1. Экология как наука. Положение экологии в системе наук о природе
 Скачать 1.32 Mb. Скачать 1.32 Mb.
|

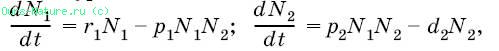 Схема отношения паразит – хозяин. где N1 – численность популяции хозяина; N2 – численность популяции паразита; r1 – удельная скорость увеличения популяции хозяина; d2 – удельная скорость гибели популяции паразита; p1 и р2 – константы. График процесса паразитической инвазии, построенный по таким уравнениям, обнаруживает, что в результате взаимодействия двух видов должны возникать осцилляции (колебания) с постоянной амплитудой, которая зависит от соотношения между скоростями увеличения численности двух видов. 14-15.Основные теоретические понятия в рамках концепций современной экологии. Аксио́ма (др.-греч. ἀξίωμα — утверждение, положение), постула́т — исходное положение какой-либо теории, принимаемое в рамках данной теории истинным без требования доказательства и используемое в основе доказательства других ее положений. Гипо́теза (др.-греч. ὑπόθεσις — предположение; от ὑπό — снизу, под + θέσις — тезис) — предположение или догадка; утверждение, предполагающее доказательство, в отличие от аксиом, постулатов, не требующих доказательств. Закон — фактологически доказанное утверждение (в рамках теории, концепции, гипотезы), объясняющее объективные факты; либо некое явление, обладающее общностью и повторяемостью, зафиксированное и описанное. Конце́пция (от лат. conceptio — понимание, система) — определённый способ понимания, трактовки каких-либо явлений, основная точка зрения, руководящая идея для их освещения[1]; система взглядов на явления в мире, в природе, в обществе[2]; ведущий замысел, конструктивный принцип в научной, художественной, технической, политической и других видах деятельности; комплекс взглядов, связанных между собой и вытекающих один из другого, система путей решения выбранной задачи. Концепция определяет стратегию действий. Моде́ль (фр. modèle, от лат. modulus — «мера, аналог, образец») — это система, исследование которой служит средством для получения информации о другой системе[1], это упрощённое представление реального устройства и/или протекающих в нём процессов, явлений. Прави́ло — описывает действие в рамках соблюдения принципов. Принцип или начало (лат. principium, греч. αρχή) — 1. Основополагающая истина, закон, положение или движущая сила, лежащая (лежащий) в основе других истин, законов, положений или движущих сил. 2. Руководящее положение, основное правило, установка для какой-либо деятельности. Теоре́ма (др.-греч. θεώρημα — «доказательство, вид; взгляд; представление, положение») — утверждение, для которого в рассматриваемой теории существует доказательство (иначе говоря, вывод). Уравнение – это равенство, которое выполняется лишь при некоторых значениях 16.Экологические факторы. Классификация экологических факторов. Факторы среды воздействуют на организм не по отдельности, а в комплексе, соответственно, любая реакция организма является многофакторно обусловленной. При этом интегральное влияние факторов не равно сумме влияний отдельных факторов, так как между ними происходят различного рода взаимодействия, которые можно подразделить на четыре основных типа:
Влияние факторов также зависит от природы и текущего состояния организма, поэтому они оказывают неодинаковое воздействие как на разные виды, так и на один организм на разных этапах онтогенеза: низкая влажность губительна для гидрофитов, но безвредна для ксерофитов; низкие температуры без вреда переносятся взрослыми хвойными умеренного пояса, но опасны для молодых растений. Классификации экологических факторов: По характеру воздействия
По происхождению: 1. Абиотические — факторы неживой природы: климатические: годовая сумма температур, среднегодовая температура, влажность, давление воздуха эдафические (эдафогенные): механический состав почвы, воздухопроницаемость почвы, кислотность почвы, химический состав почвы орографические: рельеф, высота над уровнем моря, крутизна и экспозиция склона химические: газовый состав воздуха, солевой состав воды, концентрация, кислотность физические: шум, магнитные поля, теплопроводность и теплоёмкость, радиоактивность, интенсивность солнечного излучения 2. Биотические — связанные с деятельностью живых организмов: фитогенные — влияние растений микогенные — влияние грибов зоогенные — влияние животных микробиогенные — влияние микроорганизмов 3. Антропогенные (антропические): физические: использование атомной энергии, перемещение в поездах и самолётах, влияние шума и вибрации химические: использование минеральных удобрений и ядохимикатов, загрязнение оболочек Земли отходами промышленности и транспорта биологические: продукты питания; организмы, для которых человек может быть средой обитания или источником питания социальные — связанные с отношениями людей и жизнью в обществе По расходованию:
По направленности:
17.Гипотеза компенсации экологических факторов и незаменимости фундаментальных факторов. Отсутствие или недостаток некоторых экологических факторов может быть компенсирован каким-либо другим близким (аналогичным) фактором. Организмы не являются «рабами» физических факторов (условий среды): они сами и приспосабливаются, и изменяют условия среды так, чтобы ослабить лимитирующее влияние тех или иных факторов. Некоторые моллюски (в частности, Mytilus galloprovincialis Lam.) при отсутствии (или дефиците) кальция могут строить свои раковины, частично заменяя кальций стронцием (при достаточном содержании всреде последнего). Полное отсутствие в среде фундаментальных экологических факторов (физиологически необходимых;например, света, воды, углекислого газа, питательных веществ) не может быть компенсировано (заменено) другими факторами. 18.Принцип лимитирующих факторов. Принцип лимитирующего фактора – избыток или недостаток одного абиотического фактора может повлечь за собой ограничение или остановку роста численности популяции в экосистеме, даже если значения других факторов оптимальны. Лимитирующий фактор – любой фактор, тормозящий рост популяции в экосистеме. Лимитирующие факторы для наземных экосистем: - температура; - вода; - свет; - питательные вещества в почве. Лимитирующие факторы для водных экосистем: - температура; - солнечный свет; - содержание растворенного кислорода; - соленость. 19.Влияние экологических факторов на организм. Закон минимума Ю.Либиха Закон, открытый Ю.Либихом ( 1840 ), согласно которому относительное действие отдельного экологического фактора тем сильнее, чем больше он находится по сравнению с другими факторами в минимуме ; по данному закону, от вещества, концентрация которого лежит в минимуме, зависят рост растений, величина и устойчивость их урожайности. Закон минимума Либиха гласит : рост растения зависит от того элемента питания, который присутствует в минимальном количестве. В изобилии присутствуют двуокись углерода и вода, а потому они не являются факторами, ограничивающими рост. А вот цинка в почве очень мало, потребность растения в нем невелика, и рост растения будет успешен до тех пор, пока не будет израсходован весь его запас. Поэтому наличие цинка является ограничивающим, или лимитирующим фактором. Ю.Одум :
Совокупность факторов воздействует сильнее всего на те фазы развития организмов, которые имеют наименьшую пластичность - минимальную способность к приспособлению. Толерантность (от греческого толеранция - терпение) - способность организмов выдерживать изменения условий жизни (колебания температуры, влажности, света). Диапазон толерантности. Чтобы выразить относительную степень толерантности, в экологии используют приставки стено- (от греч. stenos -узкий, тесный) и эври- (от греч. eurys - широкий), поли- (от греч. polys - многий, многочисленный) и олиго- (от греч. oligos - немногий, незначительный). Так (см. схему; Одум,1975) если в качестве фактора взять, например, температуру, то вид I - стенотермный и олиготермный, вид II - эвритермный, вид III - стенотермный и политермный: Организмы с широким диапазоном толерантности обозначают приставкой "Эври". Эврибионт - организм, способный жить при различных условиях среды. Например: эвритермный - переносящий широкие колебания температуры. С узким диапазоном - обозначают приставкой "Стено". Стенобионт - организм, требующий строго определённых условий среды. Например: форель - стенотермный вид, а окунь - эвритермный. Форель не выносит большие колебания температуры, если исчезнут все деревья по берегам горного потока, это приведет к повышению температуры на несколько градусов, в результате чего форель погибнет, а окунь выживет. 19. Роль тепла в жизни растений и животных. Эвритермные и стенотермные виды. Границами существования жизни являются температурные условия, при которых, не происходит денатурации, белков, необратимого изменения коллоидных свойств цитоплазмы, нарушения активности ферментов, дыхания. Для большинства организмов этот диапазон температур составляет от 0 до +50°С. Однако ряд организмов обладает специализированными ферментными системами и приспособлен к активному существованию при температурах, выходящих за указанные пределы. Виды, оптимальные условия жизнедеятельности которых приурочены к области высоких значений температур, относят к экологической группе термофилов. Термофильность характерна для многих бактерий, вызывающих самонагревание влажного зерна, сена, цианобактерии осцилатории, населяющей термальные источники Камчатки с температурой воды 85—93°С. Успешно переносят высокие температуры (65—80°С) несколько видов зеленых водорослей, накипные лишайники, семена пустынных растений, находящиеся в верхнем раскаленном слое почвы. Температурный предел представителей животного мира обычно не превышает +55—58°С (раковинные амебы, нематоды, клещи, некоторые ракообразные, личинки многих двукрылых). У многих видов растений и животных клетки сохраняют активность при температуре от 0 до -8°С. Такие организмы относятся к экологической группе криофилов (грен. Kryos —холод, лед). Кри-офилия характерна для многих бактерий, грибов, лишайников, членистоногих и других существ, обитающих в тундрах, арктических и антарктических пустынях, в высокогорьях, холодных полярных водах и т. п. Только у птиц и млекопитающих тепло, вырабатываемое в процессе интенсивного обмена веществ, служит достаточно надежным источником повышения температуры тела и поддержания ее на постоянном уровне независимо от температуры окружающей среды. Этому способствует хорошая тепловая изоляция, создаваемая шерстным покровом, плотным оперением, толстым слоем подкожной жировой ткани. Такие организмы называют гомойотермными (эндотермными, или теплокровными). Свойство эндотермности позволяет многим видам животных (белым медведям, ластоногим, пингвинам и др.) вести активный образ жизни при низких температурах. Температурная адаптация растений. Дня большинства наземных растений оптимальной является температура +25—30°С, а для таких требовательных к теплу растений, как кукуруза, фасоль, соя и другие виды тропического и субтропического происхождения, — +30—35°С. Следует иметь в виду, что для каждой фазы и стадии развития растений существует как оптимальный, так и верхний и нижний пределы температурного режима. При воздействии на растение высоких температур происходит сильное обезвоживание и иссушение, ожоги, разрушение хлорофилла, необратимые расстройства дыхания, наконец, тепловая денатурация белков, коагуляция цитоплазмы и гибель. Противостоять опасному влиянию экстремально высоких температур растения способны благодаря усиленной транспирации, накапливанию в цитоплазме защитных веществ (слизи, органических кислот и др.), сдвигам температурного оптимума активности важнейших ферментов, переходу в состояние глубокого покоя, а также занятию ими временных местообитаний, защищенных от сильного перегрева. Это означает, что у некоторых растений вся вегетация сдвигается на сезон с более благоприятными тепловыми условиями. Так, в пустынях и степях есть немало видов растений, начинающих вегетацию очень рано весной и успевающих ее закончить до наступления летней жары. Они переживают эти условия в состоянии летнего покоя — уже созрели семена или появились подземные органы —луковицы, клубни, корневища (тюльпаны, крокусы, мятлик луковичный и др.) Опасность низких температур для растений сводится к тому, что в межклетниках и клетках замерзает вода и, как следствие, происходит обезвоживание и механическое повреждение клеток, а затем коагуляция белков и разрушение цитоплазмы. Холод тормозит процессы роста растений, фотосинтеза, образования хлорофилла, снижает энергетическую эффективность дыхания, резко замедляет скорость развития. К Э. о. относятся все наземные растения умеренных и холодных поясов Земли. Среди животных Э. о. являются те обитатели указанных поясов, к-рые не могут мигрировать на зиму в более тёплые страны, не устраивают норы и не впадают на холодный период в спячку. Э. о., как правило, имеют более широкое распространение, чем стенотермные организмы, живущие в условиях относит, постоянных темп-р. К Э. о. относятся: волк, горностай, сокол-сапсан, азиатская саранча и мн. др. Среды, к которой приспособлены стенотермные животные, различна для животных разных видов: теплолюбивые могут жить только при относительно высокой температуре, обычно не ниже 20 °С (например, рифовые кораллы, ряд видов насекомых, большинство пресмыкающихся), холодолюбивые - при низкой, иногда близкой к 0 °С (например, некоторые виды арктических ракообразных, лососёвых рыб, ряд видов тюленей и др.). 21. Деление животных на группы по источникам тепла и способности к терморегуляции. Пойкилотермные и гомойотермные организмы. Представители большинства видов живых организмов не обладают способностью активной терморегуляции своего тела. Их активность зависит прежде всего от тепла, поступающего извне, а температура тела — от величины температуры окружающей среды. Такие организмы называют пойкилотермными (эктотермпыми). Пойкило-термия свойственна всем микроорганизмам, растениям, беспозвоночным и большей части хордовых. Только у птиц и млекопитающих тепло, вырабатываемое в процессе интенсивного обмена веществ, служит достаточно надежным источником повышения температуры тела и поддержания ее на постоянном уровне независимо от температуры окружающей среды. Этому способствует хорошая тепловая изоляция, создаваемая шерстным покровом, плотным оперением, толстым слоем подкожной жировой ткани. Такие организмы называют гомойотермными (эндотермными, или теплокровными). Свойство эндотермности позволяет многим видам животных (белым медведям, ластоногим, пингвинам и др.) вести активный образ жизни при низких температурах. Частный случай гомойотермии — гетеротермия — свойственна животным, впадающим в неблагоприятный период года в спячку или временное оцепенение (суслики, ежи, летучие мыши, сони и др.). В активном состоянии они поддерживают высокую температуру тела, а в случае низкой активности организма — пониженную, что сопровождается замедлением процессов обмена веществ и, как следствие, низкой теплоотдачей. Температурная адаптация животных. По сравнению с растениями животные обладают более разнообразными возможностями адаптации к воздействию различных температур. Обычно выделяют три основных пути температурных адаптации: 1) химическая терморегуляция (усиленное образование тепла в ответ на понижение температуры среды); 2) физическая терморегуляция (изменение уровня теплоотдачи, способность удерживать тепло или, наоборот, рассеивать его избыток); 3) поведенческая терморегуляция (избегание неблагоприятных температур путем перемещений в пространстве или изменение поведения более сложным образом). Пойкилотермные животные, в отличие от гомойотермных, характеризуются более низким уровнем обмена веществ даже при одинаковой температуре тела. Например, пустынная игуана при температуре +37°С потребляет кислорода в 7 раз меньше, чем грызуны такой же массы. По этой причине в теле пойкилотермных животных вырабатывается мало тепла, и, как следствие, возможности химической и физической терморегуляции ничтожны. Основным способом регуляции температуры тела у них являются особенности поведения — перемена позы, активный поиск благоприятных климатических условий, смена мест обитания, самостоятельное создание нужного микроклимата (сооружение гнезд, рытье нор и т. п.). Например, в сильную жару животные прячутся в тень, скрываются в норах, а некоторые виды пустынных ящериц и змей взбираются на кусты, избегая соприкосновения с раскаленной поверхностью почвы. Некоторые пойкилотермные животные способны поддерживать оптимальную температуру тела за счет работы мышц. Так, шмели разогревают тело путем активизации мышечных сокращений (дрожью) до +32 и 33°С, что дает им возможность взлетать и кормиться в прохладную погоду. Гомойотермия развилась из пойкилотермии путем интенсификации обменных процессов и усовершенствования способов регуляции теплообмена животных с окружающей средой. Эффективная регуляция поступления и отдачи тепла позволяет взрослым гомойотермным животным поддерживать постоянную оптимальную температуру тела во все времена года. Благодаря высокой интенсивности обмена веществ и выработке значительного количества тепла гомойотермные животные отличаются высокой способностью к химической терморегуляции, что особенно важно при действии холода. Однако поддержание температуры за счет возрастания теплопродукции требует большого расхода энергии, поэтому животные в холодный период года нуждаются в большом количестве пищи или тратят много жировых запасов, накопленных ранее. Физическая терморегуляция, обеспечивающая адаптацию к холоду не за счет дополнительной выработки тепла, а за счет сохранения его в теле животного, осуществляется путем рефлекторного сужения и расширения кровеносных сосудов кожи, меняющих ее теплопроводность, изменения теплоизолирующих свойств меха и перьевого покрова, регуляции испарительной теплоотдачи. 22. Видимый свет, ФАР, значение света для растений и животных. Видимое излучение — электромагнитные волны, воспринимаемые человеческим глазом[. Чувствительность человеческого глаза к электромагнитному излучению зависит от длины волны (частоты) излучения, при этом максимум чувствительности приходится на 555 нм (540 терагерц), в зелёной части спектра[2]. Поскольку при удалении от точки максимума чувствительность спадает до нуля постепенно, указать точные границы спектрального диапазона видимого излучения невозможно. Обычно в качестве коротковолновой границы принимают участок 380—400 нм (750—790 ТГц), а в качестве длинноволновой — 760—780 нм (385—395 ТГц) [1][3]. Электромагнитное излучение с такими длинами волн также называется видимым светом, или просто светом (в узком смысле этого слова). Фотосинтетическая активность радиации(ФАР) - часть доходящей до биоценозов солнечной радиации в пределах 400-700 нм спектра электромагнитных волн с различным КПД, используемая растениями для фотосинтеза. Инфракрасное излучение воспринимается всеми организмами, а лучи с длиной волны 1,05 мкм принимают участие в теплообмене растений. Ультрафиолет с длиной волны 0,25-0,3 мкм стимулирует образование витамина D у животных; с длиной волны 0,2-0,3 мкм губительно действует на некоторые микроорганизмы, в том числе болезнетворные; с длиной волны 0,38-0,4 мкм необходимо для фотосинтеза у растений. Благодаря озоновому экрану ультрафиолетовое и рентгеновское излучение частично задерживаются. Видимый свет оказывает комплексное влияние на организм: красные лучи - преимущественно тепловое воздействие; синие и фиолетовые - изменяют скорость и направление биохимических реакций. В целом видимый свет влияет на скорость роста и развития растений, на интенсивность фотосинтеза, на активность животных, вызывает изменение влажности и температуры среды, является важным сигнальным фактором, обеспечивающим суточные и сезонные биоциклы. Световой режим - один из ведущих абиотических факторов, определяющий особенности распределения и изменения интенсивности солнечной радиации, которая поступает к природным и искусственным экосистемам. Световой режим любого места обитания определяется различными факторами. Показатели светового режима - интенсивность света, его количество и качество. Интенсивность (сила света) - определяется количеством солнечной энергии, приходящейся на 1 см2 горизонтальной поверхности в 1 минуту. Для прямых солнечных лучей этот показательпочти не зависит от географической широты, однако на него влияют особенности рельефа местности. Например, на южных склонах интенсивность света всегда больше, чем на северных. Количество света - суммарная солнечная радиация, измеренная за астрономический год. Увеличивается от полюсов к экватору, сопровождается изменением его качества. Для светового режима также имеет значение количество отражаемого света. 23.Экологические группы растений и животных по отношению к свету. По требованию к условиям освещения принято делить растения на следующие экологические группы: 1) светолюбивые (световые), или гелиофиты, – растения открытых, постоянно хорошо освещаемых местообитаний; 2) тенелюбивые (теневые), или сциофиты, – растения нижних ярусов тенистых лесов, пещер и глубоководные растения; они плохо переносят сильное освещение прямыми солнечными лучами; 3) теневыносливые, или факультативные гелиофиты, – могут переносить большее или меньшее затенение, но хорошо растут и на свету; они легче других растений перестраиваются под влиянием изменяющихся условий освещения. Среди животных различают светолюбивые виды, или фотофилы, и тенелюбивые виды, или фотофобы; а также эврифотные, выносящие широкий диапазон освещенности, и стенофотные, переносящие узкоограниченные условия освещенности. Свет для животных, в первую очередь, необходимое условие зрительной ориентации в пространстве. Рассеянные, отраженные от предметов лучи, воспринимаемые органами зрения животных, дают им значительную часть информации о внешнем мире. Человек получает более 90% информации о внешнем мире через зрительный анализатор. В процессе эволюции происходило постепенное усложнение зрительных аналиаторов. Наиболее совершенные органы зрения - глаза позвоночных, головоногих моллюсков и насекомых. Они позволяют воспринимать форму и размеры предметов, их цвет, определять расстояние. Отдельные виды животных могут сильно различаться по способности воспринимать разные лучи солнечного спектра. Для человека область видимых лучей - от фиолетовых до темно-красных. Некоторые животные, например гремучие змеи, видят инфракрасную часть спектра и ловят добычу в темноте. Цветовое зрение широко распространено у позвоночных, ракообразных, насекомых и паукообразных. Способность к различению цвета, помимо наличия специальных фоторецепторов (колбочек), в значительной мере зависит от того, при каком спектральном составе излучения существует или активен вид. Большинство млекопитающих, ведущих происхождение от предков с сумеречной и ночной активностью, плохо различают цвета и видят все в черно-белом изображении. Особенности зрения животных зависят от характера местообитания и образа жизни. У постоянных обитателей пещер (троглобионтов) или почвенных животных (педобионтов) глаза могут быть полностью или частично редуцированы (жужелицы, протеи, крот, слепыш и др.). Многие глубоководные животные лишены глаз. В ряде случаев у глубоководных организмов выработалось способность к биолюминесценции (холодное свечение) за счет окисления сложных органических соединений (ночесветки, глубоководные рыбы, головоногие моллюски и др.). Биолюминесценция используется как средство для коммуникаций, а также хищниками для привлечения жертв и жертвами для отпугивания хищников. Важное значение для животных и для растений играет продолжительность освещения и ритм чередования темного и светлого периодов суток. Реакция организмов на сезонные изменения длины дня называется фотопериодизмом. Есть животные с положительным фототаксисом (идут на свет) и отрицательным. 24.Сигнальное значение света. Биологические ритмы. Сигнальное значение света отражается через фотодинамической эффект, то есть через те или иные двигательные реакции. Во многих гидробионтов четко выраженный Фототропизм, причем в планктонных форм он чаще положительный, у бентосных - отрицательный. У подавляющего числа фотонегативних бентосных животных личинки светолюбивы, благодаря чему время держатся в толще воды, где находят для себя более благоприятные условия (пища, кислород). Перемещение личинок тока воды обеспечивает малоподвижным бентосные формы возможность широкого расселения в водоеме. С возрастом личинки становятся фотонегативнимы и, опускаясь на дно, переходят к бентосных образа жизни. Знак фототропизма может меняться в зависимости от состояния внешней среды. В условиях резкого дефицита кислорода очень много представителей пресноводного бентоса из фотонегативних становятся фотопозитивнимы и, ориентируясь на свет, всплывают к поверхности, где респираторные условия лучше. Подобная картина наблюдается в случае резкого повышения концентрации ряда вредных веществ. Высокая освещенность как бы символизирует для животных чистоту воды, поскольку у дна она содержит больше растворенных веществ, чем у поверхности. В условиях сильного освещения фотопозитивни организмы могут приобретать негативный Фототропизм и предотвращать света. По этой причине в прозрачных водоемах во время высокого стояния солнца много водорослей перемещаются из самого поверхностного слоя воды на глубину нескольких метров, избегая действия вредного лишней радиации. Например, водоросли Gymnodinium kovalevskyi и Prorocentrum micans становятся фотонегативнимы при освещенности 20 тыс. лк. которая уже тормозит фотосинтез. Знак фототропизма не представляет собой постоянную свойство организмов, а изменяется, имея приспособительное значение. Ориентируясь на свет, гидробионты находят для себя наиболее выгодное положение в пространстве. Особенно большое значение это имеет для морских планктонных организмов, которые, осуществляя регулярные суточные миграции, ночью поднимаются к поверхности воды, а днем опускаются на глубину 100-200 м. В большинстве случаев начало подъема и опускания определяется временем наступления той или иной освещенности, и в Таким образом свет приобретает для организмов сигнального значения. Значительную роль играет свет и в проявлении вертикальных миграций в бентосных форм, когда они всплывают в толщу воды ради расселения, размножения и других нужд. Биологи́ческие ри́тмы — (биоритмы) периодически повторяющиеся изменения характера и интенсивности биологических процессов и явлений. Они свойственны живой материи на всех уровнях ее организации — от молекулярных и субклеточных до биосферы. Являются фундаментальным процессом в живой природе. Одни биологические ритмы относительно самостоятельны (например, частота сокращений сердца, дыхания), другие связаны с приспособлением организмов к геофизическим циклам — суточным (например, колебания интенсивности деления клеток, обмена веществ, двигательной активности животных), приливным (например, открывание и закрывание раковин у морских моллюсков, связанные с уровнем морских приливов), годичным (изменение численности и активности животных, роста и развития растений и др.) Классификация биоритмов по Ю. Ашоффу (1984 г.) подразделяется:
Инфрадианные ритмы - ритмы длительностью больше суток. Примеры: впадение в зимнюю спячку (животные), менструальные циклы у женщин (человек). Существует тесная зависимость между фазой солнечного цикла и антропометрическими данными молодежи. Акселерация весьма подвержена солнечному циклу: тенденция к повышению модулируется волнами, синхронными с периодом «переполюсовки» магнитного поля Солнца (а это удвоенный 11-летний цикл, то есть 22 года). В деятельности Солнца выявлены и более длительные периоды, охватывающие несколько столетий. Важное практическое значение имеет также исследование других многодневных (околомесячных, годовых и пр.) ритмов, датчиком времени для которых являются такие периодические изменения в природе, как смена сезонов, лунные циклы и др. Влияние (отражение) лунных ритмов на отлив и прилив морей и океанов. Соответствуют по циклу фазам Луны (29.53 суток) или лунным суткам (24.8 часов). Лунные ритмы хорошо заметны у морских растений и животных, наблюдаются при культивировании микроорганизмов. Психологи отмечают изменения в поведении некоторых людей, связанные с фазами луны, в частности, известно, что в новолуние растёт число самоубийств, сердечных приступов и пр. Ультрадианные - ритмы длительностью меньше суток. Примеры: концентрация внимания, изменение болевой чувствительности, процессы выделения и секреции, цикличность фаз, чередующихся на протяжении 6…8-часового нормального сна у человека. В опытах на животных было установлено, что чувствительность к химическим и лучевым поражениям колеблется в течение суток очень заметно. Циркадианные (Околосуточные) Центральное место среди ритмических процессов занимает циркадианный (циркадный) ритм, имеющий наибольшее значение для организма. Понятие циркадианного (околосуточного) ритма ввел в 1959 году Халберг. Он является видоизменением суточного ритма с периодом 24 часа, протекает в константных условиях и принадлежит к свободно текущим ритмам. Это ритмы с не навязанным внешними условиями периодом. Они врожденные, эндогенные, то есть обусловлены свойствами самого организма. Период циркадианных ритмов длится у растений 23-28 часов, у животных 23-25 часов. 25. Экологические группы организмов по отношению к воде. Экологические группы гидробионтов. Толща воды заселена организмами, которые обладают способностью плавать или удерживаться в определенных слоях. В связи с этим, водные организмы подразделяются на группы. Нектон (nektos – плавающий) – это совокупность пелагических активно передвигающихся животных, не имеющих непосредственной связи с дном. Это главным образом крупные животные, которые способны преодолевать большие расстояния и сильные течения воды. Они имеют обтекаемую форму тела и хорошо развитые органы движения. К типичным нектонным организмам относятся рыбы, кальмары, киты, ластоногие. К нектону в пресных водах кроме рыб относятся земноводные и активно перемещающиеся насекомые. Планктон (planktos – блуждающий, парящий) – это совокупность пелагических организмов, которые не обладают способностью к быстрым активным передвижениям. Как правило, это мелкие животные – зоопланктон и растения – фитопланктон, которые не могут противостоять течениям. В состав планктона включают и «парящие» в толще воды личинки многих животных. Плейстон (от греч. pleusis – плавание) – организмы, которые пассивно плавают на поверхности воды или ведут полупогруженный образ жизни получили название. Часто они используют как опору пленку поверхностного натяжения или образуют воздушные полости и другие поплавки. Типичными плейстонными животными являются сифонофоры, некоторые моллюски и др. Из растительных организмов к плейстону относятся саргассовые водоросли, ряски. Бентос (benthos – глубина) – это совокупность организмов, обитающих на дне (на грунте и в грунте) водоемов. Он подразделяется на зообентос и фитобентос. Большей частью представлен прикрепленными, или медленно передвигающимися, или роющими в грунте животными. На мелководье он состоит из организмов, синтезирующих органическое вещество (продуценты), потребляющих его (консументы) и разрушающих (ре-дуценты). На глубинах, где нет света, фитобентос (продуценты) отсутствует. В морском зообентосе доминируют фораминифоры, губки, кишечно-полостные, черви, плеченогие, моллюски, асцидии, рыбы и др. Более многочисленны бентосные формы на мелководьях. Их общая биомасса здесь может достигать десятков килограммов на 1 м2. Нейстон (от греч. neustos – плавающий) – сообщество организмов, обитающих у поверхностной пленки воды. Организмы, обитающие сверху поверхностной пленки – эпинейстон, снизу – гипонейстон. К нейстону относят также обитателей верхнего пятисантиметрового слоя воды. Нейстон составляют некоторые простейшие, одноклеточные водоросли, мелкие легочные моллюски, водомерки, вертячки, личинки комаров и др. Перифитон (от греч. peri – около и phyton – растение). Обрастатели, совокупность организмов, которые поселяются на подводных предметах или растениях и образуют так называемые обрастания на природных или искусственных твердых поверхностях – камнях, скалах, подводных частях судов, сваях, гидротехнических сооружениях (водоросли, усоногие раки, моллюски, мшанки, губки и др.). Экологические группы наземных организмов. По отношению к водному режиму среди наземных растений выделяются: Гигрофиты – обитающие на влажных местах (тропические растения, росянка, злаки, осоки). Мезофиты – растения умеренно увлажненных местообитаний (многие лесные травы, лиственные деревья, большинство сельскохозяйственных культур). Ксерофиты – растения сухих местообитаний, способные хорошо переносить засухи. Разделяются на суккулентов (способных накапливать в тканях большое количество воды – кактусы, алоэ, очиток молодило) и склерофитов (сухие, жесткие кустарники или травы – саксаул, верблюжья колючка, ковыли). Среди наземных животных по отношению к водному режиму можно также выделить три экологические группы: Гигрофилы – влаголюбивые, нуждаются в высокой влажности (мокрицы, комары, наземные моллюски, амфибии). Ксерофилы – сухолюбивые животные, не переносящие высокой влажности (верблюды, пустынные пресмыкающиеся). Мезофилы – животные, предпочитающие умеренную влажность (многие птицы, млекопитающие). 26. Вода как среда обитания организмов. Экологические особенности водных организмов. Водная среда, или гидросфера была первой из сред жизни, освоенной организмами. Это самая обширная среда обитания, занимающая 71% площади нашей планеты. Вода как среда обитания имеет ряд специфических свойств, таких, как большая плотность, сильные перепады давления, относительно малое содержание кислорода, сильное поглощение солнечных лучей и др. Водоемы и отдельные их участки различаются, кроме того, солевым режимом, скоростью горизонтальных перемещений (течений), содержанием взвешенных частиц. Для жизни придонных организмов имеют значение свойства грунта, режим разложения органических остатков и т. п. Поэтому наряду с адаптациями к общим свойствам водной среды ее обитатели должны быть приспособлены и к разнообразным частным условиям. Обитатели водной среды получили в экологии общее название гидробионтов. Они населяют Мировой океан, континентальные водоемы и подземные воды. Особенности гидросферы как среды обитания водных организмов, называемых гидробионтами, можно понять, лишь ознакомившись с физическими свойствами воды по сравнению с воздухом. К числу главнейших свойств воды относится плотность, которая примерно в 1220 раз превышает плотность воздуха. Следствием этого является наличие большого сопротивления движению гидробионтов, увеличение давления на них воды с возрастанием глубины, большая опорность, используемая водными организмами, а также высокие выталкивающая сила (архимедова сила) и вязкость. Плотность воды зависит от температуры. Максимальная плотность, равная 1 г/мл, наблюдается при +4 °С. При повышении или понижении температуры плотность воды уменьшается. При замерзании вода расширяется, увеличивая свой объем примерно на 11%. Благодаря этому свойству лед располагается на поверхности водоема, а более плотная вода с положительными температурами находится подо льдом. Самая плотная вода находится в придонной области, давая возможность для жизни донным организмам в зимнее время. У воды одна из самых высоких величин удельной теплоемкости. Чтобы изменить температуру 1 г. воды на 1 °С, нужно затратить 4,19 Дж тепла (в 500 раз больше, чем у воздуха). Поэтому вода, медленно нагреваясь и медленно остывая, уменьшает амплитуду суточных и сезонных колебаний температуры, стабилизируя ее. У воды высокая теплопроводность (в 30 раз выше, чем у воздуха), благодаря чему осуществляется равномерное распределение температуры в водной среде. Вода превосходный растворитель разнообразных минеральных веществ. В зависимости от количества растворенных солей выделяют пресные (до 0,5 г/л), солоноватые (0,5—16 г/л), морские (16—47 г/л) и пересоленные (47—350 г/л) воды. Природным водам свойствен определенный состав минеральных солей. Так, в пресных водах преобладают карбонаты, в морских — хлориды. С повышением солености воды возрастает ее плотность и понижается температура замерзания. В воде растворяются и газы. Однако кислорода в воде содержится в 30 раз меньше, чем при той же температуре в равном объеме воздуха. При интенсивном развитии гидробионтов в ночное время, когда нет обогащения воды за счет фотосинтеза водных растений, может возникнуть дефицит кислорода. Нередко это приводит к гибели водных организмов (например, заморы рыб). Поэтому кислород в водной среде — лимитирующий фактор. Второй лимитирующий фактор — свет. Освещенность быстро снижается с увеличением глубины. Идеально чистые воды имеют прозрачность 40–60 м, а силно загрязненные — не более 10 см (величину прозрачности определяют путем погружения белого диска в воду до предельной глубин его видимости). Поэтому наибольшее количество света получает поверхностный слой воды, в котором интенсивно осуществляется фотосинтез. Адаптации живых организме к водной среде. Главнейшими адаптациями водных организмов к жизни в водной среде являются следующие. Из-за низкого содержания кислорода в водной среде отсутствуют гидробионты с высоким уровнем процессов жизнедеятельности. Типичными ее обитателями являются организмы с непостоянной температурой тела, относящиеся к группе эктотермных организмов. В периоды недостатка кислорода они способны снижать интенсивность процессов жизнедеятельности, многие из них — вплоть до состояния анабиоза. Высокоорганизованные теплокровные (животные с высоким уровнем процессов жизнедеятельности — киты, дельфины, тюлени, морские котики и др.) живут в водной среде только благодаря дыханию атмосферным воздухом, который они вдыхают, периодически подымаясь из глубин к поверхности воды. Активно плавающие животные, способные преодолевать силу течения (самыми типичными представителями этой группы являются рыбы и головоногие моллюски) имеют обтекаемую форму тела и развитую мускулатура, позволяющую быстро передвигаться в водной среде. Микроскопически мелкие организмы используют воду как опору и находятся в состоянии свободного парения благодаря приспособлениям, снижающим удельную массу тела (отсутствие утяжеляющего скелета, наличие капелек жира или воздуха и др.) либо увеличивающим трение поверхности тела о воду (мелкие размеры тела, выросты покровов тела). Донную область заселили организмы бентоса (от греч. bentos — глубина). Многие из них имеют тяжелые известковые раковины (моллюски), мощную хитинизированную кутикулу (речной рак, крабы, омары, лангусты), органы прикрепления к грунту (присоски у пиявок, крючья у личинок ручейников, ризоиды и корни у растений). 27. Совместное действие факторов. Жизненные формы. Все экологические факторы в природе действуют на живые организмы одновременно. Причем это не простое суммирование, а сложное взаимовлияющее соотношение. Такая совокупность факторов называется их констелляцией. Точно так же и в формировании того или иного сообщества принимает участие целый комплекс факторов, определяющих условия существования. Важная экологическая закономерность – сцепленность, взаимозависимость различных факторов среды: изменение какого-либо одного фактора способно повлечь изменения остальных. С другой стороны, вернув измененный фактор в исходное состояние, можно (хотя и не всегда) восстановить до исходного состояния и другие, не подвергавшиеся прямому воздействию. Экологическим рядом называется совокупность растительных сообществ (фитоценозов), располагающихся соответственно нарастанию или убыванию (т. е. по градиенту) какого-либо фактора или группы факторов среды. Различают экологические ряды растений и их группировок, которые составляются по отношению к сухости (влажности) почвы, теневыносливости, тепловому режиму, засоленности, устойчивости к ветру и т. п. Пример экологического ряда древесных пород по возрастающей теневыносливости: лиственница – береза – сосна – осина – ива – ольха серая – липа – дуб – ясень – клен – ольха черная – ель – пихта. Морфологический тип приспособления животного или растения к основным экологическим факторам местообитания и к определенному образу жизни называют жизненной формой организма. Одна из первых классификаций жизненных форм была сделана для растений датским ботаником К. Раункиером по одному из признаков, имеющих большое приспособительное значение – положению почек или верхушек побегов в течение неблагоприятного периода года по отношению к поверхности почвы или снежного покрова:
Процентное распределение видов по жизненным формам Раункиер назвал биологическим (флористическим) спектром. Для разных зон и стран составлены биологические спектры, которые могут служить индикаторами климата. Жизненные формы, доминирующие в том или ином сообществе, могут выступать индикаторами условий обитания. Существует много различных классификаций жизненных форм животных: по особенностям размножения; способам передвижения или добывания пищи; по приуроченности к определенным экологическим нишам, ландшафтам, ярусам. Классификация жизненных форм по приспособленности к передвижению по Кашкарову Д.Н.:
Существуют классификации по влаголюбию, питанию, месту размножения и т. д. Все жизненные формы – это результат естественного отбора в ходе эволюции. В результате внешних или внутренних причин одно сообщество может разделиться на два, при этом нарастают различия между популяциями одного вида или между близкородственными видами, такой процесс называется дивергенцией. У неродственных форм может вырабатываться внешнее сходство, если эти виды ведут идентичный образ жизни в аналогичных условиях среды. Этот процесс получил название конвергенции. 28.Пища как экологический фактор. Классификация организмов по типу пищи и способу питания. Питание – это процесс потребления вещества и энергии. По типу питания живые организмы делятся: 1) по источнику энергии: - фототрофы (используют световую энергию), например, дерево; - хемотрофы (используют энергию химических связей), например, серобактерии, окисляющие H2S; 2) по источнику углерода: - автотрофы (синтезируют органику на свету), например, дерево; - гетеротрофы (используют в пищу готовые органические вещества), например, коровы; 3) по типу окисляемого субстрата: - литотрофы (окисляют неорганические соединения H2O, H2S, S, H2); - органотрофы (окисляют органические вещества). Гетеротрофы, в свою очередь, делятся: 1) по способу получения пищи: - фаготрофы (заглатывают куски) – животные; - осмотрофы (поглощают питательнее вещества через клеточную стенку) – грибы, бактерии; 2) по состоянию пищи: - биотрофы (питаются живыми организмами): * фитофаги (едят растения) – коровы, * зоофаги (едят животных) – тигр, - сапротрофы (питаются мертвой органикой): * сапрофиты (растения, живущие на мертвой органике), * сапрофаги (животные, питающиеся мертвой органикой), * копрофаги (питающиеся экскрементами), * некрофаги (питающиеся трупами), * детритофаги (питаются донной органикой в различной степени разложения). Некоторые организмы способны к смешанному типу питания – миксотрофы. Это представители отдела эвгленовых водорослей (зоологи эту группу считают своей и называют ее «растительные жгутиконосцы»). Эти организмы питаются на свету как растения, а в темноте как животные. 29.Определение понятия популяция. Примеры популяций животных и растений. Популяцией в экологии называют совокупность особей одного вида, находящихся во взаимодействии между собой и населяющих общую территорию. Члены популяции оказывают друг на друга не меньшее воздействие, чем физико-химические факторы среды или обитающие рядом организмы других видов. Поскольку контакты между особями одной популяции происходят чаще, чем между особями разных популяций, то именно внутривидовые отношения являются более напряженными, чем межвидовые. В популяциях проявляются в той или иной степени все формы биотических отношений, но наиболее ярко выражены мутуалистические и конкурентные. Мутуализм – крайняя степень ассоциации между особями, при которой каждый извлекает выгоду из связи с другим организмом. Специфические внутривидовые связи – это отношения, направленные на воспроизводство: между особями разных полов и между родительскими и дочерними поколениями. Популяция представляет собой форму внутривидовой организации или форму существования вида, обеспечивающую наиболее полное использование данной группой особей природных ресурсов территории, к которой популяция приурочена. Биологическое значение популяции состоит в наиболее полном и рациональном использовании энергетических ресурсов, позволяющем обеспечить оставление потомства. При половом размножении обмен генами превращает популяцию в целостную генетическую систему. Если размножение происходит вегетативным путем (побегами, почками и т.д.) или другими способами, то популяция представляет собой систему клонов, или чистых линий, совместно использующих среду. В современной биологии популяция рассматривается как элементарная единица в процессе эволюции, способная реагировать на изменения среды перестройкой своего генофонда. Говоря об экологических популяциях, необходимо отметить большое разнообразие их масштабов. У одного и того же вида в разной среде популяции могут сильно различаться. Эти различия обусловлены: а) площадью ареала популяции – они могут занимать территорию, сравнимую по площади с материком (популяции песца, кряквы), и могут ограничиваться несколькими квадратными метрами (некоторые амфибии и моллюски); б) количеством особей, образующих популяцию, – популяция может объединять миллионы особей (комары) или всего несколько десятков животных (крупные хищники); в) количеством микропопуляций – одни популяции представлены множеством микропопуляций, приуроченных к разным биотопам, другие – едины в пространственном отношении. Таким образом, популяции представляют собой весьма разнообразные видовые группировки, количество и особенности которых соответствуют пестроте и условиям местообитания, специфическим свойствам среды и биологии самих животных. 30.Численность и плотность популяций. Методы абсолютного и относительного определения плотности популяций. Индексом плотности называется любая однозначно и четко регистрируемая величина, эквивалентно связанная с численностью особей. Индексом плотности может быть и число фекалий животных на единицу маршрута, и количество силуэтов птиц, появляющихся на фоне диска Луны за единицу времени, и любая другая оценка плотности популяции линейно связанная с ее численностью. Именно обязательное соблюдение наличия прямолинейной связи между индексом плотности и абсолютной численностью популяции лежит в основе двух предложенных ниже методов оценки ее численности. Потому что в этом случае можно ожидать, что при изъятии или добавлении особей в исследуемой популяции индекс плотности будет пропорционально снижаться или возрастать, соответственно. Для получения достоверных результатов необходимо соблюдать ряд важных требований: 1. Границы участка обязательно должны быть четкие и непроницаемые для животных. 2. Смертность и рождаемость между периодами исследования должны быть по возможности минимальными. 3. Практически должна отсутствовать эмиграции и иммиграция животных. 4. Промежуток времени между обследованиями должен быть по возможности минимальным. 5. Достаточно высокой должна быть точность оценок индекса плотности. 1.Метод двукратного использования индексов Данный метод заключается в следующем. В конкретных условиях определяется индекс плотности (i1) популяции. Затем к этой популяции добавляется (или из нее изымается) известное количество животных (C). Определяется новое значение индекса плотности (i2). Оценка абсолютной численности животных тогда может быть определена по формуле (Коли, 1979):  Если животные добавляются, то величина C входит в формулу 1.1 со знаком плюс (+), если изымаются — со знаком минус (-). 2. Метод Келкера Если популяция сама оказывается достаточно гетерогенной (т.е. в ней можно выделить два или более четко различимых класса по окраске, по полу, по возрасту и т.п.), то численность такой популяции может быть определена, используя метод селективного изъятия (или добавления) Келкера (Kelker, 1940; 1944; Коли, 1979). Данный метод может быть использован не только при наличии двух (или более) классов среди животных исследуемой популяции, но также и при наличии двух (или более) видов, совместно обитающих на одной территории (они должны быть экологически сходными). Последовательность этапов определения абсолютной численности с использованием метода Келкера заключается в следующем. 1. Вначале бескровным образом отлавливается выборка животных и в ней определяется относительная доля особей каждого класса. 2. Далее из этой популяции изымается (или к ней добавляется) фиксированное количество особей одного из классов (или обоих классов). 3. Отлавливается новая выборка и в ней снова определяется относительная доля особей каждого класса. 4. Исходную численность популяции можно тогда определить по формуле:  , ,где X, Y — два класса животных в популяции; N1 — численность популяции при первом учете; p1 — доля особей, принадлежащих классу X при первом учете; p2 — доля особей, принадлежащих классу X при втором учете; CX, CY — число особей классов X и Y, соответственно, добавляемое (или изымаемое) из популяции между двумя последовательными обследованиями. Если особи добавляются, то CX, CY — положительные величины, если изымаются — отрицательные. Если особи какого-то из классов не добавляются и не изымаются, то эта величина равна нулю. Величина C = CX - CY. Данный метод нельзя использовать в тех случаях, если особи в популяции не распределены равномерно. 3. Метод неселективного изъятия Данный метод используется для оценки абсолютной численности животных, обитающих на ограниченной территории. Он может быть применен для оценки численности насекомых на определенном участке луга, млекопитающих в локальной популяции и т.п. В основе использования метода неселективного изъятия лежит явление постепенного снижения вероятности встречаемости животного в серии последовательных отловов, вызванное снижением численности популяции в результате изъятия из нее особей. Таким образом, при применении данного метода животных отлавливают, подсчитывают их количество и не выпускают до конца исследования. Затем производят еще 3-4 последовательных отлова по аналогичной методике, при этом число отловленных животных постепенно уменьшается, вследствие уменьшения их общего количества на исследуемой территории. Если теперь построить график зависимости числа отловленных животных при каждом отлове от общего числа ранее отловленных, то по нему можно найти оценку исходной численности популяции (Грин и др., 1990). Однако довольно часто встречаются случаи, когда особи обладают высокой миграционной активностью и место отловленных резидентных особей сразу же занимают иммигранты. (Подобная ситуация может встретиться, например, при отловах мелких млекопитающих.) Тогда совокупное число отловленных животных не будет снижаться при каждом последующем отлове, что даст заниженную оценку коэффициента b и, следовательно, завысит оценку начальной численности популяции. Метод трехкратных отловов Бейли позволяет, помимо оценки численности популяции, оценить также показатели рождаемости и смертности в течение периода исследования (Bailey, 1951; 1952; Коли, 1979). Кроме того, он применим в том случае, если животные довольно активно перемещаются. Для того чтобы оценить численность популяции с помощью метода Бейли необходимы иметь данные по двум последовательным мечениям и двум последовательным отловам. Таким образом, в момент t0 производится отлов выборки животных. Их всех единообразно метят и выпускают обратно. Спустя некоторое время в момент t1 отлавливается новая выборка, в ней подсчитывается количество меченых особей. Остальные (немеченые) животные из этой выборки также метятся, но на этот раз новой меткой, отличной от первой. Спустя время в момент t2 вновь отлавливается выборка животных и в ней подсчитывается число особей, меченых в первый раз и во второй. Оценить численность популяции можно и проводя многократные отловы, при этом производя единообразное мечения всех ранее немеченых животных при каждом последовательном отлове (Poole, 1974). В этом случае можно ожидать, что с каждым последовательным отловом и мечением в популяции будет расти доля меченых особей вплоть до того момента, когда все особи в данной популяции окажутся с метками. Таким образом, на основании оценки доли меченых особей в популяции (полученной с помощью выборочных данных) в зависимости от совокупного числа всех ранее меченых животных можно найти оценку величины популяции – ее численность, при которой доля меченых животных должна достичь 100 %. Метод Джолли-Зебера Если при каждом последовательном отлове особям присваиваются индивидуальные метки, то в этом случае можно оценить численность популяции (а также целый набор других экологических популяционных показателей) на основе метода Джолли-Зебера (Jolly, 1965; 1982; Seber, 1965; Begon, 1979; Коли, 1979). Использование этого метода требует, чтобы в распоряжении исследователя были подробные сведения относительно дат мечения и отловов каждого конкретного животного. 31-12.Рост и скорость роста популяции./ Рождаемость и смертность в популяции. Естественные популяции — это не раз и навсегда застывшая совокупность особей, а динамическое единство находящихся во взаимоотношениях организмов. Изменение в численности, структуре и распределении популяций как реакция на условия окружающей среды называется динамикой популяции. Динамика популяций в упрощенном варианте может быть описана такими показателями, как рождаемость и смертность. Это наиболее важные популяционные характеристики, на основании анализа которых можно судить об устойчивости и перспективном развитии популяции. Рождаемость определяется как число особей, рожденных в популяции за некоторый промежуток времени (час, день, месяц, год). Термин «рождаемость» относится к особям любых видов, независимо от способов появления их на свет: будь это прорастание семян подорожника или овса, появление детенышей из яиц у курицы или черепахи, рождение потомства у слона, кита либо человека. Экологи выделяют максимальную рождаемость в условиях отсутствия лимитирующих экологических факторов (добиться этого весьма сложно, даже невозможно). Под максимальной рождаемостью понимается теоретически возможный максимум скорости образования новых особей в идеальных условиях. Размножение организмов сдерживается только их физиологическими особенностями. Теоретическая скорость размножения различных видов может быть довольно высокой. В отличие от максимальной экологическая, или реализованная, рождаемость (или просто рождаемость) характеризует прирост или увеличение численности популяции при фактических или специфических условиях среды. Смертность — это число особей, погибших в популяции за единицу времени. Подобно рождаемости, смертность можно выразить числом особей, погибших за данный период (число смертей в единицу времени), или в виде удельной смертности для всей популяции (или ее части). При определении смертности популяции учитываются все погибшие особи независимо от причины смерти (умерли ли они от старости или погибли в когтях хищника, отравились ядохимикатами или замерзли и т.д.). Общие изменения численности популяции складываются за счет четырех явлений: рождаемости, смертности, вселения и выселения особей (иммиграция и эмиграция). рвая характеризуется общим числом родившихся особей. Удельную рождаемость вычисляют как среднее изменение численности особей за определенный период времени, деленное на их первоначальное число. Величина рождаемости зависит от многих причин. Большую роль играет плодовитость особей. Смертность в популяциях также зависит от многих причин: генетически запрограммированной длительности жизни особей, их генетической и физиологической полноценности, влияния неблагоприятных физических условий среды, воздействия хищников, паразитов, болезней и т.п. Наиболее часто в природе встречается повышенная смертность особей в ранний период жизни. Расселение (выселение) особей из популяции или пополнение ее пришельцами — закономерное явление, основанное на одной из важнейших биологических черт вида — его расселительной способности. В каждой популяции часть особей регулярно покидает ее (дисперсия популяции), пополняя соседние или заселяя новые, еще не занятые видом территории. Современная теория рассматривает темп роста численности популяций как авторегулируемый процесс. Любой популяции организмов в конкретных условиях свойствен определенный средний уровень численности, вокруг которого происходят колебания. В одном случае темп роста с самого начала высокий и постоянный, не зависящий от нарастающей плотности, что соответствует лавинообразному, по экспоненте, увеличению численности популяции (рис. 6.2а). Его графически описывает простейшая кривая, характеризующая изменение численности популяции, движущейся к равновесию, при условии изобилии пищи. При достижении же определенной плотности рост популяции прекращается. Если лимитирующий фактор среды обитания действует весьма быстро, то рост популяции прекращается внезапно (кривая «Б» на рис. 626). Однако среда обитания оказывает ограничивающее влияние не резко, а постепенно, что приводит к 5-образной кривой роста (рис. 6.2#). Такая форма кривой роста наблюдается при внедрении популяции в новую территорию. В этом случае вначале происходит ускоренный рост (по логарифмическому закону). Затем, под влиянием сопротивления среды обитания, рост замедляется, и в популяции наступает фаза равновесия. Если же популяция испытывает внешние воздействия (например, нападение хищников), то при постоянной удельной скорости изъятия особей в природе устойчиво существуют взаимодействующие популяции хищник — жертва (кривая «Г» на рис. 6.2и), но при меньшем уровне численности: Nиз меньше Nmax. 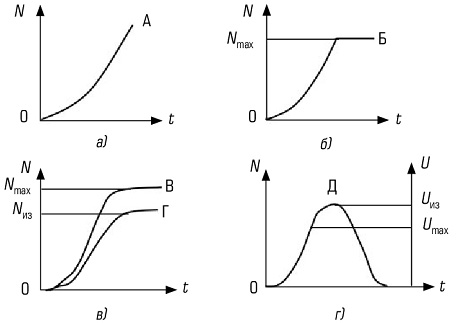 Кривые роста популяции: А — экспоненциальная; 5 — экспоненциальная с прекращением роста; В — логистическая; Г — логистическая с изъятием особей без превышения квоты; Д — с превышением квоты. N — численность популяции (Nmах — максимальная); (Uиз- реальная скорость изъятия, Umax — критическая для популяции скорость изъятия продукции; t — время Удельная скорость изъятия — число изъятых особей в единицу времен, отнесенных к численности популяции. Если же человек изымает биопродукцию из популяции с постоянной (интегральной, но не удельной) скоростью, то возникает понятие квоты. Следовательно квота представляет собой скорость отлова. Когда квота не превышает установленной критической величины, то равновесие популяции сохраняется. В этом случае отлов можно вести сколь угодно долго без губительных последствий для популяции. S-образная кривая называется логистической кривой роста, поскольку она получена путем интегрирования уравнения, основанного на логически обоснованных допущениях. Если квота превышает критическую величину отлова, то происходит полный отлов популяции за конечное время: популяция не успевает самовосстановиться и гибнет (рис. 6.2г) Весьма интересны для ученых-экологов циклические популяции, подверженные закономерным колебаниям численности. Однако единой теории удовлетворительно объясняющей закономерности в циклических популяциях пока нет. Поддержание определенной плотности получило название гомеостаза популяций. В основе способностей популяций к гомеостазу лежат изменения физиологических особенностей, роста, поведения каждой особи в ответ на увеличение или уменьшение числа членов популяции, к которой она принадлежит. Механизмы популяционного гомеостаза зависят от экологической специфики вида, его подвижности, степени воздействия хищников и паразитов и др. У одних видов они могут проявляться в жесткой форме, приводя к гибели избытка особей, у других — в смягченной, например, в понижении плодовитости на основе условных рефлексов. Другой механизм ограничения численности популяций — такие изменения физиологии и поведения при увеличении плотности, которые, в конечном счете, приводят к проявлению инстинктов массовой миграции. Наиболее эффективным механизмом сдерживания роста численности популяции на данном ареале является определенная система инстинктов — мечение и охрана участков, не допускающие размножения на них «чужих» особей. Генетические процессы в популяциях. В настоящее время известно, что все природные популяции гетерогенны и насыщены мутациями. Генетическая гетерогенность любой популяции при отсутствии давления внешних факторов должна быть неизменной, находиться в определенном равновесии. Положение о генетическом единстве популяции является одним из наиболее важных выводов популяционной генетики: любая популяция представляет сложную генетическую систему, находящуюся в динамическом равновесии. Прирост популяции - разница между рождаемостью и смертностью; прирост может быть положительным, нулевым и отрицательным. Скорость роста популяции — изменение численности популяции в единицу времени. Зависит от показателей рождаемости, смертности, миграции и эмиграции. 33.Таблицы и кривые выживания. Кривые выживания Один из основных факторов, влияющих на размеры популяции, -это процент особей, погибающих до достижения половой зрелости (смертность в до-репродуктивном периоде). В пределах данного вида эта величина гораздо более изменчива, чем плодовитость. Для того чтобы численность популяции оставалась постоянной, в среднем только два потомка каждой пары должны доживать до репродуктивного возраста. Кривую выживания можно получить, если начать некоторой популяции новорожденных особей и затем отмечать число выживших в зависимости от времени. По вертикальной оси обычно откладывают или абсолютное число выживших особей или их процент от исходной популяции: 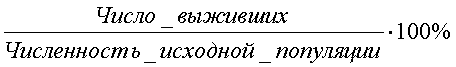 Каждому виду свойственна характерная кривая выживания, форма которой отчасти зависит от смертности неполовозрелых особей. 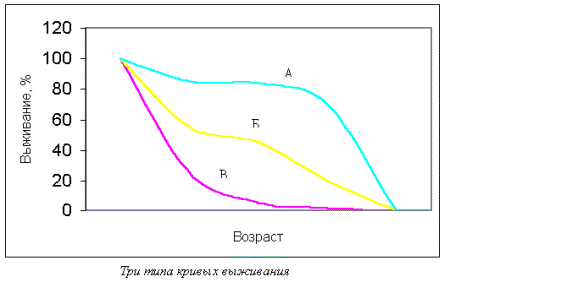 Большинство животных и растений подвержено старению, которое проявляется в снижении жизненности с возрастом после периода зрелости. Как только начинается старение, вероятность наступления смерти в определенный промежуток времени возрастает. Непосредственные причины смерти могут быть разными, но в основе их лежит уменьшение сопротивляемости организма к действию неблагоприятных факторов, например болезням. Кривая А на рисунке очень близка к идеальной кривой для популяции, в которой старение служит главным фактором, влияющим на смертность. Свойственна однолетним культурным растениям, например пшенице, когда все растения на данном поле стареют одновременно. Кривая типа Б характерна для популяций организмов с высокой смертностью в ранний период жизни, например для горных овец или для популяции человека в стране, где широко распространены голод и болезни. Плавная кривая типа В может быть получена, если смертность постоянна в течение всей жизни организмов (50% за определенную единицу времени). Это может быть тогда, когда главным фактором, определяющим смертность, становится случай, причем особи гибнут до начала заметного старения.. Очень сходная кривая характерна для популяций некоторых животных (например, гидры), не подвергающихся особой опасности в раннем возрасте. Для большинства беспозвоночных и растений тоже характерна кривая такого типа, но высокая смертность среди молодых особей приводит к тому, что начальная часть кривой спускается еще более круто. Вычерчивая кривые выживания для различных видов, можно определять смертность для особей разного возраста и таким образом выяснять, в каком возрасте данный вид наиболее уязвим. Установив причины смерти в этом возрасте, можно понять, как регулируется величина популяции. Таблицы выживания. Демографические таблицы. Как мы сказали, важнейшими статическими характеристиками популяции являются ее половой состав (соотношение особей разного пола) и возрастной состав (соотношение особей разного возраста). Эти параметры принято описывать с помощью демографических таблиц. Первая такая таблица была построена основателем демографии Джоном Грантом в XVII веке на основании данных о смертности жителей Лондона, которые церковные приходы собирали, чтобы вовремя обнаруживать начало эпидемий чумы. Демографические таблицы удобны для наблюдения за динамикой рождаемости и смертности в разных возрастных и (или) половых группах. Одним из способов их построения (табл. 4.3.1) является наблюдение за судьбой определенной группы особей (когорты), рожденных за короткий промежуток времени, и регистрация возраста наступления смерти всех членов когорты. Таблица 4.3.1. Демографическая таблица популяции морского желудя (Balanus glandula) — представителя усоногих ракообразных (Connell, 1970 по Гилярову, 1987)
|