РК № 2-1. 1 Классификация углеводов
 Скачать 1.85 Mb. Скачать 1.85 Mb.
|

-1-Классификация углеводовМоносахариды, которые не могут быть гидролизованы на более простые сахара. В зависимости от числа атомов углерода их подразделяют на триозы, тетрозы, пентозы, гексозы. В зависимости от присутствия альдегидной или кетоновой группы на альдозы и кетозы. Дисахариды состоят из двух остатков моносахаридов: 1) сахароза состоит из остатков глюкозы и фруктозы, соединенных a-1,4-гликозидной связью; 2) лактоза состоит из остатков глюкозы и галактозы, соединенных b-1,4-гликозидной связью; 3) мальтоза состоит из двух остатков глюкозы, соединенных a-1,4-гликозидной связью; 4) целлобиоза состоит из двух остатков глюкозы, соединенных b-1,4-гликозидной связью. Гомополисахариды - длинные разветвленные цепи, состоящие из одних и тех же моносахаридов: 1) крахмал - полимер глюкозы, соединенной a-1,4 и a-1,6-гликозидными связями. При этом неразветвленные цепи образуют амилозу (20%), а разветвленные амилопектин (80%); 2) гликоген - животный крахмал, состоящий из остатков глюкозы. Это более разветвленный полимер, чем крахмал. При частичном гидролизе крахмала или гликогена образуются декстрины (более короткие разветвленные цепи); 3) целлюлоза - главный компонент структурной основы растительных клеток. Это линейный полимер глюкозы, соединенной b-1,4-гликозидными связями. Гетерополисахариды состоят из разных мономеров: 1) гепарин содержит остатки D-глюконат-2-сульфита и N-ацетилглюкозамин-6-сульфата; 2) гиалуроновая кислота состоит из остатков D-глюкуроновой кислоты и остатков N-ацетилглюкозамина. Входит в состав соединительной ткани и участвует в регуляции проницаемости кканей. Суточная норма углеводов в пище составляет 400-500 г. -2- Переваривание и всасывание углеводов Целлюлоза не расщепляется в желудочно-кишечном тракте, у человека соответствующие ферменты не вырабатываются. Однако непереваренная целлюлоза из растительной пищи способствует нормальной перистальтике кишечника. Гидролиз крахмала начинается в полости рта при участии амилазы слюны, которая частично расщепляет внутренние -1,4-гликозидные связи, образуя менее крупные, чем крахмал молекулы – декстрины. Потенциально -амилаза слюны в ротовой полости способна расщепить пищевой крахмал или гликоген до дисахаридов мальтозы и изомальтозы. Переваривание углеводов возобновляется при поступлении пищевых масс из желудка в тонкий кишечник. Поступающий оттуда кислый химус нейтрализуется щелочными солями (бикарбонатами), поступающими в 12-перстную кишку вместе с соком поджелудочной железы. К тому же, в стенке этой кишки есть железы, тоже вырабатываюшие бикарбонаты. Панкреатическая -амилаза завершает расщепление полисахаридов и олигосахаридов до дисахарида мальтозы.Дисахарид мальтоза и остальные дисахариды, поступившие с пищей расщепляются ферментами пристеночного переваривания углеводов до моносахаридов (сахаразой, лактазой, мальтазой и изомальтазой) Эти ферменты выделяются слизистой оболочкой кишечника в составе кишечного сока. После всасывания глюкоза по системе воротной вены поступает в печень. В печени основное количество глюкозы откладывается запасается в виде гликогена, а остальная глюкоза идёт в общий кровоток для питания других клеток. Так происходит после принятия пищи на высоте пищеварения.В состоянии "натощак" (вне приёма пищи) гликоген в печени постепенно распадается до глюкозы, и глюкоза из печени уходит в общий кровоток к другим тканям. -3- Транспорт моносахаридов из просвета кишечника в клетки слизистой оболочки может осуществляться путем облегченной диффузии и активного транспорта. При активном транспорте глюкоза и Na+ проходят с люминальной стороны, связываясь с разными участками белка-переносчика. При этом Na+ поступает в клетку под влиянием электрохимического градиента и «тащит» глюкозу за собой. чем больше градиент Na+, тем больше поступление глюкозы. Если концентрация Na+ во внеклеточной жидкости уменьшается, транспорт глюкозы подавляется. Градиент концентрации Na+ являющийся движущей силой этого симпорта, создается работой Na+K+ нacoca.Глюкоза из клетки кишечника затем перемещается во внеклеточную жидкость и далее в кровь с помощью облегченной диффузии. Поступающая из кишечника глюкоза с кровью воротной вены попадает в печень, где часть ее задерживается, а часть через общий кровоток попадает в клетки других органов и тканей.Потребление глюкозы клетками из кровотока происходит также путем облегченной диффузии при участии специальных белков-транспортеров. Следовательно, скорость трансмембранного потока глюкозы зависит только от градиента ее концентрации. Исключением являются клетки мышц и жировой ткани, где облегченная диффузия регулируется инсулином. Белки-переносчики (транспортеры глюкозы — ГЛЮТ) обнаружены во всех тканях. Существует несколько разновидностей ГЛЮТ, которые пронумерованы по порядку их обнаружения. Все 5 типов ГЛЮТ имеют сходную первичную структуру. ГЛЮТ-1 служит для обеспечения стабильного потока глюкозы в мозг. В других тканях он поставляет глюкозу в клетки, когда они находятся в стоянии покоя. ГЛЮТ-2 обнаружен в клетках органов, выделяющих глюкозу в кровь. Именно при участии ГЛЮТ-2 глюкоза переходит в кровь из энтероцитов после всасывания в кишечнике. ГЛЮТ-3 обладает большим, чем ГЛЮТ-1, сродством к глюкозе. Он также обеспечивает постоянный приток глюкозы к клеткам нервной ткани. ГЛЮТ-4 — главный переносчик глюкозы в мышцах и адипоцитах. ГЛЮТ-5 встречается главным образом в клоках тонкой кишки. Его функции известны недостаточно. В клетки печени глюкоза проходит при участии ГЛЮТ-2, независимого от инсулина. Транспорт глюкозы из первичной мочи в клетки канальцев происходит путем активного транспорта. -4- Гликозаминогликаны (ГАГ) – углеводная часть углеводсодержащих биополимеров гликозамино-протеогликанов или протеогликанов. Прежнее название гликозаминопротеогликанов "мукополисахариды" исключено из химической номенклатуры. ГАГ входят в состав сложных белков, которые называются ПРОТЕОГЛИКАНАМИ. ГАГ составляют в протеогликанах 95% их веса. Остальные 5% веса - это белок. Белковый и небелковый компоненты в протеогликанах связаны прочными, ковалентными связями. На рисунке 15. представлено строение молекулы протеогликанов. 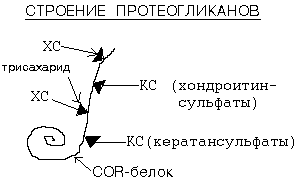 Рисунок 15. Белковый компонент - это особый COR-белок. К нему при помощи трисахаридов присоединяются ГАГ. 1 молекула COR-белка может присоединить до 100 ГАГ.В клетке протеогликаны связаны с гиалуроновой кислотой. Образуется сложный надмолекулярный комплекс. Упругие цепи ГАГ в составе протеогликанов образуют образуют макромолекулярные сетчатые структуры. Такое химическое строение обеспечивает выполнение функции молекулярного сита с определенными размерами пор при транспорте различных веществ и метаболитов. Размер пор определяется типом ГАГ, преобладающим в данной конкретной ткани. Например, соединительнотканная капсула почечного клубочка обеспечивает селективный транспорт веществ в процессе ультрафильтрации. За счет множества сульфо- и карбоксильных групп сетчатые структуры являются полианионами, способными депонировать воду, некоторые катионы (К+, Na+, Ca+2, Mg+2).Состав и функции гликопротеидов. Общая характеристика функции гаптоглобина, трансферрина, церулоплазмина, фибриногена, - фетопротеина. Углеводсодержащие белки — гликопротеины — представляют собой комплексы белков, гексоз и их производных. Они вырабатываются и секретируются клетками печени под влиянием катехоламинов. Наиболее известными представителями собственно гликопротеинов являются различные ферменты (например, церулоплазмин), фибриноген, гаптоглобин, трансферрин, фибронектин, ламинин и т.д. Напр: Гаптоглобин – синтезируемый в печени белок. Его характерной особенностью является способность связываться с гемоглобином с образованием комплекса («гемоглобин–гаптоглобин»), не проходящего через почечный барьер. Тем самым в организме задерживается очень ценный для него элемент – железо, а почки предохраняются от формирования гемосидероза. -5- Гликолиз Процесс распада глюкозы до лактата в анаэробных условиях называется ГЛИКОЛИЗОМ,В состоянии покоя, наступающего после интенсивной мыщечной работы, в клетку начинает поступать кислород. Это приводит к запуску митохондриальных дыхательных цепей. Запускаются окислительное декарбоксилирование пирувата, ЦТК и челночный механизм переноса водорода в митохондрии, а, значит, и ГБФ-путь распада глюкозы (гликогена). При этом процесс гликолиза тормозится автоматически. Торможение гликолиза поступившим в клетку кислородом называется ЭФФЕКТОМ ПАСТЕРА по имени ученого, открывшего это явление.Еще существует аэробный гликолиз: Аэробным гликолизом называют процесс окисления глюкозы до пировиноградной кислоты, протекающей в присутствии кислорода. Все ферменты, катализирующие реакции этого процесса, локализованы в цитозоле клетки. БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ГБФ-ПУТИ. 1. Это главный путь распада углеводов до конечных продуктов. Во многих клетках это - единственный путь. Так распадается 70-75% глюкозы, которая поступает в клетку. 2. Только ГБФ-путь дает клетке энергию в виде АТФ. Это основной источник получения энергии в клетке. 3. Это самый длинный путь распада углеводов. Этапы гликолеза: Глюкоза(фермент:гексокиназа) - глюкозо-6-фосфат Глюкозо-6-фосфат(глюкозо-6-фофат изомераза)- фруктозо-6-фосфат фруктозо-6-фосфат(фосфофруктокиназа)-фруктозо1,6-биофосфат фруктозо1,6-биофосфат(альдолаза)-ФГА+ФДА Обратимая: ФДА(триозофосфатизомераза)-ФГА ФГА(ФГА ДГ)- 1,3 бифосфоглицерат 1,3 бифосфоглицерат(фосфоглицераткиназа)- 3-фосфоглицерат 3-фосфоглицерат(фосфоглицератмутаза)-2-фосфоглицерат 2-фосфоглицерат(енолаза)-фосфоенулпируват Фосфоенулпируват(пируваткиназа)-пируват Ключевые ферменты первого этапа ГБФ-пути: ГЕКСОКИНАЗА и ФОСФОФРУКТОКИНАЗА. -6- Гликолитическая оксидоредукция |