Коллоквиум 1 по биологии. 1 Биология как наука. Методы научного познания. Биология это прежде всего, основа медицины. Медицина, взятая в плане теории это прежде всего общая биология
 Скачать 281.85 Kb. Скачать 281.85 Kb.
|

|
23. Что такое пластиды, какова их роль в клетке? Классификация пластид. Пластиды являются основными цитоплазматическими органеллами клеток автотрофных растений. Название происходит от греческого слова «plastos», что в переводе означает «вылепленный». Главная функция пластид – синтез органических веществ, благодаря наличию собственных ДНК и РНК и структур белкового синтеза. В пластидах также содержатся пигменты, обусловливающие их цвет. Все виды данных органелл имеют сложное внутреннее строение. Снаружи пластиду покрывают две элементарные мембраны, имеется система внутренних мембран, погруженных в строму или матрикс. Классификация пластид по окраске и выполняемой функции подразумевает деление этих органоидов на три типа: хлоропласты, лейкопласты и хромопласты. Пластиды водорослей именуются хроматофорами. Хлоропласты – это зеленые пластиды высших растений, содержащие хлорофилл – фотосинтезирующий пигмент. Представляют собой тельца округлой формы размерами от 4 до 10 мкм. Химический состав хлоропласта: примерно 50% белка, 35% жиров, 7% пигментов, малое количество ДНК и РНК. У представителей разных групп растений комплекс пигментов, определяющих окраску и принимающих участие в фотосинтезе, отличается. Это подтипы хлорофилла и каротиноиды (ксантофилл и каротин). При рассматривании под световым микроскопом видна зернистая структура пластид – это граны. Под электронным микроскопом наблюдаются небольшие прозрачные уплощенные мешочки (цистерны, или граны), образованные белково-липидной мембраной и располагающиеся в непосредственно в строме. Причем некоторые из них сгруппированы в пачки, похожие на столбики монет (тилакоиды гран), другие, более крупные находятся между тилакоидами. Благодаря такому строению, увеличивается активная синтезирующая поверхность липидно-белково-пигментного комплекса гран, в котором на свету происходит фотосинтез. Хромопласты – пластиды, окраска которых бывает желтого, оранжевого или красного цвета, что обусловлено накоплением в них каротиноидов. Благодаря наличию хромопластов, характерную окраску имеют осенние листья, лепестки цветов, созревшие плоды (помидоры, яблоки). Данные органоиды могут быть различной формы – округлой, многоугольной, иногда игольчатой. Лейкопласты представляют собой бесцветные пластиды, основная функция которых обычно запасающая. Размеры этих органелл относительно небольшие. Они округлой либо слегка продолговатой формы, характерны для всех живых клеток растений. В лейкопластах осуществляется синтез из простых соединений более сложных – крахмала, жиров, белков, которые сохраняются про запас в клубнях, корнях, семенах, плодах. Под электронным микроскопом заметно, что каждый лейкопласт покрыт двухслойной мембраной, в строме есть только один или небольшое число выростов мембраны, основное пространство заполнено органическими веществами. В зависимости от того, какие вещества накапливаются в строме, лейкопласты делят на амилопласты, протеинопласты и элеопласты. Все виды пластид имеют общее происхождение и способны переходить из одного вида в другой. Так, превращение лейкопластов в хлоропласты наблюдается при позеленении картофельных клубней на свету, а в осенний период в хлоропластах зеленых листьев разрушается хлорофилл, и они трансформируются в хромопласты, что проявляется пожелтением листьев. В каждой определенной клетке растения может быть только один вид пластид. 24. Что такое включение, какова их роль в клетке? Классификация включений. Классификация включений Включения - непостоянные структурные компоненты цитоплазмы. Классификация включений: трофические: лецитин в яйцеклетках; гликоген; липиды, имеются почти во всех клетках; секреторные: секреторные гранулы в секретирующих клетках (зимогенные гранулы в ацинозных клетках поджелудочной железы); секреторные гранулы в эндокринных железах и другие; экскреторные: вещества, подлежащие удалению из организма (например, гранулы мочевой кислоты в эпителии почечных канальцев); пигментные: меланин; гемоглобин; липофусцин; билирубин и другие. В процессе жизнедеятельности в некоторых клетках накапливаются случайные включения: медикаментозные; частички угля; кремния и так далее. Эти включения имеют определенный цвет и придают окраску всей клетке (меланин - черный или коричневый, гемоглобин - желто-красный и так далее). Необходимо отметить, что пигментные включения характерны только для определенных типов клеток (меланин содержится в меланоцитах, гемоглобин - в эритроцитах). Однако, липофусцин может накапливаться во многих типах клеток обычно при их старении. Его наличие в клетках свидетельствует о их старении и функциональной неполноценности. 25. Происхождение Эукариотической клетки. Эндосимбиотическая теория происхождения ряда органоидов клетки. Наиболее популярна в настоящее время симбиотическая гипотеза происхождения эукариотических клеток, согласно которой основой, или клеткой-хозяином, в эволюции клетки эукариотического типа послужил анаэробный прокариот, способный лишь к амебоидному движению. Переход к аэробному дыханию связан с наличием в клетке митохондрии, которые произошли путем изменений симбионтов — аэробных бактерий, проникших в клетку-хозяина и сосуществовавших с ней. РЕШАЮЩИЕ ДОКАЗАТЕЛЬСТВА ЭНДОСИМБИОТИЧЕСКОГО ПРОИСХОЖДЕНИЯ ХЛОРОПЛАСТОВ И МИТОХОНДРИЙ Наиболее существенные доказательства происхождения важнейших органелл эукариотических клеток - хлоропластов и митохондрий из прокариот получены в последние годы при изучении структуры одного из наиболее консервативных (в отношении возможности изменений в процессе эволюции) компонентов живых клеток - рибосомальных РНК. Выдающийся американский молекулярный биолог К. Воз с сотрудниками [1] показал уже в 80-х годах, что структура так называемой 16S РНК из рибосом хлоропластов растений совершенно непохожа на соответствующую ей 18S РНК из цитоплазматических рибосом растений и почти идентична структуре аналогичной РНК из некоторых цианобактерий. В то же время эта группа исследователей установила, что 16S РНК рибосом, полученных из митохондрий разных эукариотических организмов, непохожа по своей структуре на 16S РНК рибосом из цитозоля тех же самых биологических объектов, но чрезвычайно схожа с 16S РНК некоторых бактерий, в частности бактерий рода Paracoccus. Интересно, что и набор дыхательных ферментов указанной бактерии очень похож на "дыхательный ансамбль" митохондрий животных. Все эти данные достаточно ясно свидетельствуют в пользу большой вероятности эндосимбиотического происхождения эукариотических клеток. Уместно также сказать, что и по сей день явление эндосимбиоза чрезвычайно широко распространено в природе. Так, например, анаэробная амеба Pelomyxa polustris, которая в отличие от других эукариот не имеет митохондрий, содержит, однако, когда-то фагоцитированные ею аэробные бактерии-симбионты, выполняющие функцию митохондрий. Кроме того, известно, что на теле некоторых морских рыб и других животных часто поселяются светящиеся бактерии, которые не только сами получают большие преимущества для своей жизнедеятельности (в первую очередь из-за того, что им не нужно постоянно искать пищу), но и очень полезны для организмов-хозяев. Они, по-видимому, облегчают им жизнь и помогают ориентироваться среди "своих" и "чужих". Существует и множество других примеров взаимовыгодного эндосимбиоза прокариот с различными эукариотическими организмами. 26. Строение и функций хромосом. Хромосомы состоят в основном из ДНК и белков, которые образуют нуклеопротеиновый комплекс. Белки составляют значительную часть вещества хромосом. На их долю приходится около 65 % массы этих структур. Все хромосомные белки разделяются на две группы: гистоны и негистоновые белки. РНК хромосом представлена в основном продуктами транскрипции, еще не покинувшим место синтеза. Регуляторная роль компонентов хромосом заключается в «запрещении» или «разрешении» считывания информации с молекулы ДНК. В первой половине митоза хромосомы состоят из двух хроматид. соединенных между собой в области первичной перетяжки (центромеры) особым образом организованного участка хромосомы, общего для обеих сестринских хроматид. Во второй половине митоза происходит отделение хроматид друг от друга. Из них образуются однонитчатые дочерние хромосомы,распределяющиеся между дочерними клетками. Кариотип – диплоидный набор хромосом, свойственный соматическим клеткам организмов данного вида, являющийся видоспецефическим признаком и характеризующийся определённым числом и строением хромосом. Если число хромосом в гаплоидном наборе половых клеток обозначить п, то общая формула кариотипа будет выглядеть как 2п, где число п различно для разных видов. Хромосомы — структуры клетки, хранящие и передающие наследственную информацию. Хромосома состоит из ДНК и белка. Комплекс белков, связанных с ДНК, образует хроматин. Белки играют важную роль в упаковке молекул ДНК в ядре. ДНК в хромосомах упакована таким образом, что умещается в ядре, диаметр которого обычно не превышает 5 мкм (5-10- 4 см). Упаковка ДНК приобретает вид петельной структуры, похожей на хромосомы типаламповых щеток амфибий или политенных хромосом насекомых. Петли поддерживаются с помощью белков, которые узнают определенные последовательности нуклеотидов и сближают их. Строение хромосомы лучше всего видно в метафазе митоза. Хромосома представляет собой палочковидную структуру и состоит из двух сестринских хроматид, которые удерживаются центромерой в области первичной перетяжки. Каждая хроматид а построена из хроматиновых петель. Хроматин не реплицируется. Реплицируется только ДНК. С началом репликации ДНК синтез РНК прекращается. Хромосомы могут находиться в двух состояниях: конденсированном (неактивном) и деконденсированном (активном). Диплоидный набор хромосом организма называют ка-риотипом. Современные методы исследования позволяют определить каждую хромосому в кариотипе. Для этого учитывают распределение видимых под микроскопом светлых и темных полос (чередование AT и ГЦ-пар) в хромосомах, обработанных специальными красителями. Поперечной исчер-ченностью обладают хромосомы представителей разных видов. У родственных видов, например у человека и шимпанзе, очень сходный характер чередования полос в хромосомах. Каждый вид организмов обладает постоянным числом, формой и составом хромосом. В кариотипе человека 46 хромосом — 44 аутосомы и 2 половые хромосомы. Мужчины ге-терогаметны (ХУ), а женщины гомогаметны (XX). У-хромосо-ма отличается от Х-хромосомы отсутствием некоторых аллелей (например, аллеля свертываемости крови). Хромосомы одной пары называют гомологичными. Гомологичные хромосомы в одинаковых локусах несут аллельные гены. 27. Принципы классификаций хромосом. Денверская и Парижская классификация хромосом их сущность. Классификация хромосом человека Хромосомный набор (кариотип) соматической клетки характеризуется формой хромосом, их количеством, размерами, характерными для каждого вида. Препараты хромосом можно приготовить извсех тканей и клеточных суспензий, содержащих делящиеся клетки. Наиболее часто препараты метафазных хромосом готовят из лимфоцитов периферической крови, которые предварительно культивируют в присутствии стимулятора митозов — митогена фитогемаг- глютинина (ФГА), так как клетки периферической крови не делятся. Классификация и номенклатура равномерно окрашенных хромосом (стандартное окрашивание) была разработана на международных совещаниях в Денвере (1960), Лондоне (1963) и Чикаго (1966). Согласно рекомендациям этих конференций, хромосомы располагаются в порядке уменьшения их длины от 1 до 23 хромосомы. Они разделены на 7 групп, которые обозначены буквами английского алфавита от А до G. Все пары хромосом предложено нумеровать арабскими цифрами: группа А 1-3, В 4-5, С 6-12 и Х-хромосома, D 13-15, Е 16-18, F 19- 20, G 21-22, Y-хромосома (табл. 13). При этом хромосомы различных групп хорошо отличаются друг от друга (критерии: размер и форма — метацентрические, субметацентрические, акроцентрические), в то время как внутри группы их сложно различить, за исключением хромосом группы А (рис. 64,65). Важным параметром является центромерный индекс, который отражает в процентах длину короткого плеча к длине всей хромосомы, например, хромосома № 1 — 48,3%, хромосома № 22 — 22,1%. 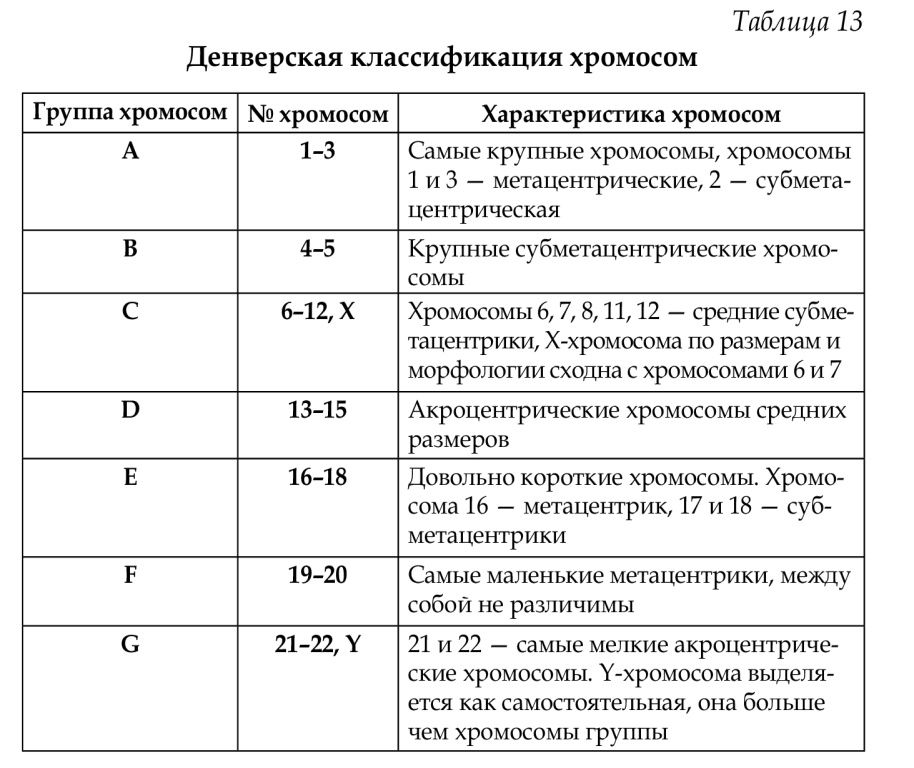 Парижская классификация хромосом В начале 70-х годов XX века был разработан метод дифференциальной окраски хромосом, выявляющий характерную сегментацию, который позволил индивидуализировать каждую хромосому (рис. 58). Различные типы сегментов обозначают по методам, с помощью которых они выявляются наиболее отчетливо (Q-сегменты, G-сегменты, Т-сегменты, S-сегменты). Каждая хромосома человека содержит свойственную только ей последовательность полос, что позволяет идентифицировать каждую хромосому. Хромосомы спирализованы максимально в метафазе, менее спирализованы в профазе и прометафазе, что позволяет выделить большее число сегментов, чем в метафазе. На метафазной хромосоме (рис. 59) приводятся символы, которыми принято обозначать короткое и длинное плечо, а также расположение районов и сегментов. В настоящее время существуют ДНК-маркеры или зонды, с помощью которых можно определить изменение определенного, даже очень маленького, сегмента в хромосомах (цитогенетические карты). На международном конгрессе генетики человека в Париже в 1971 г. (Парижская конференция по стандартизации и номенклатуре хромосом человека) была согласована система символов для более краткого и однозначного обозначения кариотипов. При описании кариотипа: • указывается общее число хромосом и набор половых хромосом, между ними ставится запятая (46, XX; 46, XY); • отмечается какая хромосома лишняя или какой не хватает (это ука-зывается ее номером 5, 6 и др., или буквами данной группы А, В и др.); знаком «+» указывают на увеличение количества хромосом, знаком «-» указывают на отсутствие данной хромосомы 47, XY,+ 21; • плечо хромосомы, в котором произошло изменение (удлинение короткого плеча указывается символом (р+); укорочение (р-); удлинение длинного плеча указывается символом (q+); укорочение (q-); • символы перестроек (транслокация обозначается t, а делеция — del) помещают перед номерами вовлеченных хромосом, а перестроечные хромосомы заключают в скобки. Наличие двух структурно-аномальных хромосом обозначается точкой с запятой (;) или нормальной дробью (15/21). Система записи кариотипов 46, XX — нормальный кариотип (женщина) 46, XY — нормальный кариотип (мужчина) 45, X — синдром Шерешевского-Тернера 47 XXY 1 4g' xxxY I — синдром Клайнфельтера 47, XXX — синдром «трисомии по Х-хромосоме» 47, XYY — синдром Вай-Вай 47, XX, + 21 — синдром Дауна (женщина) 47, XY, + 21 — синдром Дауна (мужчина) 47, XX, + 18 — синдром Эдвардса (женщина) 47, XY, + 18 — синдром Эдвардса (мужчина) 47, XX, + 13 — синдром Патау (женщина) 47, XY, + 13 — синдром Патау (мужчина) 46, XX, t (9/22) — хронический миелолейкоз (женщина) 46, XY, t (9/22) — хронический миелолейкоз (мужчина) 46, XX, t (15/21) — транслокационный Даун (женщина) 46, XY, t (15/21) — транслокационный Даун (мужчина) 46, XX, del (5p-) — синдром кошачьего крика (женщина) 46, XY, del (5р-) — синдром кошачьего крика (мужчина) 46, XX, del (13q-) — синдром Орбели (женщина) 46, XY, del (13q-) — синдром Орбели (мужчина) Сегменты и районы метафазной хромосомы обозначаются цифрами, центромера служит исходной точкой для цифровой схемы. При определении локализации гена используют 4 критерия: номер хромосомы, символ плеча, номер района и номер сегмента в пределах этого района. Например, запись 1р32 означает, что речь идет о хромосоме первой пары, коротком плече, районе 3, сегменте 2. Для гена Rh (рис. 59) формула локализации: 1р35. 28.цитологических методов исследования. Световая и электронная микроскопия Постоянные и временные препараты биологических объектов А. Цитологическое исследование (греч. kytos вместилище, здесь – клетка + logos учение) – исследование, основанное на изучении с помощью микроскопа особенностей строения клеток, клеточного состава органов, тканей, жидкостей организма человека и животных в норме и при патологических процессах. Цитологическое исследование широко применяется в биологии для изучения закономерностей строения и жизнедеятельности клетки и в медицине – для диагностики различных заболеваний. Методы цитологического исследования, используемые в судебной медицине, позволяют обнаруживать клеточные элементы повреждённых тканей на различных орудиях, транспортных средствах и др. Когда применяется цитологическое исследование? Цитологический анализ используют при: Скрининге (профилактическом осмотре). Установлении (уточнении) диагноза при заболевании. Установлении (уточнении) диагноза во время операции. Контроле в ходе лечения и после лечения. Динамическом наблюдении (для раннего выявления рецидивов). Материалы для цитологического исследования Жидкости: мокрота, моча, сок предстательной железы, смывы из различных органов во время эндоскопии, а также из шейки и полости матки (цитологическое исследование мазков, цитологическое исследование шейки матки), выделения из молочных желез, соскобы и отпечатки с эрозированных или язвенных поверхностей, свищей, ран, жидкость из суставных и серозных полостей, цереброспинальная и амниотическая жидкость; Пунктаты: материалы, полученного при аспирационной диагностической пункции, преимущественно тонкой иглой; Отпечатки с удаленных тканей, например поверхности свежего разреза оперативно удаленной или взятой для гистологического исследования ткани. Б. Световая и электронная микроскопия Микроскоп - устройство, которое позволяет видеть увеличенное изображение объектов и структур, которые не видны глазу человека. В медико-биологических исследованиях используются световые и электронные методы микроскопии. Микроскопы, основанные на световой технологии, позволяют увеличивать объекты от 0.5 микрометров с разрешением объектов до 0.1 микрометра больше чем в 1500 раз. Микроскопы, основанные на электронной технологии - до 20 000 раз. Технология световой микроскопии базируется на фундаментальных законах оптики, а так же на волновой теории в образовании изображений. Для освещения используют естественный, либо искусственный источники света. Первые простые микроскопы появились еще в 17-м веке. Более высоких успехов в их разработке смог добиться ученый из Голландии, А. Левенгук. В период с 1609 по 1610 гг. Г. Галилеем был построен более сложный микроскоп. В 1846 г. немецкий инженер-механик К. Цейсе открыл свою мастерскую и, примерно через год, начал изготавливать микроскопы. Цейс в своей фирме успешно использовал научные открытия профессора по физике Эрнста Аббе, который позже становится полноправным компаньоном Цейса. Теоретические и практические работы Э. Аббе, О. Шотта и А. Келера определили направления в развитии и принципы строения оптических систем в современных микроскопах. Электронная микроскопия Технология электронной микроскопии позволяет получать электронно-оптическое изображение при помощи потока электронов. Построение изображений базируется на фундаментальных законах волновой и геометрической оптики, а так же теории электромагнитного поля. Технология электронной микроскопии дает возможности для исследования объектов, у которых размеры лежат за пределами разрешающих возможностей светового микроскопа, а именно – объекты, менее 0.2 микрометров, и находит свое применение в изучении вирусов, бактериофагов, тонкого клеточного строения и других микрообъектов. Также такие микроскопы с успехом применяются для изучения макромолекулярных структур. Очень важно не путать электронный микроскоп и цифровой микроскоп. В последнее время многие ошибочно называют цифровые usb микроскопы – электронными микроскопами. На самом же деле, это не так. Микроскоп с электронной технологией появился в конце 30-х годов. Серийным запуском электронных микроскопов занялась фирма немецкого происхождения "SIEMENS". В 1940 году в ГОИ имени С. И. Вавилова, находящимся в Ленинграде, создали первый отечественный микроскоп с электронной технологией, увеличительные способности которого достигали 10000 крат. Микроскопы с электронной технологией используются, в основном, в научно-исследовательских лабораторных комплексах, для серьезных научных разработок. Минимальная стоимость таких изделий начинается примерно с двух-трех миллионов рублей. В. Биологические препараты — группа медицинских продуктов биологического происхождения, в том числе вакцины, препараты крови, аллергены, соматические клетки, ткани, рекомбинантные белки. В состав биологических препаратов могут входить сахара, белки, нуклеиновые кислоты или сложные комбинации этих веществ; биологические препараты могут представлять собой биологические объекты — например, клетки и ткани. Биологические препараты получают из различных природных источников — животных, микроорганизмов, также биологические препараты могут быть синтезированы методами биотехнологии. Активно исследуется потенциал медицинского применения клеточных и генных биологических препаратов для лечения многих заболеваний, неизлечимых в настоящий момент. |