вообще на это претендует) проверяется лишь экспериментально. Поскольку экологи много раз наблюдали каждую из динамик, изображенных на рис. 7.39, можно сделать вывод о полезности соответствующей модели (иначе она была бы лишь математическим упражнением). 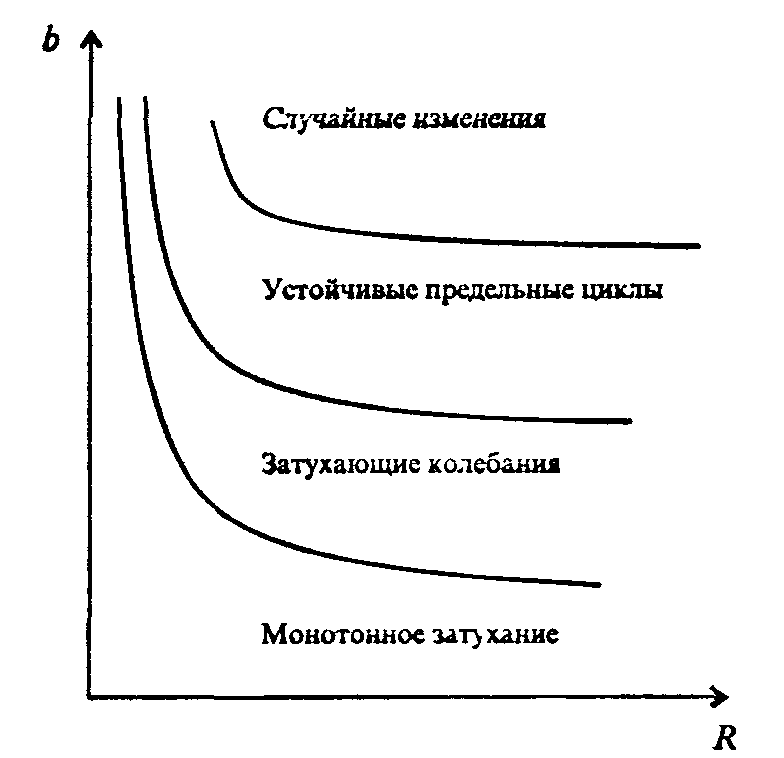 Рис. 7.40. Схематическое изображение фазовой диаграммы динамики численности популяции с дискретным размножением До сих пор мы рассматривали модели, применимые к популяциям с дискретными периодами размножения. Но в природе встречаются и популяции, где рождение и гибель организмов происходит непрерывно; для таких популяции модели, выраженные уравнениями из предыдущего пункта, непригодны. Рассмотрим популяцию с непрерывным размножением и построим модель изменения ее численности. Математическим аппаратом здесь являются дифференциальные уравнения. Скорость роста в этом случае можно обозначить 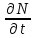 , тогда средняя скорость увеличения численности в расчете на одну особь определяется величиной , тогда средняя скорость увеличения численности в расчете на одну особь определяется величиной 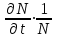 . . Без учета внутривидовои конкуренции получаем = r или Через r обозначена мгновенная удельная скорость роста численности, т.е. приращение численности за единицу времени в пересчете на одну особь. Согласие этой модели, вследствие того, что не учтена внутривидовая конкуренция, численность популяции при r > 0 будет неограниченно возрастать, т.е. будет наблюдаться экспоненциальный рост. Теперь попробуем учесть внутривидовую конкуренцию. Для этого воспользуемся методом, который однажды уже применяли (см. рис. 7.37). Обратимся к рис. 7.41. Когда численность популяции близка к нулю, скорость роста определяется величиной r, так как конкуренция еще не оказывает влияния на прирост популяции (точка А). Когда же при возрастании N достигается значение K (предельной плотности насыщения), скорость роста популяции снижается до нуля (точка В). Записав уравнение прямой АВ, получим 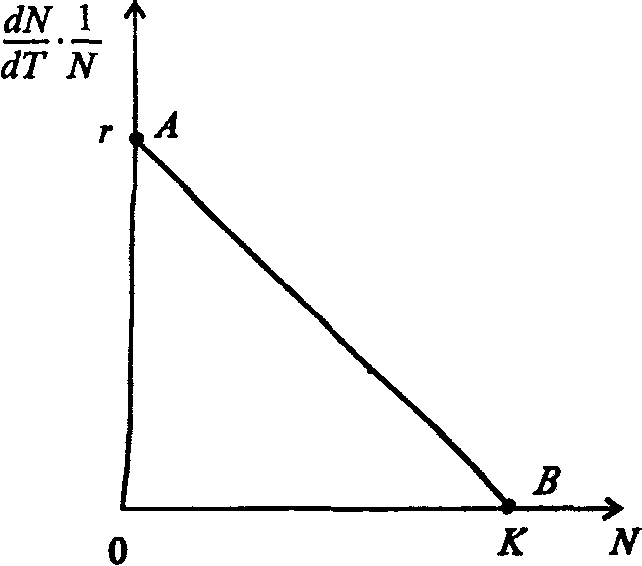 Рис. 7.41. Зависимость скорости воспроизводства от численности популяции Последнее уравнение известно под названием «логистического». Одно из его достоинств - простота. Уравнение относится к одному из стандартных типов и допускает аналитическое решение: В истории экологии логистическое уравнение сыграло значительную роль, оказав большое влияние на применение в экологии математических методов. С другой стороны, простота уравнения накладывает ограничения на область его применения, так как с его помощью можно описать немногие реальные популяции. Но, несмотря на все ограничения, логистическое уравнение используется как составная часть и в других моделях динамики численности популяции, рассматриваемых ниже. Изменение численности популяции в соответствии с формулами (7.65)-(7.66) иллюстрирует рис. 7.42. 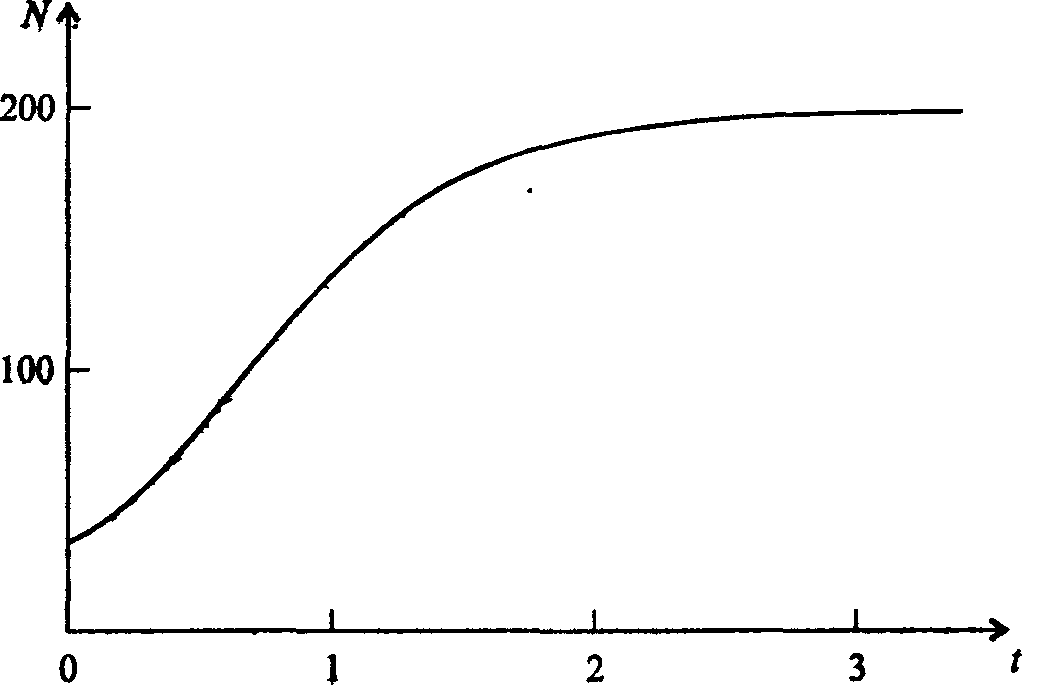 Рис. 7.42. Динамика численности популяции при No =25, К = 200, r = 3 4.3. ЛОГИСТИЧЕСКАЯ МОДЕЛЬ МЕЖВИДОВОЙ КОНКУРЕНЦИИСущность межвидовой конкуренции заключается в том, что у особей одного вида уменьшается плодовитость, выживаемость и скорость роста в результате использования того же ресурса, что и особями другого вида, причем влиянию конкуренции в той или иной мере подвергаются особи обоих видов. Так как мы имеем дело с двумя различными популяциями, то введем соответствующие обозначения. Пусть N1 - численность первой популяции, a N2 - второй. Предельные плотности насыщения и максимальные врожденные скорости роста популяций обозначим соответственно K1, К2, r1 и r2, рис. 7.43. Обратимся к логистическому уравнению (7.65) и попробуем учесть в нем межвидовую конкуренцию. Сделаем это так. Предположим, что М особей вида 2 оказывают такое же воздействие на вид 1, как одна особь вида 1. Константу 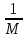 в этом случае называют коэффициентом конкуренции. Обозначим ее α12. Таким образом, чтобы отразить суммарное воздействие на вид 1, надо в логистическом уравнении в числителе дроби вместо N записать N1 + α12∙N2. Аналогично получается уравнение для исследовании численности второй популяции. В результате получаем систему двух дифференциальных уравнений: в этом случае называют коэффициентом конкуренции. Обозначим ее α12. Таким образом, чтобы отразить суммарное воздействие на вид 1, надо в логистическом уравнении в числителе дроби вместо N записать N1 + α12∙N2. Аналогично получается уравнение для исследовании численности второй популяции. В результате получаем систему двух дифференциальных уравнений: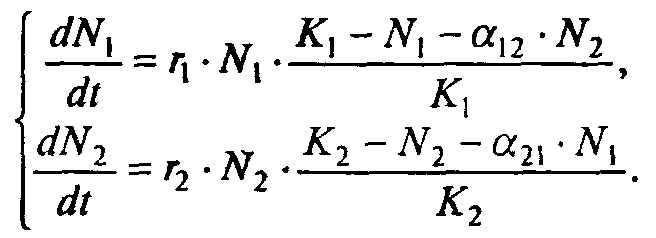 (7.67) (7.67)Модель межвидовой конкуренции, выраженная этой системой» названа в честь ее авторов «моделью Лотки-Вольтерры». Заметим, что если коэффициенты α12 или α21больше единицы, то влияние со стороны конкурирующей популяции на особей данного вида сильнее, чем со стороны особей своего вида. 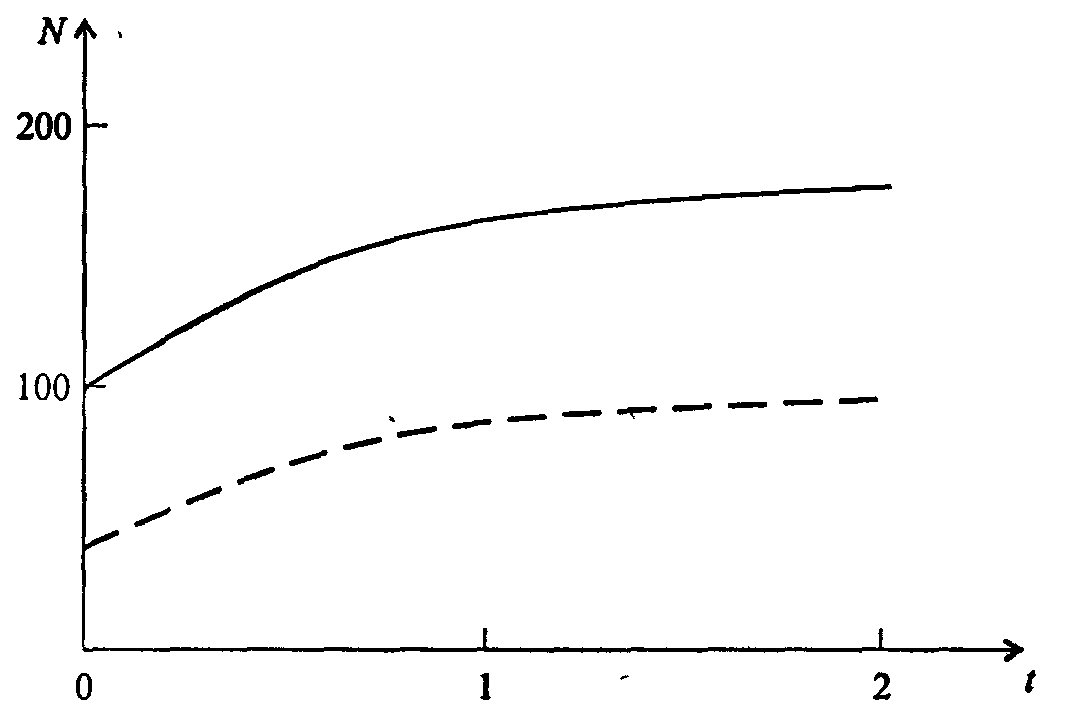 Рис. 7.43. Устойчивое сосуществование популяций при r1 = 2, r2 = 4, К1 = 200, К2 = 180, α12 = 0,5, α21 = 0,65, N  = 100, N = 100, N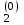 = 25. Устойчивое сосуществование достигается лишь при α12∙ α21 < 1. Сплошная линия - численность первой популяции, штриховая – второй = 25. Устойчивое сосуществование достигается лишь при α12∙ α21 < 1. Сплошная линия - численность первой популяции, штриховая – второйГлавный вопрос, который интересует исследователя межвидовой конкуренции -при каких условиях увеличивается или уменьшается численность каждого вида? Для ответа на этот вопрос надо построить диаграммы, где были бы изображены все возможные сочетания численностеи обоих видов. На таких диаграммах численность одного вида откладывают по горизонтальной оси, а другого - по вертикальной. При одних сочетаниях численностеи будет отмечаться рост выбранной для наблюдения популяции, при других - уменьшение ее численности. Также для каждого из видов можно провести изоклины - линии, вдоль которых не наблюдается ни увеличения, ни уменьшения численности. Рассмотрим, как можно построить изоклину для первого вида. По определению, для этой линии = 0. Из первого уравнения системы (7.67) получаемЭто равенство выполняется, если какой-либо из множителей равен нулю. Наибольший интерес представляет ситуация, в которой или 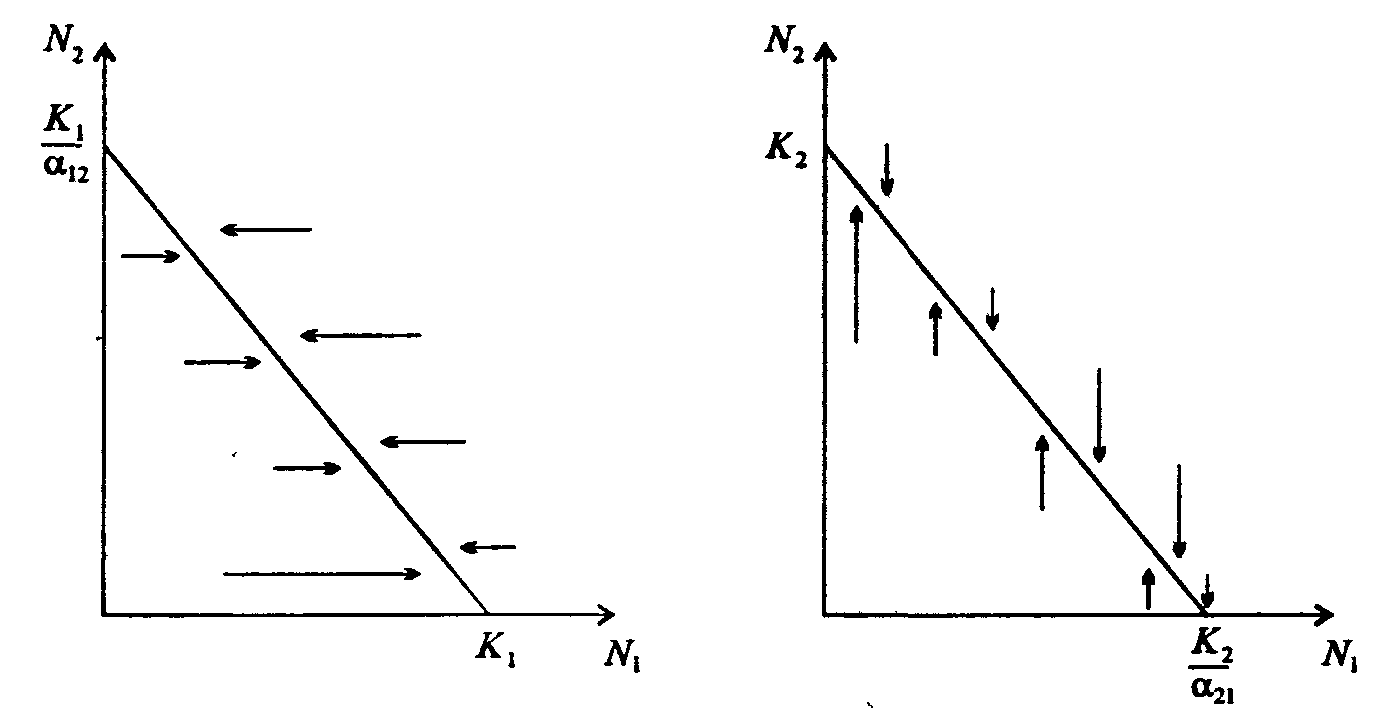 Рис. 7.44. Изоклины, полученные с помощью модели Лотки-Вольтерры. Длины стрелок пропорциональны изменению численности, стрелки указывают направление изменения численности Таким образом, получено уравнение изоклины, которое, как можно заметить, является уравнением прямой в плоскости (N1, N2). Вверх и вправо от изоклины из-за высокой численности обеих популяций численность вида 1 снижается, в противоположных направлениях - повышается. Аналогично можно построить изоклину для вида 2. На рис. 7.44 построены соответствующие изоклины, и показано изменение численности популяций.  Рис. 7.45. Результаты конкуренции, полученные с помощью модели Лотки-Вольтерры при различных параметрах. На рисунке а в зоне I численность обеих популяций падает; в зоне II - численность первой популяции растет, второй - уменьшается; в зоне 111 - численность обеих популяций увеличивается Таким образом, получено уравнение изоклины, которое, как можно заметить, является уравнением прямой в плоскости (N1, N2). Вверх и вправо от изоклины из-за высокой численности обеих популяций численность вида 1 снижается, в противоположных направлениях - повышается. Аналогично можно построить изоклину для вида 2. На рис. 7.44 построены соответствующие изоклины и показано изменение численности популяций. Для решения поставленной выше задачи объединим в одной фазовой плоскости изоклины для обоих видов и будем одновременно исследовать динамику их численности. Изоклины относительно друг друга располагаются четырьмя различными способами, что дает различный исход конкуренции. На рис. 7.45 представлены результаты конкуренции, полученные с помощью системы уравнений (7.67), заимствованные из книги М. Бигона и др. «Экология». 4.4. ДИНАМИКА ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ ХИЩНИКА И ЖЕРТВЫРассматривая динамику численности популяций хищника и жертвы, экологи прежде всего стремятся понять ее закономерности и разъяснить различия между типами динамик. В простейших моделях хищник и жертва рассматриваются безотносительно влияния на них других видов. Одна из самых первых и простых моделей была предложена, как и модель межвидовой конкуренции, Лоткой и Вольтеррой, и носит их имя. Модель состоит из двух компонентов: С - численность популяции хищника и N -численность популяции жертвы. Предполагается, что в отсутствие хищника популяция жертвы растет экспоненциально. Чем больше численность той и другой популяции, тем чаще происходят встречи. Число встреченных и съеденных жертв будет зависеть от эффективности, с которой хищник находит и ловит жертву. Если обозначить через а' «эффективность поиска», то скорость поедания жертвы будет равна a'∙C∙N, и окончательно для численности жертвы получаем В отсутствие пищи отдельные особи хищника голодают и гибнут. Предположим вновь, что численность хищника в отсутствие пищи будет уменьшаться экспоненциально: (q - смертность). Скорость рождения новых особей в данной модели полагается зависящей от двух обстоятельств: скорости потребления пищи a'∙C∙N, и эффективности f, с которой эта пища переходит в потомство хищника. Итак, для численности хищника окончательно получаем Так как процессы надо рассматривать вместе, объединим уравнения в систему:  (7.68) (7.68)Как и в предыдущем пункте, свойства этой модели можно исследовать, построив изоклины. Для жертвы имеем или, выражая С, получаем Соответствующее уравнение изоклины для популяции хищника Если поместить обе изоклины на одном рисунке, получим картину взаимодействия популяций (рис. 7.46). Как видно на рис. 7.47, численности популяций хищника и жертвы совершают периодические колебания: при увеличении численности хищников уменьшается 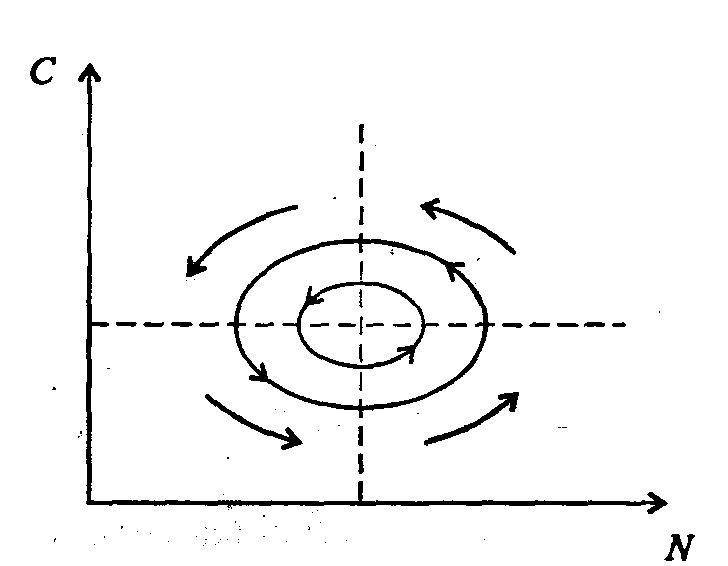 Рис. 7.46. Динамика численности популяции хищника и жертвы. Численность обеих популяций совершает периодические колебания численность популяции жертвы и наоборот. Такие колебания численности будут продолжаться в соответствии с моделью до тех пор, пока какое-либо внешнее воздействие не изменит численность популяций, после чего произойдет переход в новое устойчивое состояние (такая ситуация называется «нейтральные устойчивые циклы»). 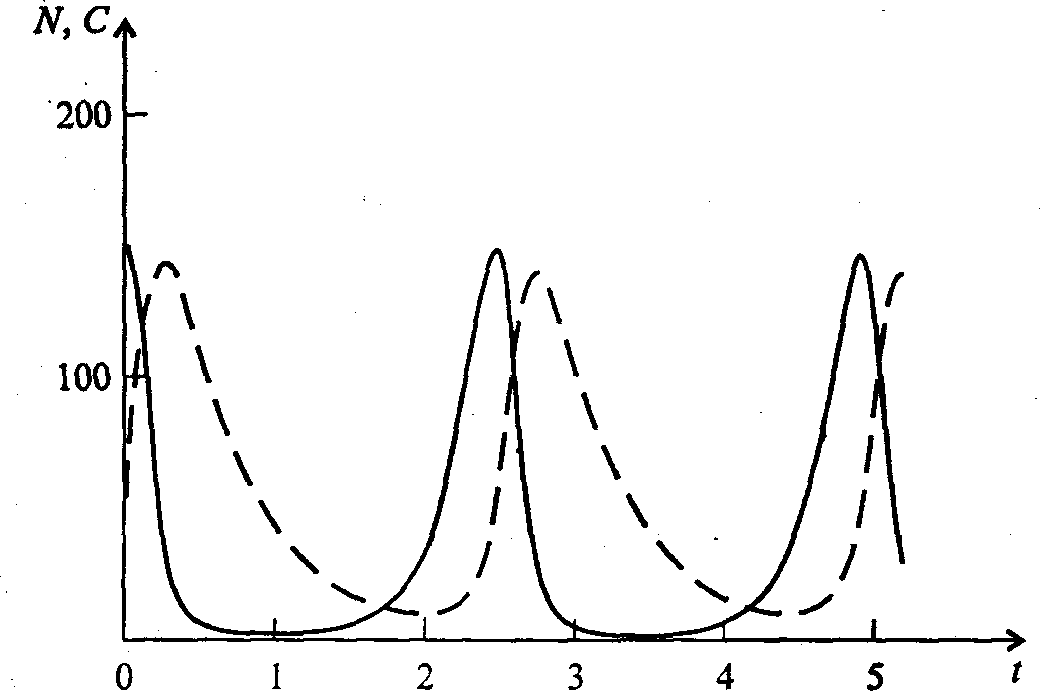 Рис. 7.47. Динамика численности популяции хищника и жертвы при r = 5, а' = 0,1, q = 2, f= 0,6, N0 = 150, C0 = 50. Сплошная линия - численность жертвы, штриховая – хищника 4.5. ИМИТАЦИОННОЕ МОДЕЛИРОВАНИЕ ДИНАМИКИ ПОПУЛЯЦИЙВыше мы обсудили несколько классических математических моделей в экологии. Известно и немало других, которые можно найти в книгах и в статьях научно-популярных журналов. Использование математических моделей, основанных на дифференциальных уравнениях или их дискретных аналогах, далеко не единственный путь в компьютерном моделировании динамики популяций. Другую интересную группу составляют чисто стохастические модели или динамические модели с элементами стохастичности. Опишем две возможные модели такого класса: одну - без подробностей, которые рекомендуем продумать самостоятельно, другую - во всех деталях, включая реализующую ее программу. Первая модель. Две популяции находятся в соотношении «хищник - жертва», описанном выше, но модель совершенно иная. На некотором клетчатом поле (которое наглядно - все или по частям - изображается на дисплее) находятся волки (несколько) и зайцы (много). Зайцы совершают беспорядочные перемещения, длины и направления которых - случайные величины, подчиняющиеся некоторому закону распределения вероятностей, а каждый волк в каждое мгновение ориентируется на ближайшего зайца и может совершать ограниченные прыжки. Если волк в течение некоторого, заранее определенного, промежутка времени остается без добычи (т.е. не попадает в клетку, где находится заяц), то он погибает. Зайцы и волки через определенные моменты размножаются, например, каждый второй заяц приносит двух потомков, каждый второй волк приносит одного потомка, причем скорость размножения волков в два раза ниже, чем зайцев. Все это может красочно изображаться на экране, состояние которого меняется через фиксированные моменты времени. Это - чисто имитационное моделирование, конечной целью которого является, как и в описанных моделях, установление судьбы популяций в зависимости от многих факторов, входящих в модель. Здесь нет дифференциальных уравнений, но зато в полной мере проявляется стохастика, моделирование случайных процессов. Реализация такой модели требует большого программистского искусства, способствует формированию вероятностных представлений. Возможны и пограничные - динамико-стохастические модели. Например, в уравнениях Лотки - Вольтерры один из параметров - случайная величина с известным законом распределения. Самостоятельные размышления над такими задачами, даже лишь над их постановкой, очень полезны. Вторая модель, которую опишем во всех деталях, предложена Д.Конвейем -имитационная модель роста, распада и различных изменений в популяции живых организмов, известная под названием «Жизнь».  Рис. 7.48. К имитационной модели «Жизнь» Для построения алгоритма игры рассмотрим квадратное поле из n+1 столбцов и строк с обычной нумерацией от 0 до n, рис. 7.48. Крайние граничные столбцы и строки для удобства определим как «мертвую зону», они играют лишь вспомогательную роль. Для любой внутренней клетки поля с координатами (i, j) можто определить 8 соседей. Если клетка «живая», ее закрашиваем, если клетка «мертвая», она пустая. Зададим правила игры. Если клетка (i, j) «живая» и в окружении более трех «живых» клеток, она погибает (от перенаселения). «Живая» клетка также погибает, если в окружении менее двух «живых» клеток (от одиночества). «Мертвая» клетка оживает, если вокруг нее имеются три «живые» клетки. Для удобства введем двумерный массив А[0..п, 0..п], элементы которого принимают значение 0, если соответствующая клетка пустая, и 1, если клетка «живая». Тогда алгоритм определения состояния клетки с координатой (i,j) можно определить следующим образом: S := A[I-1, J-1] + A[I-1, J] + A[I-1, J+1] + A[I+1, J-1] + А[I+1, J] + A[I+1, J+l] + A[I, J+l] + А[1, J-1]; If (A[I, J] = 1) And ((S > 3) Or (S < 2); Then B(I, J] := 0; If (A[I, J] = 0) And (S = 3) Then B[I, J] := 1; Здесь массив В [0..п, 0..п] определяет координаты поля на следующем этапе. Для всех внутренних клеток от i = 1 до n- 1 и j = 1 до n- 1 справедливо сказанное выше. Отметим, что последующие поколения определяются аналогично, лишь стоит осуществить процедуру переприсваивания For I := 1 То N-l DO For J := 1 To N-l Do A[I, J] := B[I, J] ; На экране дисплея удобнее выводить состояние поля не в матричном, а в графическом виде. Читатель без труда это может осуществить. Осталось лишь определить процедуру задания начальной конфигурации игрового поля. При случайном определении начального состояния клеток подходит алгоритм For I := 1 То К Do Begin Kl := Random(N-l); K2 := Random(N-l)+1; A[K1, K2] := 1 End; Интереснее случай, когда начальную конфигурацию мы задаем сами, и удобнее для пользователя делать это непосредственно в графическом виде. Полный текст подобной программы приведен ниже. Программа 150. «Жизнь». Program PlayLife; Uses Crt, Graph; Const R = 41; Var P ; 0..1; А, В : Array[0..R, 0..R] Of 0..1; I, J, N, Kl, K2, Ss, X, Y, Gm, Gd : Integer; С : Char; Begin DetectGraph(I, J); InitGraph(I, J, "); { -- заставка -- ) For I := 0 To 5 Do Begin SetFillStyle (1, I+2); Bar(180+1*10, 100+1*10, 430-1*10, 270-1*10); End; OutTextXY(260, 180, 'ИГРА "ЖИЗНЬ"'); Repeat Until KeyPressed; { - начальная конфигурация - } RestoreCrtMode ; For I := 0 To R Do For J := 0 To R Do Begin A[I, J] := 0; В[I, J] := 0 End; CIrScr; Write('Начальное поселение будете задавать случайно или вводить (0/1)?'); ReadLn(P); If P = 0 Then Begin Write('Введите начальное количество поселений 2 < n < 200: '); ReadLn(N); Gm := VgaHi; SetGraphMode(Gm) ; For I := 1 To N Do Begin Kl := Random(R - 1) + 1; K2 := Random(R - 1) + 1; A{K1, K2] := 1; B[K1, K2] := 1; End; SetLineStyle(0, 0, 1); SetColor(4); For I := 0 To R - 1 Do Begin Line(10 + I * 10, 10, 10 + I * 10, R * 10); Line(10, 10 + I * 10, R * 10, 10 + I * 10); End; For I := 1 To R - 1 Do For J := 1 To R - 1 Do If A[I, J] = 1 Then FloodFill(5 + I * 10, 5 + J * 10, 4); Repeat Until KeyPressed; End Else Begin Gm := VgaHi; SetGraphMode(Gm); SetLineStyle(0, 0, 1); SetColor(4); For I := 0 To R - 1 Do Begin Line(10 + I * 10, 10, 10 + I * 10, R * 10); Line(10, 10 + I * 10, R * 10, 10 + I * 10); End; X := 15; Y := 15; Repeat С := ReadKey; If С = #0 Then С := ReadKey; Case C Of #72 : Begin SetColor(O); Circle(X, Y, 2); Y := Y - 10; SetColor(15); Circle(X, Y, 2) End; #80 : Begin SetColor(O); Circle(X, Y, 2); Y := У + 10; SecColor(15); Circle(X, Y, 2) End; #75 : Begin SetColor(O); Circle(X, Y, 2); X := X - 10; SetColor(15); Circle(X, Y, 2) End; #77 : Begin SetColor(0); Circle(X, Y, 2); X ^ X + 10; SetColor(15) ; Circle(X, Y, 2) End; #13 : Begin FloodFill(X, Y, 4); A[(X-5) Div 10, (Y-5) Div 10] := 1; B[(X - 5) Div 10, (Y - 5) Div 10] := 1 End; #32 : Begin SetFillStyled, 0); Bar(X - 4, Y - 4, X + 4, Y + 4); A[(X - 5) Div 10, (Y - 5) Div 10] := 0: B[(X - 5) Div 10, (Y - 5) Div 10] := 0; SetFillStyle(1, 15); End; End {case}; Until С = #27; SetColor(0); Circle(X, Y, 2) ; End; SetColor(15); Repeat Until Key Pressed; OutTextXY(GetMaxX - 400, GetMaxY - 5, 'для продолжения –любая клавиша, выход - s'); {- начало игры -- } Repeat For I := 1 То R - 1 Do For J := 1 To R - 1 Do Begin Ss : = A[I - 1, J - 1] + A[I - 1, J] + A[I - 1, J + 1]; Ss := Ss + A(I + 1, J - 1] + A[I + 1, J] + A[I +1, J + 1] ; Ss := Ss + A[I. J + 1] + A[I, J - 1]; If (A[I, J] = 1) And (Ss > 3) Then Begin SetFillStyled, 0); Bar(l + I * 10, 1 + J * 10, 9 + I * 10, 9 + J * 10); B(I, J] := 0; SetFillStyled, 15) End; If (A[I, J] == 1) And (Ss < 2) Then Begin SetFillStyled, 0); Bar(l + I * 10, 1 + J * 10, 9 + I * 10, 9 + J * 10); B[I, J] := 0; SetFillStyled, 15) End; If (A[I, J] = 0) And (Ss = 3) Then Begin FloodFill(5 + I * 10, 5 + J * 10, 4); B[I, J] := 1 End; Delay(20) End; For I := 1 To R - 1 Do For J := 1 To R - 1 Do A[I, J] ••= B[I, J] ; С := ReadKey Until С = 's'; CloseGraph End. Теперь приступим к моделированию. Поставим несколько вопросов: какие конфигурации исчезают с течением времени и как быстро? какие структуры могут существовать бесконечно? каковы законы организации структур, их взаимодействия? Перед попыткой ответить на эти вопросы читателю будет полезно поэкспериментировать самому. Предварительно отметим лишь некоторые структуры (рис. 7.49-7.50).  Рис. 7.49. Стационарные структуры, не зависящие от времени Существуют структуры, которые повторяют себя через 1, 2, ...,п временных шагов, их называют циклами «-n» (см. рис. 7.50).  Рис. 7.50. Циклы «-2» Но наиболее интересны движущиеся структуры - «планеры», «корабли», «паровозы», их столкновения, «аннигиляции» и зарождение новых структур. Представляет интерес модель игры в одномерном случае. Здесь имеется один ряд клеток. В некоторых нз них задаем «жизнь». Требуется изучить эволюцию клеток, если заданы правила их существования и зарождения. Определим сумму значений пяти соседних клеток (самой клетки и двух ближайших к ней слева и справа). На следующем шаге в зависимости от суммы, которая может быть равна 0,1, 2, 3,4, 5, клеткам присваивается значение 0,1,0,0,0,0 или 0, 1,1, 1,0,0. Пусть X(N) - массив клеток. X(i) = 0, если i-я клетка пустая, и X(i) = 1, если в ('-и клетке имеется «жизнь». Если Y(N) - массив клеток на следующем временном шаге, то он заполняется, например, по следующему правилу - если S = 1, то Y(i) = 1, иначе Y(i) = 0. Программа 7.6. «Жизнь на полоске». Program Lifel; Uses Crt, Graph; Const N = 60; Var J, К : Integer; X, Y : Array(O..N] Of 0..1; I, S, Yg : Integer; P : Real; С : Chart; Begin For I := 0 To N Do Begin X[I] := 0; Y[I] := 0 End; (начальное поселение) X[N Div 2] := 1; DetectGraph(J, К); InitGraph(J, К, "); Yg := 20; For I := 0 To N Do If X(I] = 1 Then Circled * 10 + 10, Yg, 2); Repeat С := ReadKey; Yg := Yg + 10; For I := 2 To N - 2 Do Begin S ;= X[I - 2] + X[I - 1] + X[I] + X[I + 1] + X[I + 2]; If S In [1..3] Then Y[I1 := 1 End; For I := 0 To N Do Begin If Y[I] = 1 Then Circle(I * 10 + 10, Yg, 2); X[I] := Y[I]; Y[I] := 0 End Until С = #27; CloseGraph; End. |