1. Экология как наука. Положение экологии в системе наук о природе
 Скачать 1.32 Mb. Скачать 1.32 Mb.
|

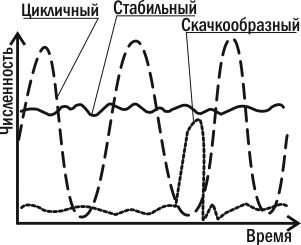 Три типа популяционной динамики. Можно заключить, что численность каждой популяции находится под контролем множества отрицательных обратных связей. Когда численность популяции начинает расти, ее сдерживают и недостаток собственных ресурсов, и переключение на питание ее особями хищников-полифагов, и избыточное размножение специализированных хищников, и рост паразитарных инфекций. Все эти факторы «возвращают» численность популяции к прежнему уровню и снижают давление на нее, когда эта численность оказывается невысокой. В зависимости от того, насколько существенным окажется отклонение численности популяции от нормы, к ее возвращению подключится большее или меньшее количество регулирующих ее факторов 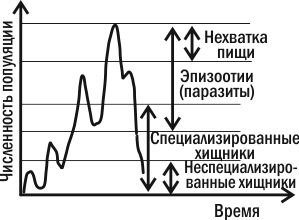 Из всех причин, вызывающих колебания численности, некоторые исследователи главную роль отводят климатическим условиям, считая биотические факторы второстепенными. По мнению других, только те факторы, которые зависят от плотности, вызывают колебания численности. Многие ученые признают роль тех и других. В экосистемах с низким уровнем видового разнообразия численность популяций подвержена сильному воздействию физических стрессоров и зависит от погоды, химического состава среды и степени ее загрязнения. В экосистемах с высоким уровнем видового разнообразия колебания численности популяций в основном контролируются биотическими факторами. Все экологические факторы в зависимости от характера их влияния на численность популяции можно разделить на две группы: 1) факторы, независимые от плотности; 2) факторы, зависимые от плотности. К первой группе относятся факторы, действующие на популяции постоянно и изменяющие их численность однонаправленно, независимо от величины популяции. Так действуют абиотические факторы и прежде всего климатические факторы, а также антропогенные факторы, за исключением природоохранных факторов. Действие их не всегда проявляется сразу. Эти факторы еще называют модифицирующими, поскольку они не обеспечивают регуляции численности, а лишь вызывают ее изменения. Ко второй группе относятся факторы, действие которых на популяцию может быть прямым, т.е. усиливаться по мере приближения к верхнему пределу, или инверсным, т.е. ослабевать по мере увеличения плотности. Эти факторы изменяют численность в сторону оптимального уровня и предотвращают перенаселение, поэтому В.Викторов предложил называть их регулирующими факторами. К ним относятся биотические факторы и природоохранные антропогенные факторы. Зависимые от плотности факторы, как правило, влияют на скорость роста численности популяции путем изменения ее рождаемости или смертности. Биотические факторы, зависимые от плотности, по характеру влияния на численность популяции подразделяются на две группы: 1) внутривидовые; 2) межвидовые. Рассмотрим характер действия внутривидовых факторов. К.Вини-Эдвардс в 1962 году предложил два механизма стабилизации плотности ниже уровня насыщения за счет внутривидовых факторов: 1) территориальное поведение; 2) групповое поведение. При нарушении границ за счет увеличения плотности наблюдаются разные формы внутривидовой конкуренции: 1) конкуренция за ресурсы. При недостатке жизненно необходимых ресурсов часть животных погибает, а у растений наблюдается самоизреживание и изменение вегетативной мощности; 2) прямой антагонизм - биологическая и химическая «война» в природе. Биологическая «война» - это умерщвление конкурентов внутри популяции путем прямого нападения. Химическая «война» - это выделение химических веществ, задерживающих рост и развитие или убивающих молодых особей (растения, водные животные). 41.Регуляция численности на популяционном уровне. Синтетическая теория колебания численности. новное и принципиально новое положение синтетической теории И.Я. Полякова состоит в том, что она позволяет заблаговременно судить о динамике численности и о вероятных факторах, способных на нее воздействовать, по состоянию кормовой базы, физической среды и морфофизиологическим свойствам популяций. Это делает ее приемлемой для решения задач прогноза численности вредителей (Поляков, 1976). В последние годы 20-го столетия среди отечественных и зарубежных экологов были популярны теоретические концепции, названные Г.А. Викторовым (1973) стохастизмом и регуляционизмом, а современный этап исследований популяционной динамики поиском механизмов регуляции численности. Представление о том, что каждая популяция обладает равновесным уровнем плотности и существуют выработавшиеся в ходе эволюции внутрипопуляционные (или внутриэкосистемные) механизмы, направленные на поддержание этой плотности, лежит в основе подхода, который наиболее точно можно охарактеризовать) как регуляционизм. Для формирования данного подхода большое значение имели выполненные в 30—50-х гг. теоретические и экспериментальные работы австралийского энтомолога А. Никольсона (Nicholson, 1933, 1957), подчеркнувшего, что динамика численности любой популяции есть автоматически регулируемый процесс, а действие, факторов, контролирующих популяцию, управляется плотностью самой контролируемой популяции. Выше уже говорилось об опытах Никольсона с падальной мухой, наглядно продемонстрировавших механизм возникновения в популяциях автоколебаний численности. Принципиально другой подход — это стохастизм, уделяющий основное внимание факторам, случайно действующим, или, точнее, случайно распределенным во времени и в пространстве. Сторонники стохастизма обычно отрицают существование «равновесного» уровня, всякое отклонение от которого будто бы автоматически включает процессы, возвращающие популяцию к исходному уровню. С позиции стохастизма «равновесный уровень численности (т. е. тот, при котором R0 = 1) есть просто артефакт усреднения за длительный срок: чем длиннее имеющийся ряд наблюдений за какой-нибудь популяцией, тем больше шансов утверждать, что средняя плотность, полученная за ряд лет, это и есть «равновесная» плотность, активно поддерживаемая специальными механизмами. Спор сторонников регуляционизма и стохастизма достиг своей кульминации в конце 50-х гг., особенно после выхода в свет капитальной сводки «Распространение и обилие животных», принадлежащей перу австралийских исследователей Г. Андреварты и Л. Берча (Andrewartha, Birch, 1954). Проанализировав очень большой эмпирический материал (главным образом по насекомым), Андреварта и Берч пришли к выводу о том, что популяции животных в природе обычно ограничены: 1) нехваткой ресурсов (пищи, подходящих мест для гнездовий и т. д.); 2) недоступностью этих ресурсов вследствие ограниченных возможностей расселения животных; 3) кратковременностью периода, в течение которого скорость роста популяции r сохраняет положительное значение. По мнению сторонников стохастизма, распространение какого-либо вида организмов в пространстве и динамика численности его популяций во времени ограничены одними и теми же факторами, тогда как сторонники регуляционизма полагают, что распространение организмов лимитируется главным образом абиотическими (независимыми от плотности) факторами, а динамика численности — биотическими, как правило, зависимыми от плотности. Соронники первого направления (Thompson, 1956; Bodenheimer, 1958; Andrewartha, Brich, 1954; Milne, 1984 и др.) считают воздействия факторов внешней среды на популяции случайными. Комбинации различных факторов определяют изменения численности насекомых (подъемы и спады), причем благоприятное сочетание условий, определяющих подъемы численности, наблюдается в природе гораздо реже, чем неблагоприятное. Представители второго направления рассматривают колебания численности как регулируемый процесс. Они считают, что случайные изменения ее, вызванные прямым или косвенным воздействием абиотических (главным образом физических) факторов, компенсируются деятельностью регуляторных механизмов, которые управляются изменениями плотности популяций по принципу отрицательной обратной связи. По мнению сторонников регуляционизма, в этой роли могут выступать биотические факторы среды, реагирующие на изменения численности других организмов (Howard, Ficke, 1911; Varley, 1953; Lack, 1956; Winne-Edmards, 1965; Klomp, 1964; Уатт, 1971; Дажо, 1975; Ю. Одум, 1986; Уильямсон, 1975; Варли, Градуэлл, Хассел, 1978; Риклефс, 1979; Пианка, 1981). Согласно представлениям большинства современных экологов, изменение численности насекомых рассматривается как взаимодействие различных механизмов. Г.А. Викторов (1967) подразделил их на модифицирующие и регулирующие. К модифицирующим он отнес климатические и другие географические факторы среды, к регулирующим — естественных врагов (паразиты, хищники, возбудители болезней), внутривидовые отношения (конкуренция), а также трофические факторы (количество, качество и доступность пищи). Синтетическая теория в варианте И.Я. Полякова при всех ее положительных сторонах не решает проблему многолетних флюктуаций численности животных. Характерно, что она используется в практике защиты растений лишь для краткосрочных биологических прогнозов. 42-46.Классификация межвидовых взаимоотношений. Отрицательные и положительные взаимоотношения./Конкуренция и хищничество./Паразитизм и аллелопатия./Комменсализм, протокооперация, мутуализм./Консорции. Возможны следующие виды влияний одних организмов на другие:
Симбиоз: + + Облигатный симбиоз — форма симбиоза, при которой в естественных условиях популяции не могут существовать друг без друга (пример: симбиоз гриба и водоросли в лишайнике).Разновидностью облигатного симбиоза является: Мутуализм — форма облигатного взаимовыгодного сожительства организмов двух и более видов. Примером мутуализма является симбиоз рыб-клоунов с актиниями. Вначале рыба слегка касается актинии, позволяя ей ужалить себя и выясняя точный состав слизи, которым покрыта актиния, — эта слизь нужна актинии, чтобы она сама себя не жалила. Затем рыба-клоун воспроизводит этот состав и после этого может прятаться от врагов среди щупалец актинии. Рыба-клоун заботится об актинии — вентилирует воду и уносит непереваренные остатки пищи. Рыбки никогда не удаляются далеко от «своей» актинии. Самцы прогоняют от неё самцов, самки — самок. Территориальное поведение, видимо, стало причиной контрастной окраски. Самая тесная форма мутуализма — когда один организм живёт внутри другого. Поразительным примером этого служит система органов пищеварения коров и других жвачных животных. Коровы, как и человек, не способны переварить целлюлозу — вещество, которое в большом количестве содержится в растениях. Но у жвачных животных есть особый орган — рубец. Он представляет собой полость, в которой живут множество микробов. Растительная пища, после того как животное её прожевало, попадает в рубец, и там эти микробы разрушают целлюлозу. (Животное может отрыгнуть и вновь прожевать частично расщеплённую пищу — именно этим и занимаются коровы, когда пережёвывают свою жвачку.) Рубец коровы — это замкнутая микроэкосистема, образованная множеством различных микроорганизмов, задача которых состоит в переработке целлюлозы для своего хозяина. Аналогично корневая система высших растений образована переплетением корневой ткани и грибных нитей, так что грибы снабжают растение минеральными веществами. Мутуализм может быть «жёстким» или «мягким». В первом случае сотрудничество жизненно необходимо для обоих партнёров (они связаны отношениями коадаптации), во втором отношения более или менее факультативны (это называется протокооперацией). + + Факультативный симбиоз (Протокооперация (англ.)) — форма симбиоза, при которой совместное существование выгодно, но не обязательно для сожителей. (например, взаимоотношения краба и актинии: актиния защищает краба и использует его в качестве средства передвижения) 0 + Комменсализм — форма симбиоза, при которой одна популяция извлекает пользу от взаимоотношения, а другая не получает ни пользы, ни вреда. Выделяют следующие типы комменсализма: зоохорию, паройкию, синойкию (квартирантство), энтойкию, эпибиоз, эпиойкию (эпойкию, нахлебничество) , форезию. Комменсализм (от лат. com — «с», «вместе» и mensa — «стол», «трапеза»; буквально «у стола», «за одним столом»; ранее — сотрапезничество) — способ совместного существования (симбиоза) двух разных видов живых организмов, при котором один из партнёров этой системы (комменсал) возлагает на другого (хозяина) регуляцию своих отношений с внешней средой, но не вступает с ним в тесные взаимоотношения[1]. При этом, популяция комменсалов извлекает пользу от взаимоотношения, а популяция хозяев не получает ни пользы, ни вреда (например, чешуйница обыкновенная и человек), т.е. метаболические взаимодействия и антагонизм между такими партнёрами чаще всего отсутствуют. Комменсализм — как бы переходная форма от нейтрализма к мутуализму. Классификация:
− + Паразитизм — форма симбиоза, при которой один организм (паразит) использует другой (хозяин) в качестве источника питания или/и среды обитания, возлагая при этом (частично или полностью) на хозяина регуляцию своих отношений с внешней средой. Паразитизм так же бывает облигатным, когда паразит не может существовать без хозяина (типичный пример — вирусы) и факультативным (вши, блохи, паразитические черви и т. д.).
Хищничество: − + Хищничество — явление, при котором один организм питается органами и тканями другого (при этом умерщвление жертвы не обязательно), при этом не наблюдается симбиотических отношений. Но в современной экологии часто используется общее понятие хищничества, в которое также входят паразитизм и растительноядность (фитофагия). Нейтрализм: 0 0 Нейтрализм — обе популяции не оказывают никакого воздействия друг на друга. Антибиоз: − 0 Аменсализм— форма антибиоза, при которой одна популяция отрицательно влияет на другую, но сама не испытывает ни отрицательного, ни положительного влияния. Типичный пример — высокие кроны деревьев, угнетающие рост низкорослых растений и мхов, за счет частичного перекрывания доступа солнечного света. − − Аллелопатия — форма антибиоза, при которой организмы оказывают взаимно вредное влияние друг на друга, обусловленное их жизненными факторами (например, выделениями веществ). Встречается, в основном, у микроорганизмов, растений, грибов. При этом вредное влияние одного организма на другой не является необходимым для его жизнедеятельности и не приносит ему пользы. − − Конкуренция — форма антибиоза, при которой два вида организмов являются биологическими врагами по своей сути (как правило, из-за общей кормовой базы или ограниченных возможностей для размножения). Например, между хищниками одного вида и одной популяции или разных видов, питающихся одной пищей и обитающих на одной территории. В этом случае вред, причиняемый одному организму приносит пользу другому, и наоборот. Консо́рция (от англ. consortium — соучастие, сообщество) — ценотическая популяция свободноживущего зелёного растения вместе со всеми связанными с ним популяциями других организмов (животные, бактерии, грибы и т. д.). Представление о консорциуме, как о структурной единице биогеоценоза, сформулировано в начале 50-х годов XX столетия зоологом В. Н. Беклемишевым (1890—1962) и геоботаником Л. Г. Раменским (1884—1953). Популяцию зелёного растения называют ядром или детерминантом консорции, остальные организмы — консортами. Так, в лесном биогеоценозе ядром консорции может быть ель, консортами — различные насекомые, питающиеся за счёт ели, грибы, паразитирующие на ели, птицы, гнездящиеся на ней и т. д. Взаимосвязи между организмами в пределах консорции чрезвычайно разнообразны. В одних случаях консорты связаны с автотрофным растением непосредственно (прямо), в других — опосредованно (косвенно) через других консортов; в одних случаях детерминант консорции является для консорта источником вещества и энергии (наблюдаются так называемые трофические связи), в другом случае он используется как местообитание (топические связи), в ряде случаев имеют место смешанные связи. Число видов организмов, образующих консорцию, очень велико. В качестве консортов выступают позвоночные и беспозвоночные животные, грибы, бактерии, актиномицеты, простейшие и т. д. 47-48.Фундаментальная и реализованная экологические ниши./Перекрывание экологических ниш. Хатчинсон (Hutchinson G. Е.) ввел понятие фундаментальной и реализованной экологических ниш. Фундаментальная экологическая ниша — это вся совокупность необходимых для вида условий существования при отсутствии какого-либо давления со стороны другого вида. Реализована ниша — более узкая, она включает конкурентов, хищников. Таким образом, экологическая ниша — это физическое пространство с присущими экологичными условиями, определяющими существование любого организма, место вида в природе (положение в пространстве, функциональная роль в биоценозе, отношение к абиотическим факторам среды). Экологическая ниша характеризует степень биологической специализации вида (Кучерявый, 2000). По Хатчинсоном, экологические ниши могут перекрываться, то есть две фундаментальные ниши идентичны. Ситуация маловероятна. Один вид, более конкурентный, вытисняет другой. Фундаментальная ниша одного вида может быть включена полностью в фундаментальную нишу второго. Экологические ниши частично могут перекрываться. Участок перекрывается, занимает более конкурентоспособный вид. Экологические ниши могут прилегать друг к другу. В таком случае конкурентное исключение невероятное, или является следствием конкурентного исключения. Бывает ситуация, когда нише находятся на расстоянии друг от друга, т.е. каждый вид занимает свою фундаментальную нишу. Ю. Одум в 1959 году писал о экологическую нишу как о статусе организма в группировке и экосистеме, которая вытекает из его структурных адаптаций, физиологических реакций, специфического поведения. В своей известной монографии «Экология» (Одум, 1989) он писал, что среда обитания — это адрес организма, а ниша — это профессия. Где, как, когда, чем питается вид, кто его ест, где он отдыхает и как размножается. 49.Пространственная структура экосистемы. опуляции разных видов в экосистеме распределены определенным образом - образуют пространственную структуру. Различают вертикальную и горизонтальную структуры экосистемы. Основу вертикальной структуры формирует растительность. Растительное сообщество определяет, как правило, облик экосистемы. Растения в значительной мере влияют на условия существования остальных видов. В лесу это крупные деревья, на лугах и в степях - многолетние травы, а в тундрах господствуют мхи и кустарнички. Обитая совместно, растения одинаковой высоты создают своего рода этажи - ярусы. В лесу, например, высокие деревья составляют первый (верхний) ярус, второй ярус формируется из молодых особей деревьев верхнего яруса и из взрослых деревьев, меньших по высоте. Третий ярус состоит из кустарников, четвертый - из высоких трав. Самый нижний ярус, куда попадает совсем мало света, составляют мхи и низкорослые травы. Ярусность наблюдается также в травянистых сообществах (лугах, степях, саваннах). Имеется и подземная ярусность, что связано с разной глубиной проникновения в почву корневых систем растений: у одних корни уходят глубоко в почву, достигают уровня грунтовых вод, другие имеют поверхностную корневую систему, улавливающую воду и элементы питания из верхнего почвенного слоя. Благодаря ярусному расположению растения наиболее эффективно используют световой поток, при этом снижается конкуренция: светолюбивые растения занимают верхний ярус, а теневыносливые развиваются под их пологом. Животные тоже приспособлены к жизни в том или ином растительном ярусе (некоторые вообще не покидают свой ярус). Например, среди насекомых выделяют: подземных, обитающих в почве (медведка, норный паук); наземных, поверхностных (муравей, щитник); обитателей травостоя (кузнечик, тля, божья коровка) и обитателей более высоких ярусов (различные мухи, стрекозы, бабочки). Вследствие неоднородности рельефа, свойств почвы, различных биологических особенностей растения и в горизонтальном направлении располагаются микрогруппами, различными по видовому составу. Это явление носит название мозаичности. Мозаичность растительности - это своего рода "орнамент", образованный скоплениями растений разных видов. Благодаря вертикальной и горизонтальной структурам обитающие в экосистеме организмы более эффективно используют минеральные вещества почвы, влагу, световой поток. 50. Функциональная структуры экосистемы: цепи, сети, уровни. Все организмы, входящее в биоценоз по способу питания, подразделяют на автотрофов и гетеротрофов. Автотрофы (от греч. autos – сам) – осуществляют превращение неорганических веществ в органические (зеленые растения и некоторые микроорганизмы). По механизму превращения неорганических веществ в органические автотрофы делится на: а) фототрофы (фотосинтез) – зеленые растения, сине-зеленые водоросли; б) хемотрофы (хемосинтез) – серные бактерии и др. Гетеротрофы (от греч. разный) – используют для питания готовые органические вещества (все животные и человек, паразиты, грибы и др). По современным данным Дж. Н. Андерсона, гетеротрофов делят на: а) некротрофы (от греч. nekros – мертвый) трупноядные животные; б) биотрофы (от греч. biosis – живой) питаются за счет других живых организмов (паразиты, кровососы и др); в) сапротрофы (от греч. sapros – гниль) питаются отмершей органикой. Существуют организмы и со смешанным типом питания, которых наз. миксотрофами (П.Пфеффер. от англ. mix – смешивать). Поддержание жизнедеятельности организмов и круговорот веществ в экосистемах возможно только за счет постоянного притока энергии. В конечном итоге вся жизнь на земле существует за счет энергии солнечного излучения, которая переводится фотосинтезирующими организмами (автотрофами) в химические связи органических соединений. Все остальные организмы получают энергию с пищей. Все живые существа являются объектами питания других, т.е. связаны между собой энергетическими отношениями. Пищевые связи в сообществах – это механизмы передачи энергии от одного организма к другому. Перенос энергии пищи от ее источника – автотрофов (растений) – через ряд организмов, происходящий путем поедания одних организмов другими, называется пищевой (трофической) цепью. Для высвобождения запасенной химической энергии гетеротрофы разлагают органические соединения на исходные неорганические компоненты, завершая тем самым круговорот веществ. По отношению к трофическим (пищевым) связям организмы экосистемы подразделяются на продуцентов, консументов и редуцентов. Продуценты (производители первичной продукции) - организмы, способные из неорганических веществ создавать органические, т.е. производить и накапливать потенциальную энергию в форме химической энергии, которая содержится в синтезированных органических веществах (углеводах, жирах, белках). В наземных экосистемах такой синтез осуществляют, главным образом, цветковые растения; в водной среде – микроскопические планктонные водоросли. Консументы (т.е потребители) – это организмы, потребляющие органическое вещество продуцентов или других консументов и трансформирующие его в новые формы. Роль консументов выполняют в природе, в основном, животные. Можно выделить консументы различного порядка. Первичные консументы питаются автотрофными (фотосинтезирующими) продуцентами. Это, в основном, травоядные животные. Вторичные консументы питаются травоядными организмами, т.е. являются плотоядными формами. Третичными являются консументы, питающиеся вторичными консументами и т.д. Можно выделить также консументов 4-го и 5-го порядка. Редуценты живут за счет мертвого органического вещества, переводя его вновь в неорганическое соединение. Это, главным образом, бактерии и грибы. Они являются как бы завершающим звеном биологического круговорота веществ. Место каждого звена в цепи питания называют трофическим уровнем или цепью питания. Первый трофический уровень – это всегда продуценты, создатели органической массы; второй – растительноядные консументы; третий – плотоядные, четвертый – организмы, потребляющие других плотоядных. По мере продвижения по цепи хищников животные все более увеличиваются в размерах и уменьшаются численно. Понятие пищевой цепи удобно для изложения, хотя и носит несколько упрощенный характер. Линейные пищевые цепи - большая редкость в природе. Как правило, пищевые цепи в экосистеме тесно переплетаются. Совокупность пищевых связей в экосистеме образует пищевые сети, в которых многие консументы служат пищей нескольким членам экосистемы. В то же время некоторые животные могут принадлежать сразу к нескольким трофическим уровням, так как питаются и растительной, и животной пищей, то есть являются в Интересный пример пищевых сетей. Правило экологической пирамиды. Пищевые сети, возникающие в экосистеме, имеют структуру, для которой характерно определенное число организмов на каждом трофическом уровне. Замечено, что число организмов прямо пропорционально уменьшается при переходе с одного трофического уровня на другой. Такая закономерность получила название "правило экологической пирамиды". 51. Превращение энергии при переходе из одного уровня на другой. Переход вещества и энергии с одного трофического уровня на другой связан с потерями. Считается, что на каждом последующем уровне усваивается лишь 10% вещества и энергии предыдущего уровня. Эта закономерность получила название правила 10-ти % (правило Линдемана). С каждым переходом из одного трофического уровня в другой в пределах пищевой цепи или сети совершается работа и в окружающую среду выделяется тепловая энергия, а количество энергии высокого качества, используемой организмами следующего трофического уровня, снижается. Правило 10%: при переходе с одного трофического уровня на другой 90% энергии теряется, и 10% передается на следующий уровень. Чем длиннее пищевая цепь, тем больше теряется полезной энергии. Поэтому длина пищевой цепи обычно не превышает 4 - 5 звеньев. Пирамиды численностей и биомасс. Мы можем собрать все образцы организмов в экосистеме и подсчитать численность всех видов, обнаруженных на каждом трофическом уровне. Такая информация необходима для создания пирамиды численностей для экосистем. Сухой вес всех органических веществ, содержащихся в организмах экосистемы, называется биомассой. Каждый трофический уровень пищевой цепи или сети содержит определенное количество биомассы. Ее можно вычислить, если собрать все живые организмы с различных произвольно выбранных участков. Собранные экземпляры необходимо рассортировать по трофическим уровням, высушить и взвесить. Полученные данные в дальнейшем используются для построения пирамиды биомасс для определенной экосистемы. Чистая первичная продуктивность растений. Скорость, с которой растения экосистемы производят полезную химическую энергию или биомассу, называется чистой первичной продуктивностью. Чистая первичная продуктивность = скорость, с которой растения производят химическую энергию в процессе фотосинтеза - скорость, с которой растения расходуют химическую энергию в процессе дыхания Первый закон (начало) термодинамики или закон сохранения энергии утверждает, что энергия может переходить из одной формы в другую, но она не исчезает и не создается заново. Закон подразумевает, что в результате превращения энергии никогда нельзя получить ее больше, чем затрачено. Выход энергии всегда равен ее затратам. Закон справедлив как для обычных систем состоящих большого числа частиц, так и для систем из небольшого числа частиц. Первый закон термодинамики: Существует два способа изменения внутренней энергии - теплопередача и работа. дельта U = A+Q дельта U - изменение внутренней энергии тела; Q - полученное им количество теплоты; A - совершенная над телом работа. Эта формула гласит, что изменение внутренней энергии тела есть величина равная сумме количества теплоты, полученной этим телом. и совершенной над ним работы. Второй закон (начало) термодинамики или закон энтропии утверждает, что в замкнутой системе энтропия может только возрастать. Применительно к энергии в экосистемах удобна следующая формулировка: процессы, связанные с превращениями энергии, могут происходить самопроизвольно только при условии, что энергия переходит из кон-центрированной формы в рассеянную, то есть деградирует. Мера количества энергии, ко-торая становится недоступной для использования, есть энтропия. Чем выше упорядочен-ность системы, тем меньше ее энтропия. Таким образом, любая живая система, в том чис-ле и экосистема, поддерживает свою жизнедеятельность благодаря, во-первых, наличию в окружающей среде в избытке даровой энергии (энергия Солнца); во вторых, способности за счет устройства составляющих ее компонентов эту энергию улавливать и концентрировать, а использовав - рассеивать в окружающую среду. Таким образом, сначала улавливание, а затем концентрирование энергии с переходом от одного трофического уровня к другому обеспечивает повышение упорядоченности, организации живой системы, то есть уменьшение ее энтропии. Весь запас энергии сосредоточен в массе органического вещества - биомассе, поэтому интенсивность образования и разрушения органического вещества на каждом из уровней определяется прохождением энергии через экосистему. Скорость образования органического вещества называют продуктивностью. Различают первичную и вторичную продуктивность. В любой экосистеме происходит образование биомассы и ее разрушение, при-чем эти процессы всецело определяются жизнью низшего трофического уровня - проду-центами. Все остальные организмы только потребляют уже созданное растениями органическое вещество и, следовательно, общая продуктивность экосистемы от них не зависит. Высокие скорости продуцирования биомассы наблюдаются в естественных и искусствен-ных экосистемах там, где благоприятны абиотические факторы, и особенно при поступлении дополнительной энергии извне, что уменьшает собственные затраты системы на поддержание жизнедеятельности. Такая дополнительная энергия может поступать в разной форме: например, на возделываемом поле - в форме энергии ископаемого топлива и работы, совершаемой человеком или животным. Таким образом, для обеспечения энергией всех особей сообщества живых организмов экосистемы необходимо определенное количественное соотношение между продуцентами, консументами разных порядков, детритофагами и редуцентами. Однако для жизнедеятельности любых организмов, а значит и системы в целом, только энергии недостаточно, они обязательно должны получать различные минеральные компоненты, микроэлементы, органические вещества, необходимые для построения молекул живого вещества. (Консументы - организмы, получающие питательные вещества и необходимую энергию, питаясь живыми организмами - продуцентами или другими консументами. Редуценты - организмы, получающие питательные вещества и необходимую энергию питаясь ос-танками мертвых организмов (животных, растений). Детритофаги - организмы, питающиеся мертвым органическим веществом вместе с содержащимися в нем микроорганизмами). 52-55-.Продуценты как компонент экосистемы./Продуценты как компонент экосистемы/Продуктивность фитоценоза и запасы биомассы/Консументы как компонент экосистемы/Редуциенты и дедритофаги как компоненты системы. Продуце́нты — организмы, способные синтезировать органические вещества из неорганических, то есть, все автотрофы. Это, в основном, зелёные растения (синтезируют органические вещества из неорганических в процессе фотосинтеза), однако некоторые виды бактерий-хемотрофов способны на чисто химический синтез органики без солнечного света. ПРОДУКТИВНОСТЬ ФИТОЦЕНОЗА — (количество органической массы (биомассы), производимое растениями данного фитоценоза на единицу площади за год. ФИТОМАССА (от греч. phyton -- растение илат. massa -- ком кусок), — общая масса живого органического вещества растений, составная часть биологической массы. Различают фитомассу надземную (стволы и ветви деревьев и кустарников, побеги, хвоя, листья, цветки) и подземную (корни, корневища, клубни, луковицы и др.). Соотношение надземной и подземной фитомассы в разных типах растительных сообществ существенно различается и может меняться в зависимости от времени года. Обычно фитомасса измеряется массой абсолютно сухого вещества на единицу площади или объема (г/м2, кг/га, т/га, г/м3 и т. д.), реже -- в энергетических единицах (ккал и др.). Важными показателями фитомассы являются ее объем, поверхность, химизм, калорийность, внутренняя и внешняя структура, фракционный состав, годичный прирост и др. Величина фитомассы растительного сообщества характеризует его организацию, биосферное значение. Среди естественных растительных сообществ суши наибольший запас фитомассы накапливают леса, растущие в благоприятных условиях, например во влажных тропиках, где фотосинтез древесных пород почти равномерно осуществляется в течение всего года. Минимальные запасы фитомассы -- в сообществах, находящихся в экстремальных условиях (тундра, пустыня). Отмирающая фитомасса пополняет ресурсы минерального питания растений, участвует в формировании почвенного гумуса, влияет на химизм верхних горизонтов почвы, на состав и обилие почвенных животных и микроорганизмов. Характер и размеры накопления фитомассы зависят не только от условий обитания, но и от биологических особенностей растений (в частности от продолжительности их жизни). Консументы (от лат. consumе — употреблять) — гетеротрофы, организмы, потребляющиее готовые органические вещества, создаваемые автотрофами (продуцентами). В отличие от редуцентов, консументы не способны разлагать органические вещества до неорганических[1]. К консументам относят животных, некоторые микроорганизмы, а также паразитические и насекомоядные растения. Классифицируют консументов первого, второго и других порядков, так как на каждом этапе передачи вещества и энергии в трофической цепи теряется до 90 %, экологические пирамиды редко состоят из более чем четырёх порядков консументов. Консументы первого порядка (первичные консументы) — растительноядные гетеротрофы (травоядные животные, паразитические растения), питаются непосредственно продуцентами биомассы[2]. Консументы второго порядка — хищные гетеротрофы (хищники, паразиты хищников), питаются консументами первого порядка[2]. Отдельно взятый организм может являться в разных трофических цепях консументом разных порядков, например, сова, поедающая мышь, является одновременно консументом второго и третьего порядка, а мышь — первого и второго, так как мышь питается и растениями, и растительноядными насекомыми. Любой консумент является гетеротрофом, так как не способен синтезировать органические вещества из неорганических. Термин «консумент (первого, второго и так далее) порядка» позволяет более точно указать место организма в цепи питания. Редуценты (например, грибы, бактерии гниения) также являются гетеротрофами, от консументов их отличает способность полностью разлагать органические вещества (белки, углеводы, липиды и другие) до неорганических (углекислый газ, аммиак, мочевина, сероводород), завершая круговорот веществ в природе, создавая субстрат для деятельности продуцентов (автотрофов). Редуце́нты (также деструкторы, сапротрофы, сапрофиты, сапрофаги) — микроорганизмы (бактерии и грибы), разрушающие отмершие остатки живых существ, превращающие их в неорганические и простейшие органические соединения. От животных-детритофагов редуценты отличаются прежде всего тем, что не оставляют твердых непереваренных остатков (экскрементов). Животных-детритофагов в экологии традиционно относят к консументам. В то же время все организмы выделяют углекислый газ и воду, а часто и другие неорганические (аммиак) или простые органические (мочевина) молекулы и таким образом принимают участие в разрушении (деструкции) органического вещества. Редуценты возвращают минеральные соли в почву и воду, делая их доступными для продуцентов-автотрофов, и таким образом замыкают биотический круговорот. Поэтому экосистемы не могут обходиться без редуцентов (в отличие от консументов, которые, вероятно, отсутствовали в экосистемах в течение первых 2 млрд лет эволюции, когда экосистемы состояли из одних прокариот). Исследованиями Н. И. Базилевич и др. (1993) установлено, что в наземных экосистемах различают две группы факторов, регулирующих деструкционные процессы, играющие весьма существенную роль в биологическом круговороте. Это прежде всего абиотические факторы — выщелачивание растворимых соединений, фотохимическое окисление органического вещества и реакции его механического разрушения вследствие замерзания—оттаивания. Эти факторы наиболее проявляются в надземных ярусах экосистем, а биотические факторы — в почвенном. Абиотические факторы деструкции характерны для аридных и семиаридных ландшафтов (пустыни, степи, саванны), а также для континентальных высокогорий и полярных ландшафтов. Биотические факторы деструкции — это в первую очередь сапротрофные организмы (беспозвоночные и позвоночные животные, микроорганизмы), населяющие почву и подстилку, причём ведущим фактором в наземных ландшафтах служит главным образом почвенная микрофлора. Детритофа́ги (от лат. detritus — распад, продукт распада и греч. φάγος — пожиратель) — животные и протисты, которые питаются разлагающимся органическим материалом — детритом. 56. Экологические пирамиды численности, массы, энергии. Экологическая пирамида — графические изображения соотношения между продуцентами и консументами всех уровней (травоядных, хищников, видов, питающихся другими хищниками) в экосистеме. Эффект пирамид в виде графических моделей разработан в 1927 году Ч. Элтоном. Выражается:
Построение пирамиды численности может быть затруднено в том случае, если разброс численности организмов разных уровней оказывается чрезвычайно велик. (Например, 500 тысячам злаков в основании пирамиды может соответствовать один конечный хищник.) Кроме того, пирамида может оказаться перевернутой (в том случае, если продуцент очень крупный, или в том случае, если большое число паразитов питаются на немногочисленных консументах).Пирамида биомасс отражает состояние экосистемы на момент отбора пробы и, следовательно, показывает соотношение биомассы в данный момент и не отражает продуктивность каждого трофического уровня (т. е. его способность образовывать биомассу в течение определенного промежутка времени). Поэтому в том случае, когда в число продуцентов входят быстрорастущие виды, пирамида биомасс также может оказаться перевернутой. Этих недостатков лишена пирамида энергии. Она позволяет сравнить продуктивность различных трофических уровней, поскольку учитывает фактор времени. Кроме того, она учитываем разницу в энергетической ценности различных веществ (например. 1 г жира дает почти в два раза больше энергии, чем 1 г глюкозы). Поэтому пирамида энергии всегда суживается кверху и никогда не бывает перевернутой. |