1. Экология как наука. Положение экологии в системе наук о природе
 Скачать 1.32 Mb. Скачать 1.32 Mb.
|

|
| Возрастная группа | Количество человек в каждой возрастной группе | Количество умерших в каждой возрастной группе | Смертность в расчете на 1000 человек |
| 0-1 | 173 400 | 1 651 | 9,52 |
| 1-4 | 685 900 | 340 | 0,5 |
| 5-9 | 876 600 | 218 | 0,25 |
| 10-14 | 980 300 | 234 | 0,24 |
| 15-19 | 1 164 100 | 568 | 0,49 |
| 20-24 | 1 136 100 | 619 | 0,54 |
| 25-29 | 1 029 300 | 578 | 0,56 |
| 30-34 | 933 000 | 662 | 0,71 |
| 35-39 | 739 200 | 818 | 1,11 |
| 40-44 | 627 000 | 1 039 | 1,66 |
| 45-49 | 622 400 | 1 664 | 2,67 |
| 50-54 | 615 100 | 2 574 | 4,18 |
| 55-59 | 596 000 | 3 878 | 6,51 |
| 60-64 | 481 200 | 4 853 | 10,09 |
| 65-69 | 413 400 | 6 803 | 16,07 |
| 70-74 | 325 600 | 8 421 | 25,86 |
| 75-79 | 235 100 | 10 029 | 42,66 |
| 80-84 | 149 300 | 10 824 | 72,5 |
| 85 и более | 119 200 | 18 085 | 151,7 |
На основании демографических таблиц строят демографические пирамиды. На вертикальной шкале откладывают возрастные интервалы, по левой стороне, в виде столбчатой диаграммы, — численность самцов, по правой — самок. При этом наглядной становится разница смертности в разных возрастных категориях и у разных полов.
Демографические пирамиды помогают зримо представить историю популяции. Рассмотрите такую пирамиду для населения России (рис. 4.3.1). Вы можете увидеть, например, как на ней отражается снижение рождаемости во время Великой Отечественной войны. «Эхо» этих событий проявилось даже через поколение и, в меньшей степени, через два. Количество людей, которые по своему возрасту являются детьми и внуками тех, кто появлялся на свет во время войны, оказывается меньше численности близких к ним возрастных групп.
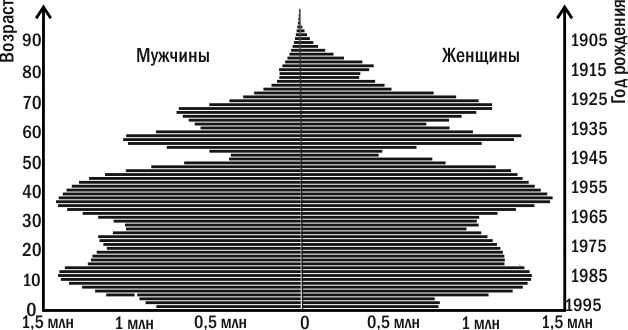
Демографические таблицы дают материал для построения кривых выживания, описанных выше.
34.Экспоненциальный и логистический рост. Математические формулы того и другого роста. Емкость среды.
Прирост популяции пропорционален численности особей в ней, то есть ΔNN, где N — численность популяции, а ΔN — ее изменение за определенный период времени. Если этот период бесконечно мал, можно написать, чтоdN/dt=r×N, где dN/dt — изменение численности популяции (прирост), а r — репродуктивный потенциал, переменная, характеризующая способность популяции увеличивать свою численность. Приведенное уравнение называется экспоненциальной моделью роста численности популяции.
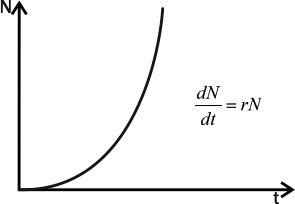
Величину r называют иногда мальтузианским параметром. Английский священник Томас Мальтус был первым, кто обратил внимание на то, что численность населения растет в геометрической прогрессии. Именно знакомство с его работой подтолкнуло и Чарльза Дарвина, и Альфреда Уоллеса к догадке о том, что потомство любых организмов должно «прореживаться» естественным отбором.
Естественно, экспоненциальный рост популяции не может быть вечным. Рано или поздно ресурс исчерпается, и рост популяции затормозится. Каким будет это торможение? Практическая экология знает самые разные варианты: и резкий взлет численности с последующим вымиранием популяции, исчерпавшей свои ресурсы, и постепенное торможение прироста по мере приближения к определенному уровню. Проще всего описать медленное торможение. Простейшая описывающая такую динамику модель называется Логистической и предложена (для описания роста численности популяции человека) французским математиком Ферхюльстом еще в 1845 году. В 1925 году аналогичная закономерность была заново открыта американским экологом Р. Перлем, который предположил, что она носит всеобщий характер.
В логистической модели вводится переменная K — емкость среды, равновесная численность популяции, при которой она потребляет все имеющиеся ресурсы. Прирост в логистической модели описывается уравнением dN/dt=r×N×(K-N)/K.
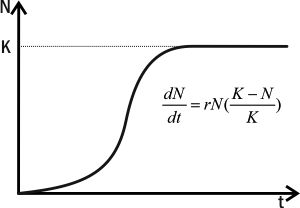
Пока N невелико, на прирост популяции основное влияние оказывает сомножитель r×N и рост популяции ускоряется. Когда становится достаточно высоким, на численность популяции начинает оказывать основное влияние сомножитель (K-N)/K и рост популяции начинает замедляться. Когда N=K, (K-N)/K=0 и рост численности популяции прекращается.
При всей своей простоте логистическое уравнение удовлетворительно описывает много наблюдаемых в природе случаев и до сих пор с успехом используется в математической экологии.
Предельная нагрузка биологического вида на среду обитания (Ёмкость среды) — это максимальный размер популяции вида, который среда может безусловно стабильно поддерживать, обеспечивать пищей, укрытием, водой и другими необходимыми благами.
35.Половой и возрастной состав. Генетический полиморфизм популяции.
Возрастной состав - соотношение в популяции особей разного возраста. В быстро растущей популяции обычно велика доля молоди, а в популяции, численность которой сокращается, обычно велика доля взрослых и стареющих особей.
Если численность популяции растет по экспоненциальному закону (в геометрической прогрессии), в ней устанавливается постоянный возрастной состав или, иначе, стабильная возрастная структура. В.с.п. является важнейшей характеристикой популяции человека.
Половой состав популяций - соотношение особей разных гендеров в популяции -обусловливается эволюционно закрепленными механизмами формирования первичного (на момент зачатия), вторичного (на момент рождения) и третичного (во взрослом состоянии) соотношения полов. В качестве примера рассмотрим изменение полового состава популяции людей. На момент рождения оно составляет 106 мальчиков на 100 девочек, в возрасте 16—18 лет выравнивается, в возрасте 50 лет насчитывает 85 мужчин на 100 женщин, а в возрасте 80 лет — 50 мужчин на 100 женщин.
ГИПОТЕЗА ДИФФЕРЕНЦИРОВАННОЙ СПЕЦИАЛИЗАЦИИ ПОЛОВ Геодокяна - дифференциация полов происходит по двум основным направлениям эволюции: изменению (мужской пол) и сохранению (женский пол). Чем больше в популяции женских особей, тем лучше сохраняется ее генотипическая структура; чем больше мужских особей, тем больше скорость или величина изменения этой структуры. Поскольку на разных этапах эволюции и в разных условиях среды требуется различная эволюционная пластичность популяции, то существует оптимальное соотношение полов (для групп половой зрелости).
Генетическое разнообразие или генетический полиморфизм — разнообразие популяций по признакам или маркерам генетической природы. Один из видов биоразнообразия. Генетическое разнообразие представляет собой важный компонент генетической характеристики популяции, группы популяций или вида. Генетическое разнообразие, в зависимости от выбора рассматриваемых генетических маркеров, характеризуется несколькими измеряемыми параметрами:
1. Средняя гетерозиготность.
2. Число аллелей на локус.
3. Генетическое расстояние (для оценки межпопуляционного генетического разнообразия)
Средняя гетерозиготность - параметр генетического разнообразия, описывающий, какую долю в популяции составляют особи, гетерозиготные по изучаемым маркерам, с усреднением этого параметра по набору использованных маркеров.
Число аллелей на локус. Этот параметр, как правило, используется для оценки генетического разнообразия по маркерам, имеющим более двух описанных аллельных состояний, например, для микросателлитных локусов.
Генетическое расстояние. Параметр описывает степень различия и разнообразия между популяциями по наличию/отсутствию или частотам аллелей используемых маркеров.
Генетическая изменчивость в популяции предоставляет исходный материал для действия естественного отбора и генетического дрейфа, то есть, является необходимым элементом для микроэволюционных процессов. В частности, известны работы о неэффективности отбора в чистых линиях (при отсутствии генетического разнообразия). С другой стороны, генетическая изменчивость сама по себе является продуктом действия факторов микроэволюции. Генетическое разнообразие имеет большое значение для экологической пластичности популяций. Наличие нескольких аллелей по аллозимным локусам в популяции позволяет этой самой популяции адаптироваться к варьирующим условиям, в которых наличие у особей тех или иных аллелей даёт преимущество. Например, два широко распространённых у Drosophila melАnogaster варианта гена алкогольдегидрогеназы оказывают в гомозиготном состоянии альтернативно полезное или вредное воздействие, в зависимости от температурных условий среды.
36-38.Размещение особей в популяциях по отношению к друг другу. /Пространственная структура популяции у оседлых животных. /Пространственная структура популяции у кочующих животных.
Типы пространственного распределения.
Равномерный, характеризуется равным удалением каждой особи от всех соседних; величина расстояния между особями соответствует порогу, за которым начинается взаимное угнетение. То есть этот тип распределения в наибольшей степени соответствует задаче полного использования ресурсов при минимальной степени конкуренции (в природе такой тип встречается достаточно редко). Например, одновидовые заросли некоторых растений, в уплотненных популяциях некоторых сидячих беспозвоночных. Диффузный тип распределения, особи распределены в пространстве неравномерно, случайно. Расстояние между особями неодинаковы, что определяется, с одной стороны, вероятностными процессами, а с другой – определенной степенью неоднородности среды (характерно для растений и многих животных). Агрегированный (мозаичный) тип распределения выражается в образовании группировок особей, между которыми остаются достаточно большие незаселенные территории. Биологически это связано либо с резкой неоднородностью среды, либо с выраженной социальной структурой, действующей на основе активного сближения особей (особенно характерно для высших животных). Широко распространено групповое (контагиозное) распределение элементов в популяциях высших растений.
Пространственная структура популяции представляет собой не только статическую систему закономерного размещения особей по территории, но одновременно и динамическую систему взаимоотношений, выражающуюся в многообразных и упорядоченных формах поведения каждой особи в ответ на информацию о месте нахождения и поведения других особей.
При всем многообразии конкретных форм пространственной структуры популяций различных видов можно выделить два принципа ее построения, отличающихся способом использования территории.
Видам, для которых характерен оседлый образ жизни, как правило, свойственен интенсивный тип использования пространства, при котором отдельные особи или их группировки (главным образом семейные) в течение длительного времени используют ресурсы относительно ограниченной территории, к которой они весьма привязаны. Для видов, отличающихся кочевым (номадным) образом жизни, характерен экстенсивный тип использования территории, при котором кормовые ресурсы местности используются обычно группами особей (в том числе весьма многочисленными), постоянно перемещающимися в пределах обширной территории.
Привязанность к ограниченной территории имеет определенные биологические преимущества. Большое значение, в частности имеет знакомство с территорией. В своих перемещения по знакомому участку животное не затрачивает дополнительного времени и энергии на исследовательскую деятельность; все его передвижения осуществляются как бы автоматически, на основе системы знакомых зрительных ориентиров, запаховых и иных меток.
При всех преимуществах оседлый образ жизни имеет и определенные «теневые» стороны. Интенсивное использование ресурсов среды в ограниченном пространстве всегда таит в себе опасность того, что скопление нескольких особей приведет к быстрому истощению запасов корма и нехватке укрытий на территории, охватываемой их передвижениями. Это обстоятельство устанавливает определенные «лимиты» в отношении разнообразия форм пространственной структуры популяций у видов подобного экологического типа. Поэтому у всех видов, характеризующихся оседлым образом жизни и интенсивным использованием территории, пространственная структура популяций практически однотипна: она представлена системой индивидуальных участков обитания, в пределах которых каждая особь находит все необходимое для жизни и осуществляет все формы своей деятельности. Благодаря такому «распределению» участков между отдельными особями достигается более равномерное размещение их в пространстве, в результате чего каждое отдельное животное, не встречая на своем участке конкуренции со стороны других особей, имеет большие шансы на выживание и выращивание потомства, а популяция в целом получает более широкие перспективы роста и захвата территории.
Величина и форма участков обитания чрезвычайно сильно варьирует у различных видов животных и даже у разных популяций одного вида. Размеры участков определяются, по-видимому, сложным комплексом факторов, включающим как внешние (особенности среды), так и внутренние (особенности биологии данного вида).
Хорошо известно, что участки обитания, как правило, обеспечены кормовыми объектами в количестве, превышающем потребность их обитателей. Участок обитания не может превышать свойственной данному виду дистанции устойчивого общения, поскольку утрата информативных связей между особями означает распад популяции.
Предполагается, что расположение естественных убежищ (или мест пригодных, для постройки нор или гнезд) определяет места формирования «центров активности» участков обитания оседлых видов, обилие и дисперсность кормовых объектов – минимальные размеры этих участков, а условия коммуникации – их максимальные размеры; конкретные структурные особенности биотопа корректируют эти параметры.
Преимущества «владения» определенной территорией в виде участка обитания еще не гарантируются механизмами индивидуальной привязанности животных к этой территории. Для популяции должны существовать и специфические популяционные механизмы, определяющие необходимую степень диффузности в распределении особей и предотвращающие возможность чрезмерного скучивания и обострения внутривидовой конкуренции. В общем виде такие механизмы можно назвать формами «индивидуализации» территории и проявляются они в виде закономерных взаимоотношений между отдельными особями, входящими в состав популяции.
«Простейшая» форма таких взаимоотношений выражается в активной защите индивидуальной территории и физическом изгнании с нее других особей своего вида. Агрессивное поведение, связанное с защитой участка, известно во многих систематических группах животных. Наиболее интересным с биологической точки зрения является то, что победителем в территориальных схватках выходит не обязательно более крупный и сильный из партнеров, а, как правило, особь, на участке которой произошла встреча («хозяин» данного участка).
Маркировочное поведение и проявление прямых агрессивных реакций очень тесно связаны с физиологическим состоянием данной особи, в первую очередь – с развитием генеративной системы и способностью к размножению. Функционально активное состояние половой системы служит мощной базой проявления всего комплекса территориального поведения. Это подтверждается как полевыми наблюдениями, так и специальными экспериментами. Биологическое значение этого явления можно видеть в том, что именно в период размножения животные наиболее жестко привязаны к определенной точке территории, в этом случае особенно важна индивидуализация территории, как гарантия достаточности кормовых ресурсов и большей вероятности выживания взрослых особей и их потомства.
Формы агрессивного территориального поведения у большинства видов проявляются по-разному в зависимости от того, какое место в составе популяции занимает особь, проникшая на защищаемый участок. Как правило, активная защита участка наиболее выражена по отношению к «равному» партнеру, потенциальному конкуренту по различным линиям внутрипопуляционных отношений.
Неодинаковы формы проявления территориального поведения между особями различного пола и возраста. Анализируя степень выраженности, стресс реакции у внутрипопуляционных мигрантов рыжих полевок, отмечалось, что она высока у взрослых мигрантов, но подчас совсем не выражена у молодых расселяющихся видов. Это довольно определенно говорит о том, что молодые животные не испытывают той агрессии со стороны оседлых особей, с которой постоянно сталкиваются взрослые мигранты.
Показано, что у мышевидных грызунов состав веществ, определяющих продуцируемый железами запах, изменяется с возрастом: молодые животные обоих полов продуцируют запах, подобный таковому у взрослых самок; по достижении половозрелости у самцов запах усложняется и приобретает специфический характер. Видимо, запаховым сходством с самками и объясняется терпимое отношение взрослых самцов к неполовозрелым животным.
Групповой образ жизни, при котором животные постоянно или периодически образуют плотные стада или стаи, имеет свои биологические преимущества: более легкое нахождение и добывание корма, раннее обнаружение хищников, возможность коллективной обороны, более экономный тип энергозатрат и др. Вместе с тем скопления особей одного вида всегда сопряжены с повышением конкуренции. Один из путей преодоления этого противоречия — переход к кочующему образу жизни, связанному с экстенсивным использованием ресурсов. В популяциях стадных животных отсутствуют индивидуальные участки обитания. Элементарная единица популяции — стадо (стая), имеющее собственную структуру как в виде закономерного расположения особей в пространстве, так и в форме определенных взаимоотношений, обеспечивающих сохранение целостности группы при постоянных ее перемещениях. Исследования, проведенные на рыбах и птицах, показали, что пространственная структура ("строй") стад (стай) обеспечивает оптимальные гидро-(аэро-)динамические условия передвижения при сохранении возможности маневра как для группы в целом, так н для отдельных особей. Поддержание такой структуры определяется рядом специфических адаптаций, в основе которых лежит врожденное стремление находиться в контакте с соседними особями, т.е. интеграционное поведение, о котором говорилось выше. У стадных форм этот вид поведения, в частности, представлен стереотипом реакций слежения и подражания. На этих же формах поведения основывается синхронизация деятельности особей в группе, без чего невозможно поддержание целостности подвижной группировки.
Структура взаимоотношений особей в стадах и стаях отличается степенью сложности у разных видов. В простейшем случае подвижные группировки животных образуются исключительно на основе стремления к контактам и поддерживаются ориентацией на соседних особей и подражанием им (стада "эквипотенциального типа"). Особи в группе однородны по своим качествам и поведению. Такие группировки характерны для большинства стайных рыб, многих мелких птиц, мигрирующих стай саранчи и др.
Для многих копытных и некоторых других животных характерна более сложная структура стада: в ней выделяются лидеры, которые своим поведением определяют характер деятельности всего стада. Лидерами чаще становятся взрослые животные с богатым индивидуальным опытом. Именно на этих особях концентрируется подражательная реакция других членов стада; через подражание опыт лидирующих животных становится достоянием всего стада, что имеет несомненно адаптивное значение на уровне популяции в целом.
Наиболее сложна структура взаимоотношений в стадах, построенных по доминантно-иерархическому типу во главе с вожаками. Такие стада наиболее свойственны многим приматам, но встречаются и у некоторых хищных и копытных млекопитающих. В отличие от лидеров вожаки активно направляют деятельность стада путем специальной сигнализации, а подчас и с помощью агрессивных действий.
Помимо выделения вожака-доминанта, стада такого типа отличаются сложной внутренней структурой, весьма различной у отдельных видов — от деспотического доминирования единственного самца-вожака (лангуры) до наличия многих внутристадных группировок со своими формами отношений и отдельных рядов иерархии для самцов, самок и молодых животных у других видов обезьян.
Тенденция к усложнению структуры взаимоотношений повышает степень организованности и управляемости группы, что открывает возможность совмещения биологических выгод группового образа жизни с преимуществами обитания на хорошо освоенной территории.
39.Типы флуктуаций численности по амплитуде колебаний периодичности. Периодические и непериодические флуктуации.
При благоприятных условиях в популяциях наблюдается рост численности и может быть столь стремительным, что приводит к популяционному взрыву. Совокупность всех факторов способствующих росту численности называется биотическим потенциалом. Он достаточно высок для разных видов, но вероятность достижения популяцией предела численности в естественных условиях низка, т.к этому противостоят лимитирующие (ограничивающие) факторы. Совокупность факторов, лимитирующих рост численности популяции, называют сопротивлением среды. Состояние равновесия между биотическим потенциалом вида и сопротивлением среды поддерживающее постоянство численности популяции получило название гомеостаза или динамического равновесия. При нарушении его происходят колебания численности популяции, т.е. ее изменения.
Различают периодические и непериодические колебания численности популяций. Первые совершаются в течение сезона или нескольких лет (4 года - периодический цикл плодоношения кедра, подъема численности лемминга, песца, полярной совы; через год плодоносят яблони на садовых участках), вторые - это вспышки массового размножения некоторых вредителей полезных растений, при нарушениях условий среды обитания (засухи, необычно холодные или теплые зимы, слишком дождливые сезоны вегетации), непредвиденные миграции в новые местообитания. Периодические и непериодические колебания численности популяций под влиянием биотических и абиотических факторов среды, свойственные всем популяциям, именуются популяционными волнами.
Любая популяция обладает строго определенной структурой: генетической, половозрастной, пространственной и др., но она не может состоять из меньшего числа особей, чем необходимо для стабильного развития и устойчивости популяции к факторам внешней среды. В этом заключается принцип минимального размера популяций. Нежелательны любые отклонения параметров популяций от оптимальных, но если чрезмерно высокие значения их не представляют прямой опасности для существования вида, то снижение до минимального уровня, особенно численности популяции, представляют угрозу для вида.
Однако наряду с принципом минимального размера популяций есть и принцип (правило) популяционного максимума. Оно заключается в том, что популяция не может увеличиваться бесконечно. Лишь теоретически она способна к неограниченному росту численности.
Согласно теории Х.Г. Андреварты - Л.К. Бирча (1954) - теория лимитов популяционной численности, - численность естественных популяций ограничена истощением пищевых ресурсов и условий размножения, недоступностью этих ресурсов, слишком коротким периодом ускорения роста популяции. Теория "лимитов" дополняется теорией биоценотической регуляции численности популяции К. Фредерикса (1927): рост численности популяции ограничивается воздействием комплекса абиотических и биотических факторов среды.
Факторы или причины колебания численности:
достаточные запасы пищи и ее недостаток;
конкуренция нескольких популяций из-за одной экологической ниши;
взаимоотношения между популяциями хищника и жертвы, хозяина и паразита;
внешние (абиотические) условия среды: гидротермический режим, освещенность, кислотность, аэрация и др.
Кроме экологических факторов, включаются внутренние (генетические и физиологические) механизмы регулирования численности популяций: при сокращении жизненного пространства и запасов корма сокращается плодовитость особей (многие насекомые, мышевидные грызуны), повышение смертности на ранних стадиях жизни (паразиты, многие насекомые), задерживается наступление половой зрелости (полевые мыши) и стадии плодоношения (виды деревьев 2 и 3 ярусов в густом лесу), имеет место каннибализм (грызуны, насекомые, рыбы), и др.; снижается выход личинок из яиц (майский хрущак), уменьшаются размеры взрослых особей. При чрезмерном росте численности популяции у млекопитающих, общественных насекомых, птиц начинается эмиграция на новые места.
Флуктуации (отклонения) численности вызываются самыми различными причинами. И они не всегда одинаковые для разных видов. Периодические колебания численности популяций, имеющих 10-11-летний период, объясняются периодичностью активности Солнца: количество пятен на Солнце меняется с периодом в 11 лет. Количество корма есть причина флуктуации у сибирского шелкопряда: он дает вспышку после сухого теплого лета. Может вызвать вспышку численности и стечение многих обстоятельств. Например, у берегов Флориды наблюдаются "красные приливы". Они непериодичны и для их проявления необходимы такие события: обильные ливни, смывающие с суши микроэлементы (железо, цинк, кобальт - их концентрация должна совпадать до десятитысячной доли процента), пониженная соленость поды, определенная температура и безветрие у берега. При таких условиях водоросли динофлагелляты начинают интенсивно делиться. Теоретически из одной одноклеточной динофлагелляты в результате 25 последовательных делений могут произойти 33 млн. особей. Вода от них становится красной. Динофлагелляты выделяют в воду смертоносный яд, вызывающий паралич, а затем гибель рыбы и других обитателей моря.
Человек может своей деятельностью вызвать вспышку численности некоторых популяций. Результатом антропического воздействия есть возрастание численности сосущих насекомых (тлей, клопов и др.) после обработки полей инсектицидами, которые уничтожают их врагов. Благодаря человеку кролики и кактус опунция в Австралии, домовые воробьи и непарный шелкопряд в Северной Америке, колорадский жук и филлоксера в Европе, канадская элодея, американская норка и ондатра в Евразии дали неимоверные вспышки численности после попадания на эти новые для них территории, где не было их врагов.
Резкие непериодические колебания численности могут возникать вследствие природных катастроф. Например, на пожарищах обычны вспышки численности иван-чая и связанного с ним сообщества насекомых. Многолетняя засуха превращает болото в луг и вызывает рост численности членов биоценоза луга.
"Волны жизни" исчезают за счет появления большого количества хищников, паразитов, возникновения эпизоотии, изменения абиотических условий.
Эволюционное значение популяционных волн в том, что они:
изменяют частоты аллелей (малочисленные на пике волны могут проявиться фенотипически, а на спаде - исчезнуть из генофонда);
на пике волны изолированные популяции сливаются, растет миграция и панмиксия, растет гетерогенность генофонда;
популяционные волны изменяют интенсивность природного отбора и его направление.
40.Факторы регуляции численности популяции.