|
|
1. Клеочная теория, этапы развития значения для биологии
28. Уникальные и повторяющиеся последовательности ДНК.
Более половины геномной ДНК эукариотических организмов принадлежит к классу уникальных, или неповторяющихся, последовательностей. Это утверждение (а также оценки, касающиеся распределения в геноме повторяющихся последовательностей ДНК) базируется на данных непрямых экспериментов с применением различных методик ДНК-РНК-гибридизации, позволяющих получить лишь приблизительную оценку. У дрожжей — низших эукариот — экспрессируется около 4000 генов. В типичной ткани млекопитающих (печень или почки) экспрессируется от 10000 до 15000 генов. При этом в каждой ткани происходит экспрессия специфического набора генов. Каким образом это достигается, по-прежнему остается одним из центральных вопросов современной биологии.
Интроны
Кодирующие области ДНК, транскрипты которых попадают в цитоплазму в составе «зрелых» молекул мРНК, прерываются в геноме длинными последовательностями некодирующей ДНК. Соответственно первичные транскрипты ДНК содержат некодирующие промежуточные последовательности.
РНК, которые должны быть удалены в процессе созревания, обеспечивающего также и правильную стыковку (сплайсинг) кодирующих сегментов в зрелых мРНК. Большинство последовательностей, транскрипты которых представлены в зрелой мРНК, разорваны в геноме от одного до пятидесяти раз некодирующими вставками (нитронами). Как правило, интроны значительно длиннее, чем кодирующие участки (экзоны). Процессинг первичного транскрипта, включающий удаление интронов и сплайсинг соответствующих экзонов.
Функция интронов точно не установлена. Можно предположить, что они служат для физического разделения экзонов, соответствующих функциональным доменам кодируемых белков, с целью оптимизации процесса генетических перестроек (рекомбинаций), которые могут происходить с более высокой эффективностью при наличии интронов, чем в случае сосредоточения генетической информации в одном континууме. Увеличение темпа генетических перестроек функциональных доменов может рассматриваться как фактор ускорения эволюции биологических функций.
Повторяющиеся последовательности ДНК
Под повторяющимися последовательностями ДНК (повторами) понимается широкий спектр как умеренно повторяющихся, так и часто повторяющихся (высокоповторяющихся) последовательностей ДНК. По крайней мере 20—30% генома человека представлено повторами.
Высокоповторяющиеся последовательности состоят из участков длиной в 5—500 пар оснований, повторяющихся много раз и расположенных один за другим (тандемно). Эти последовательности обычно образуют кластеры и присутствуют в количестве от 1 до 10 миллионов копий на гаплоидный геном. Высокоповторяющиеся последовательности транскрипционно-неактивны и, вероятно, участвуют в структурировании хроматина.
Умеренно повторяющиеся последовательности, присутствующие в количестве менее чем 106 копий на гаплоидный геном, не образуют кластеров, а чередуются с неповторяющимися (уникальными) последовательностями. Они могут быть как короткими, так и весьма протяженными. Длинные диспергированные повторы состоят из 5000—7000 пар оснований и представлены в количестве 1000—100000 копий на гаплоидный геном. Они фланкированы с обоих концов прямыми повторами длиной в 300— 600 пар оснований (рис. 38.8). Во многих случаях длинные повторы транскрибируются РНК-полимеразой II в виде молекул мРНК, содержащих такие же кэпированные 5-концы, как и мРНК.
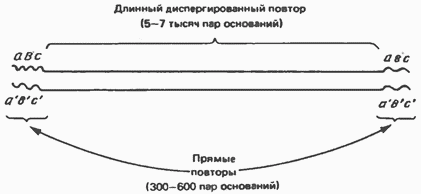
Схема длинного диспергированного повтора. Отмечено расположение на концах повтора коротких прямых повторов (abc) и соответствующих комплементарных участков (аbс).
Короткие диспергированные повторы представляют семейства родственных, но отличных друг отдруга фрагментов длиной от единиц (нескольких пар) до нескольких сотен пар нуклеотидов. Короткие повторы активно транскрибируются либо как компоненты интронов, либо под контролем ДНК-зависимой РНК-полимеразы III как самостоятельные элементы . Наиболее многочисленным семейством коротких диспергированных повторов в геноме человека является семейство Alu, насчитывающее около 500000 копий на гаплоидный геном, что составляет 3—6% от общего размера генома человека. Повторы этого семейства (а также их аналоги у животных) транскрибируются и в составе и в виде дискретных молекул РНК, включая хорошо изученные . Такого типа последовательности высококонсервативны как внутри данного вида, так и у разных видов млекопитающих. По своей структуре короткие диспергированные повторы, в том числе члены семейства Alu, напоминают длинные концевые повторы ретровирусов (LTR). По-видимому, это мобильные элементы, способные как встраиваться, так и вырезаться из различных участков генома.
29.белки хромосом гистоны, негистоновые белки; их роль в хроматине и хромосомах.
Хромосомная ДНК упакована в компактную структуру с помощью специализированных белков. Все ДНК-связывающие белки эукариот подразделяются на два класса: на гистоны (histones, структурные белки эукариотических хромосом) и на негистоновые хромосомные белки . Комплекс обоих классов белков с ядерной ДНК эукариотических клеток называется хроматином . Гистоны являются уникальной характеристикой эукариот и присутствуют в огромных количествах на клетку (около 60 миллионов молекул каждого типа на клетку).Гистоны - относительно небольшие белки с очень большой долей положительно заряженных аминокислот (лизина и аргинина); положительный заряд помогает гистонам крепко связываться с ДНК (которая заряжена сильно отрицательно) независимо от ее нуклеотидной последовательности. Возможно, гистоны только изредка диссоциируют от ДНК и таким образом, вероятно, оказывают влияние на любой процесс, происходящий на хромосомах.Типы гистонов распадаются на две главных группы - нуклеосомные гистоны и Н1 гистоны, образуя семействовысококонсервативных основных белков, состоящее из пяти больших классов - H1 и H2A ,H2B , H3 и H4 . Гистоны H1 более крупные (около 220 аминокислот) и оказались менее консервативными в ходе эволюции. Размер полипептидных цепей гистонов лежит в пределах от 220 (H1) до 102 (H4) аминокислотных остатков. Гистон H1 сильно обогащен остатками Lys, для гистонов H2A и H2B характерно умеренное содержание Lys, полипептидные цепи гистонов H3 и H4 богаты Arg. Внутри каждого класса гистонов (за исключением H4) на основании аминокислотных последовательностей различают несколько субтипов этих белков. Такая множественность особенно характерна для гистонов класса H1 млекопитающих. В этом случае различают семь субтипов, названных H1.1-H1.5 , H1o и H1t.Гистоны H3 и Н4 принадлежат к наиболее консервативным белкам. Такая эволюционная консервативность предполагает, что для функции данных гистонов важны почти что все их аминокислоты. N- концевая часть данных гистонов может быть обратимо одифицирована в клетке за счет ацетилирования отдельных остатков лизина, что убирает положительный заряд лизинов.Гистоны подвергаются многообразным посттрансляционным модификациям , которые вовлечены в разнообразные биологические процессы, например, регуляцию активности генов , репарацию ДНК ,конденсацию хроматина.
30. Виды РНК, их функции и образование в связи с активностью хроматина. Центральная догма клеточной биологии: ДНК-РНК-белок. Роль компонентов в ее реализации.
Центральная догма молекулярной биологии — обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении. Правило было сформулировано Френсисом Криком в 1958 году и приведено в соответствие с накопившимися к тому времени данными в 1970 году. Переход генетической информации от ДНК к РНК и от РНК к белку является универсальным для всех без исключения клеточных организмов, лежит в основе биосинтеза макромолекул. Репликации генома соответствует информационный переход ДНК → ДНК. В природе встречаются также переходы РНК → РНК и РНК → ДНК (например у некоторых вирусов), а также изменение конформации белков, передаваемое от молекулы к молекуле. Транскрипция и трансляция. Условно весь процесс транскрипции и трансляции можно отобразить в cхеме: Транскрипция представляет собой процесс воспроизведения информации, хранящейся в ДНК, в виде одноцепочной молекуле и РНК (информационной РНК, которая переносит информацию о строении белка из ядра клетки в цитоплазму клетки к рибосомам). Этот процесс проявляется в синтезе молекулы и РНК по матрице ДНК. Молекула и РНК состоит и нуклеотидов, каждый из которых включает в себя остаток фосфорной кислоты сахар рибозу и одно из четырёх азотистых оснований (А, Г, Ц и У-урацил вместо Т-тюлина). В основе синтеза и РНК лежит принцип комплиментарности, т.е. против А в одной цепочке ДНК располагается У в и РНК, а против Г в ДНК - Ц в и РНК (см. рис. Транскрипция- на предыдущей странице), таким образом, и РНК является комплиментарной копией ДНК или её определённого участка, и содержит информацию, кодирующую аминокислоту или белок. Каждая аминокислота в ДНК и РНК шифруется последовательностью из 3-х нуклеотидов, т.е. - триплетом, который получил название кодонЕсли в транскрипции узнавание двух молекул друг другом проявляется только в принципе комплиментарности, то в трансляции помимо комплиментарности (временное объединение кодона и РНК и антикодона РНК (транспортной РНК, которая подносит аминокислоты нужные для синтеза белка, к месту синтеза - рибосома - см. рис. Транскрипция) молекулярное узнавание проявляется в процессе присоединения аминокислоты к тРНК с помощью фермента кодазы. Дело в том, что молекула тРНК состоит из головки, включающей в себя антиэАОК-триплет, состоящий из последовательности трёх нуклеотидов, и хвостика имеющего определённую форму. Сколько существует видов антикозонов тРНК, столько и существует форм хвостиков, и каждому антикозону соответствует своя форма хвостика в тРНК. Сколько существует форм хвостиков, столько существует видов форм фермента кодазы, который присоединяет аминокислоты к хвостику, а форма каждой кодазы подходит только к форме определённой аминокислоты. Т.о., тРНК носит с собой информацию не только в п последовательности нуклеотидов в антикозоне но и в форме хвостика молекулы. А основная передача информации здесь заключается в воспроизведении последовательности аминокислот в белке, которую подсказывает ферменту, кодирующему белок и РНК.
31. Уровни упаковки ДНК в составе хроматина.
Хромосомы состоят из хроматина — смеси ДНК и белков в соотношении
1:1. Именно благодаря взаимодействию с белками молекула ДНК имеет компактную структуру. Средняя длинна одной нити ДНК человека (если растянуть) 4 см. То есть в ядре каждой клетки содержится около 1 метра ДНК.
Упаковка молекулы ДНК в хромосоме имеет три уровня:
1. нуклеосомы, нуклеосомная нить
2. соленоидная спираль
3. петли
Известная каждому своим видом двойная спираль ДНК «намотанная» вокруг комплекса из 4-х пар белковых молекул образует «бусину» - нуклеосому. Нуклеосомы, связанные между собой участками молекулы ДНК, составляют нить, которая в свою очередь закручена в форме соленоида (это по одной из существующих моделей), один виток — 6 нуклеосом. Эта структура на следующем уровне упаковки образует петли. В таком виде ДНК присутствует в ядре клетки на этапе роста клетки, синтеза её компонентов, в т. ч. удвоения ДНК.
Однако на разных стадиях жизни клетки конденсация хроматина разная. В период роста клетки хроматин в ядре не конденсирован, компактность его расположения при этом отличается для разных участков хромосомы.
Когда же происходит процесс деления клетки хроматин всё более уплотняется и петли, о которых говорилось выше, образуют «розетки», плотно уложенные одна к другой (тоже по одной из существующих моделей), по 18 петель на кольцо. В результате этого хроматин принимает вид конденсированных хромосом, так хорошо известный нам из учебников.
32. Митотические хромосомы. Морфологическая организация и функции. Кариотип ( на примере человека).
Митотические хромосомы образуются в клетке во время митоза. Это неработающие хромосомы, и молекулы ДНК в них уложены чрезвычайно плотно. Достаточно сказать, что общая длина метафазных хромосом примерно в 104 раз меньше, чем длина всей ДНК, содержащейся в ядре. Благодаря такой компактности митотических хромосом обеспечивается равномерное распределение генетического материала между дочерними клетками при митозе.
Кариоти́п — совокупность признаков (число, размеры, форма и т.д.) полного набора хромосом, присущий клеткам данного биологического вида (видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Кариотипом иногда также называют и визуальное представление полного хромосомного набора (кариограммы).
Определение кариотипа
Внешний вид хромосом существенно меняется в течение клеточного цикла: в течение интерфазы хромосомы локализованы в ядре, как правило, деспирализованы и труднодоступны для наблюдения, поэтому для определения кариотипа используются клетки в одной из стадий их деления — метафазе митоза.
Процедура определения кариотипа
Для процедуры определения кариотипа могут быть использованы любые популяции делящихся клеток, для определения человеческого кариотипа используется либо одноядерные лейкоциты, извлечённые из пробы крови, деление которых провоцируется добавлением митогенов, либо культуры клеток, интенсивно делящихся в норме (фибробласты кожи, клетки костного мозга). Обогащение популяции клеточной культуры производится остановкой деления клеток на стадии метафазы митоза добавлением колхицина — алкалоида, блокирующего образование микротрубочек и «растягивание» хромосом к полюсам деления клетки и препятствующего тем самым завершению митоза.
Полученные клетки в стадии метафазы фиксируются, окрашиваются и фотографируются под микроскопом; из набора получившихся фотографий формируются т. н.систематизированный кариотип — нумерованный набор пар гомологичных хромосом (аутосом), изображения хромосом при этом ориентируются вертикально короткими плечами вверх, их нумерация производится в порядке убывания размеров, пара половых хромосом помещается в конец набора (см. Рис. 1).
Исторически первые недетализованные кариотипы, позволявшие проводить классификацию по морфологии хромосом получались окраской по Романовскому — Гимзе, однако дальнейшая детализация структуры хромосом в кариотипах стала возможой с появлением методик дифференциального окрашивания хромосом.
Классический и спектральный кариотипы.
33. Репродукция хромосом про- и эукариот, взаимосвязь с клеточным циклом.
Обычно клеточный цикл у эукариот состоит из четырех временных отрезков: собственно митоза (M ), пресинтетической ( G1 ), синтетической ( S ) и постсинтетической ( G2 ) фаз (периодов). Известно, что общая продолжительность как всего клеточного цикла, так и отдельных его фаз значительно варьируют не только у разных организмов, но и у клеток разных тканей и органов одного организма.
Универсальная теория клеточного цикла предполагает, что клетка как целое в течение клеточного цикла проходит через ряд состояний ( Hartwell L., 1995 ). В каждом состоянии критические регуляторные белки претерпевают фосфорилирование или дефосфорилирование, определяющие переход этих белков в активное или неактивное состояние, их взаимосвязи и/или клеточную локализацию.
Изменения состояний клетки в определенных точках цикла организует особый класс протеинкиназ - циклинзависимые киназы (Cyclin-dependent kinases - cdk ). Cdk образуют комплексы со специфическими короткоживущими белками - циклинами , вызывающими их активацию, а также с другими вспомогательными белками.
Предполагается, что простейший клеточный цикл может состоять только из двух фаз - S и M, регулируемых соответствующими cdk. Такой гипотетический клеточный цикл имеет место во время раннего эмбриогенеза у организмов с большими яйцеклетками, например у Xenopus и Drosophila. В этих яйцеклетках все компоненты, необходимые для многочисленных делений, пресинтезированы во время оогенеза и сохраняются в цитоплазме. Поэтому после оплодотворения деления происходят чрезвычайно быстро, и периоды G1 и G2 отсутствуют.
Клеточная пролиферация контролируется сложной сетью внеклеточных и внутриклеточных событий, приводящих либо к инициации и поддержанию клеточного цикла, либо к выходу клеток в фазу покоя .
Центральным событием клеточного цикла является репликация ДНК.
Репликация ДНК требует присутствия достаточно большого набора ферментов и белковых факторов, упаковка вновь синтезированной ДНК в хроматин нуждается также в синтезе гистонов de novo. Экспрессия генов , кодирующих перечисленные белки, специфична для S-фазы.
После завершения репликации, когда генетический материал удвоен, клетка входит в постсинтетическую фазу G2 , во время которой происходит подготовка к митозу.В результате митоза ( М-фаза ) клетка разделяется на две дочерних. Обычно выделяют два критических перехода между фазами- G1/S и G2/M0.
Исходя из схемы клеточного цикла, можно заключить, что клетки останавливались бы в точке рестрикции R в фазе G1, если бы стадия G1 представляла собой биосинтетическую реакцию, намного более чувствительную к ингибированию общего синтеза белка, чем любые другие реакции, специфичные для отдельных фаз цикла.
Было высказано предположение, что для прохождения точки рестрикции R концентрация каких-то триггерных белков должна превысить некоторый пороговый уровень.
Согласно этой модели любые условия, снижающие общую интенсивность белкового синтеза , должны отдалять накопление пороговой концентрации триггерного белка, удлинять фазу G1 и замедлять темп деления клеток. Действительно, когда клетки растут in vitro в присутствии различных концентраций ингибиторов белкового синтеза, клеточный цикл сильно растягивается, тогда как время, необходимое для прохождения фаз S, G2 и М, существенно не изменяется. Наблюдаемое удлинение фазы G1 согласуется с данной моделью, если предположить, что каждая молекула триггерного белка остается активной в клетке лишь несколько часов. Эта модель позволяет также объяснить торможение роста клеток при увеличении их плотности или при голодании; как известно, оба этих фактора снижают синтез белка и останавливают клеточный цикл в наиболее чувствительной точке фазы G1- точке R.
По-видимому, механизмы, контролирующие рост клеток в ткани, воздействуют непосредственно на общую интенсивность белкового синтеза в клетках; согласно этой гипотезе, в отсутствии специфических стимулирующих факторов (и/или при наличии ингибирующих факторов) клетки будут синтезировать белки лишь на некотором базальном уровне, поддерживающем status quo. См белок RB: роль в регуляции клеточного цикла . При этом количество белков со средней скоростью обновления будет поддерживаться на том же уровне, что и в растущих клетках, а концентрация нестабильных белков (в т.ч. и триггерного белка будет уменьшаться пропорционально уменьшению скорости их синтеза. В условиях, способствующих ускорению общего белкового синтеза, количество триггерного белка превысит пороговый уровень, что позволит клеткам пройти точку рестрикции R и приступить к делению. |
|
|
 Скачать 217.54 Kb.
Скачать 217.54 Kb.