биология экзамен 1 курс. Шпора. Экзамен по биологии. 1. Биология. Жизнь, происхождение. Уровни организации жизни. Термин биология
 Скачать 0.87 Mb. Скачать 0.87 Mb.
|

|
Задача 6-23 Гены А, В и С находятся в одной группе сцепления. Между генами А и В кроссинговер происходит с частотой 7,4%, а между генамиВ и С – с частотой 2,9%. Определить взаиморасположение генов А, В и С, если расстояние между генами А и С равняется 10,3% единиц кроссинговера. Как изменится взаиморасположение этих генов, если частота кроссинговера между генами А и С будет составлять 4,5%? Решение По условию задачи расстояние от гена А до гена С (10,3 М) равно сумме расстояний между генами А и В (2,9 М) и генами В иС (7,4 М), следовательно, ген В располагается между генами А и С и расположение генов следующее: А В С. Если бы расстояние от гена А до гена С равнялось разности расстояний между парами генов АВ и ВС (4,5 = 7,4 – 2,9), то гены располагались бы в следующей последовательности: А С В. И в этом случае расстояние между крайними генами было бы равно сумме расстояний между промежуточными: АВ = АС + СВ. Задача 6-24 При анализирующем скрещивании тригетерозиготы АаВbСс были получены организмы, соответствующие следующим типам гамет:
Решение Расщепление при анализирующем скрещивании, близкое к 1:1, указывает на то, что все три пары генов находятся в одной хромосоме. Расстояние между генами А и В равно: 1,7 + 1,7 = 3,4 М. Расстояние между генами В и С равно: 0,8 + 0,8 = 1,6 М. Ген В находится между генами А и С. Расстояние между генами А и С равно: 1,7 + 1,7 + 0,8 + 0,8 = 5,0 М. Карта участка хромосомы: 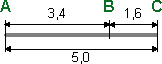 30.Соотносительное наследование признаков - независимое и сцепленное. Независимое наследование — Наследование определенного гена (признака) без влияния иных генетических факторов (др. определенного гена, пола); Независимое наследование признаков. закон независимого наследования признаков: «Разные пары признаков, определяемые неаллельными генами, передаются потомкам независимо друг от друга и комбинируются у них во всех возможных сочетаниях». этому закону должны подчиняться в первую очередь неаллельные гены, располагающиеся в разных (негомологичных) хромосомах. В таком случае независимый характер наследования признаков объясняется закономерностями поведения негомологичных хромосом в мейозе. Названные хромосомы образуют со своими гомологами разные пары, или биваленты, которые в метафазе I мейоза случайно выстраиваются в плоскости экватора веретена деления. Затем в анафазе I мейоза гомологи каждой пары расходятся к разным полюсам веретена независимо от других пар. В результате у каждого из полюсов возникают случайные сочетания отцовских и материнских хромосом в гаплоидном наборе. Следовательно, различные гаметы содержат разные комбинации отцовских и материнских аллелей неаллельных генов. Разнообразие типов гамет, образуемых организмом, определяется степенью его гетерозиготности и выражается формулой 2n, где n — число локусов в гетерозиготном состоянии. В связи с этим дигетерозиготные гибриды F1 образуют четыре типа гамет с одинаковой вероятностью. Реализация всех возможных встреч этих гамет при оплодотворении приводит к появлению в F2 четырех фенотипических групп потомков в соотношении 9:3:3:1. Сцепление генов — это совместное наследование генов, расположенных в одной и той же хромосоме. Сцепленное наследование признаков. У потомков такого скрещивания вместо свободного комбинирования признаков разных пар наблюдали тенденцию к наследованию преимущественно родительских сочетаний признаков. Такое наследование признаков было названо сцепленным. Сцепленное наследование объясняется расположением соответствующих генов в одной и той же хромосоме. В составе последней они передаются из поколения в поколение клеток и организмов, сохраняя сочетание аллелей родителей. Гомологичные хромосомы — это одинаковые группы сцепления, которые отличаются друг от друга лишь аллелями отдельных генов. При конъюгации гомологи сближаются своими аллельными генами, а при кроссинговере они обмениваются соответствующими участками. В результате появляются кроссоверные хромосомы с новым набором аллелей. Частота, с которой происходит обмен на участке между двумя данными генами, зависит от расстояния между ними (правило Т. Моргана). Процент кроссоверных гамет, несущих кроссоверные хромосомы, косвенно отражает расстояние между генами. Нарушение сцепленного наследования родительских аллелей в результате кроссинговера позволяет говорить о неполном сцеплении в отличие от полного сцепления, наблюдаемого, например, у самцов дрозофилы. 31. Хромосомные мутации. Классификация. Возможные причины возникновения и последствия хромосомных мутаций. Примеры у человека. Хромосомные перестройки называются хромосомными мутациями или аберрациями. Они являются инструментом комбинативной генотипической изменчивости. Заключаются в сокращении или увеличении числа определенных генов (изменении дозы генов), перераспределении генов между хромосомами, изменении положения генов в хромосоме. 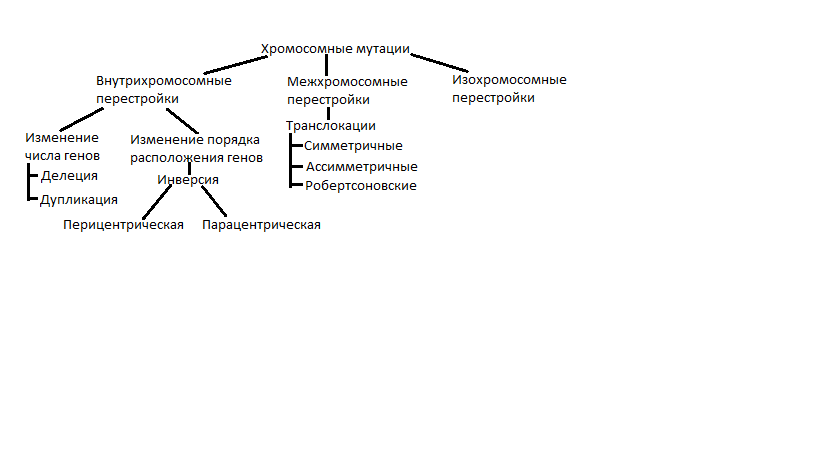 Делеция – потеря участка хромосомы. Делеция – потеря участка хромосомы.Дупликация – удвоения участка хромосомы. Инверсия – поворот участка на 1800. Если участок включает центромеру – перицентрическая инверсия, если нет – парацентрическая. Транслокация заключается в обмене участками хромосом. Если две негомологичные хромосомы обмениваются участками, говорят о реципрокной транслокации (симметричной), а если оторвавшийся участок хромосомы присоединяется, но в другом месте, говорят о транспозиции (ассиметричной транслокации). Робертсоновская транслокация (рассматривается отдельно) – «объединение» двух негомологичных хромосом в одну, или образование из одной хромосомы двух самостоятельных. Причины возникновения: нарушение хода кроссинговера, воздействие мутагенов. Последствия. Чаще структурные перестройки хромосом соматических клеток сказываются на их жизнеспособности отрицательно (соматические мутации). Указывают на возможность малигнизации. Генеративные хромосомные мутации приводят к невозможности развития особи, получившей неполноценный наследственный материал. Однако, в тех случаях, когда такие мутации оказываются совместимыми с жизнью, они способствуют биологической эволюции (видообразованию). Примеры: 1.Синдром Дауна (в том случае, когда лишняя 21-я хромосома целиком транслоцирована на другую). 2. Синдром «кошачьего крика» (возникает оно из-за частичной моносомии по короткому плечу хромосомы 5, обусловленной делецией). У большей части страдающих этим синдромом отмечается изменение строения черепа: уменьшение мозгового отдела, лунообразная форма лица. Ушные раковины при синдроме «кошачьего крика» обычно расположены низко. Иногда у больных отмечаются врожденные патологии сердца или других органов. Характерным признаком также становится умственная отсталость. Дети, страдающие этой болезнью, имеют характерный плач, который напоминает кошачье мяуканье. 32. Генные мутации. Классификация. Мутон. Возможные причины возникновения и последствия генных мутаций. Примеры у человека. Не устранённые и/или неисправленные изменения химической структуры генов, воспроизводимые в последующих циклах репликации и повторяющиеся у потомков в виде измененных вариантов признака, называют генными мутациями. Делят на 3 группы: Точечные мутации (замена одного нуклеотида на другой). Не всегда ведут к изменению смысла ген. информации. К ним относятся миссенс, нонсенс, сайленс (нейтральные мутации). Миссенс – изменение АК в белке. Сайленс – АК не меняются (благодаря вырожденности генетического кода). Нонсенс – вместо кодона для АК появляется стоп-сигнал. Сдвиг рамки считывания. Сюда относятся инсерции (дупликации) – удвоение участка генов, дефишенси (делеции) – потеря участка гена. Изменение порядка следования нуклеотидов в пределах гена (инверсия). Мутон – наименьшая единица мутационного процесса, представляющая собой отдельно взятый нуклеотид или пару нуклеотидов. Причины. Мутации бывают спонтанные (без видимой причины) и индуцированные (воздействие ионизирующего излучения, химических соединений, биологических агентов, н-р вирусов). Мутации по типу замены нуклеотидов происходят в силу разных причин. Одна из них – изменяется азотистое основание нуклеотида, уже встроенного в молекулу ДНК. Если это искажение не устраняется механизмами молекулярной репарации, то в результате репликаций оно становится достоянием обеих цепей ДНК. Т.о. нарушения процессов репликации и репарации – источник точечных мутаций. Генные мутации по типу сдвига «рамки считывания» чаще спонтанны. Однако их число возрастает при действии некоторых химических соединений, в частности акридиновых. Рентгеновские лучи могут вызывать делеции. Последствия. Большинство изменений фенотипически неблагоприятно – вредные генные мутации. Среди них выделяют летальные (несовместимы с жизнью) и полулетальные мутации (стерильные особи или не доживают до половой зрелости). Редко случаются изменения генов с благоприятными последствиями – полезные генные мутации. Известны нейтральные генные мутации, не сказывающиеся на жизнеспособности и репродуктивном потенциале. Примеры: Гемофилия Синдром Марфана – мутация в гене, ответственном за синтез белка соединительнотканных волокон фибриллина, приводящих к блоку его синтеза. Соединительная ткань обладает повышенной резистентностью, что выражается в нарушении мышечно-скелетной системы (арахнодактилия – высокий рост, длинные конечности, паучьи пальцы), поражение глазной системы (вывих хрусталика), поражению сердечно-сосудистой системы (митрального клапана, расслоение аорты). Альбинизм – болезнь обусловлена отсутствием синтеза фермента тирозиназы, в результате чего не синтезируется пигмент меланин. Характерна обесцвеченность кожи, волос и глаз, независимо от расы и возраста. Светобоязнь. Зрение снижено. Серповидно-клеточная анемия (гемоглобинопатия) – генная миссенс мутация, обусловленная заменой глютаминовой к-ты на валин. Эритроциты приобретают форму серпа. ФКУ (фенилкетонурия) 33. Множественный аллслизм. Причины, примеры у человека. Множественный аллелизм – это присутствие в генофонде вида более двух аллелей данного гена. У человека, как и у других представителей органического мира, множественный аллелизм свойствен многим генам. Так, три аллеля гена I определяют групповую принадлежность крови по системе АВ0 (IA, IB, I0). Два аллеля имеет ген, обусловливающий резус- принадлежность. Более ста аллелей насчитывают гены α- и β-полипептидов гемоглобина. Причиной множественного аллелизма являются случайные изменения структуры гена (мутации), сохраняемые в процессе естественного отбора в генофонде популяции. Многообразие аллелей, рекомбинирующихся при половом размножении, определяет степень генотипического разнообразия среди представителей данного вида, что имеет большое эволюционное значение, повышая жизнеспособность популяций в меняющихся условиях их существования. 34. Наследование групп крови по системам: ABO, MN и Rh-фактор. Резус-конфликт. Система АВ0. Группы крови системы АВ0 («а», «б», «ноль») контролируются одним аутосомным геном I или ABO, расположенным в длинном плече хромосомы 9. В этом гене идентифицировано 3 аллеля IA, IB и I0. Аллели IA и IB кодоминантны по отношению друг к другу, и оба они доминантны по отношению к аллелю I0. Таким образом, при сочетании различных аллелей могут образовываться 4 группы крови: 0 или I при генотипе I0I0, A или II при генотипах IAIA и IAI0, B или III при генотипах IBIB и IBI0 и AB или IV при генотипе IAIB в соотношении 1:3:3:2. Группы крови определяют иммунологические свойства антигена агглютиногена, локализованного на поверхности эритроцитов, и взаимодействующего с ними антитела агглютинина, растворенного в сыворотке крови. При самой редкой группе крови 0(I), которая в популяции встречается с частотой 11% (1:9), в сыворотке крови вырабатываются антитела против антигенов А и В. Если человеку с группой крови 0(I) добавить кровь любой другой группы произойдет агглютинация (слипание) эритроцитов и разовьется гемолитический шок. В тоже время кровь группы 0(I) не содержит эритроцитарных антигенов, и ее можно переливать любым реципиентам вне зависимости от их группы крови. Поэтому люди с группой крови 0(I) являются «универсальными донорами». При группах крови A(II) и B(III), каждая из которых встречается примерно у трети населения, в сыворотке крови присутствуют антитела соответственно либо против антигена В, либо против антигена А. Поэтому людям с этими группами крови можно переливать либо кровь той же самой группы, либо кровь группы 0(I). При четвертой группе крови AB(IV) антитела против эритроцитарных антигенов в сыворотке крови не вырабатываются. Этим людям можно переливать кровь любой группы, таким образом, они являются «универсальными реципиентами». Однако их кровь можно переливать людям только с той же самой четвертой группой крови AB(IV). Группы крови системы MN. Первый случай кодоминантного взаимодействия аллелей у человека был описан для групп крови системы MN. В этой системе существует три группы M, N и MN. В ходе обширного исследования было показано, что у родителей с одинаковой группой крови M или N рождаются дети, с таким же фенотипом, как и у родителей. Это значит, что обладатели группы крови M или N могут быть только гомозиготами MM или NN соответственно. Дети с группой MN появляются тогда, когда один из родителей имеет группу крови M, а другой N. В этом случае оба аллеля функционируют вместе, и это проявляется в формировании особого фенотипа MN. Группы крови системы Rh. Эта система включает три пары антигенов (D, C/c, E/e), кодируемые двумя тесно сцепленными высоко гомологичными генами, локализованными в коротком плече хромосомы 1. Основная роль в Rh-системе принадлежит антигену D. При его наличии на поверхности эритроцитов кровь является резус-положительной. Антигены C/c и E/e они образуются в результате альтернативного сплайсинга. Резус-отрицательный фенотип формируется при отсутствии антигена D. Знание групповой принадлежности по Rh-системе имеет огромное значение для предотвращения резус-конфликта между матерью и плодом, который может возникнуть во время беременности. Частота людей с резус-положительной принадлежностью – Rh(+), составляет 85%, остальные 15% являются резус-отрицательными – Rh(-). Если у резус-отрицательной женщины муж имеет резус-положительную принадлежность, то с высокой вероятностью ребенок окажется резус-положительный, и тогда может возникнуть резус-конфликт между плодом и матерью. В 15% подобных случаев после 7 недели, когда в крови плода появляются зрелые эритроциты, в крови беременных с Rh(-) могут начать вырабатываться специфические противорезусные антитела. Через плаценту они попадают в кровь плода и в отдельных случаях могут там накапливаться в большом количестве, вызывая агглютинацию эритроцитов и их разрушение. Как правило, первая беременность заканчивается благополучно, мертворождения и выкидыши встречаются редко. Особенно велика вероятность возникновения резус-конфликта при повторных беременностях Rh(-)-женщины. Следствием этого процесса может быть разрушение красных кровяных телец плода и формирование у него гемолитической болезни, проявляющейся анемией, желтухой, отеками и обусловливающей сложные интеллектуальные дефекты, нарушения слуха и речи, двигательные расстройства. Нередко у новорожденных с гемолитической болезнью, вызванной резус-конфликтом, развивается тяжелый детский церебральный паралич с эпилептической болезнью и значительным отставанием психического развития. Для профилактики резус-конфликта и гемолитической болезни у плода женщине с отрицательной резус-принадлежностью при любом внутриматочном вмешательстве во время первой беременности (медицинский аборт, самопроизвольный выкидыш с последующим выскабливанием, роды) показано введение анти-Д-иммуноглобулина. Этот препарат снижает резус-сенсибилизацию беременной, то есть её чувствительность к резус-фактору и соответственно формированию резусных антител. Введение анти-Д-иммуноглобулина при повторных беременностях не показано, так как женщина уже сенсибилизирована, то есть чувствительна к резус-фактору, и имеет резусные антитела. Женщина с Rh(-) непременно должна обсудить с врачом-генетиком проблемы профилактики рождения ребенка с последствиями билирубиновой энцефалопатии в виде тяжелого детского церебрального паралича. В редких случаях конфликт возникает и по АВ0 системе, но протекает он в значительно более легкой форме, чем при резус-конфликте. 35. Геном. Генотип. Геномные мутации и их классификация. Возможные механизмы возникновения и последствия геномных мутаций. Примеры у человека. Генотип как сбалансированная система. Геномом называют всю совокупность наследственного материала, заключенного в гаплоидном наборе хромосом клеток данного вида организмов. Геном видоспецифичен, так как представляет собой тот необходимый набор генов, который обеспечивает формирование видовых характеристик организмов в ходе их нормального онтогенеза. Например, у некоторых видов появляются гаплоидные организмы, которые развиваются на основе одинарного набора генов, заключенного в геноме. Генотип – это объединение геномов двух родительских особей в процессе оплодотворения при половом размножении. Все соматические клетки такого организма обладают двойным набором генов, полученных от обоих родителей в виде определенных аллелей. Таким образом, генотип — это генетическая конституция организма, представляющая собой совокупность всех наследственных задатков его клеток, заключенных в их хромосомном наборе — кариотипе. |