1. Клеочная теория, этапы развития значения для биологии
 Скачать 217.54 Kb. Скачать 217.54 Kb.
|

|
38.Клеточный цикл, периоды и фазы Клеточный цикл включает строго детерминированный ряд последовательных процессов, согласно позиции Hartwellа, 1995. Клетка должна между двумя последовательными делениями удвоить все свои компоненты и свою массу. клеточный цикл составляют два периода: 1) период клеточного роста, называемый " интерфаза ", и 2) период клеточного деления, называемый " фаза М " (от слова mitosis). В свою очередь, в каждом периоде выделяют несколько фаз. Обычно интерфаза занимает не меньше 90% времени всего клеточного цикла. Например, у быстро делящихся клеток высших эукариот последовательные деления происходят один раз в 16-24 часа, и каждая фаза М длится 1-2 часа. Большая часть компонентов клетки синтезируется на протяжении всей интерфазы, это затрудняет выделение в ней отдельных стадий по мнению Pardee, 1989. В интерфазе выделяют фазу G1, фазу S и фазу G2. Период интерфазы, когда происходит репликация ДНК клеточного ядра, был назван " фаза S " (от слова synthesis). Период между фазой М и началом фазы S обозначен как фаза G1 (от слова gap - промежуток), а период между концом фазы S и последующей фазой М - как фаза G2. Период клеточного деления (фаза М) включает две стадии: митоз (деление клеточного ядра) и цитокинез (деление цитоплазмы). В свою очередь, митоз делится на пять стадий, In vivo эти шесть стадий образуют динамическую последовательность. Описание клеточного деления базируется на данных световой микроскопии в сочетании с микрокиносъемкой и на результатах световой и электронной микроскопии фиксированных и окрашенных клеток. Повторяющаяся совокупность событий, обеспечивающих деление эукариотических клеток, получила название клеточного цикла. Продолжительность клеточного цикла зависит от типа делящихся клеток. Некоторые клетки, например, нейроны человека, после достижения стадии терминальной дифференцировки прекращают свое деление вообще. Клетки легких, почек или печени во взрослом организме начинают делиться лишь в ответ на повреждение соответствующих органов. Клетки эпителия кишечника делятся на протяжении всей жизни человека. Даже у быстро пролиферирующих клеток подготовка к делению занимает около 24 ч. Клеточный цикл разделяют на стадии: Митоз - М-фаза, деление клеточного ядра. G1 -фаза период перед синтезом ДНК. S-фаза - период синтеза (репликации ДНК). G2-фаза - период между синтезом ДНК и митозом. Интерфаза - период, включающий в себя G1 -, S- и G2-фазы. Цитокинез - деление цитоплазмы. Точка рестрикции, R-point - время в клеточном цикле, когда продвижение клетки к делению становится необратимым. G0 фаза - состояние клеток, достигших монослоя или лишенных фактора роста в ранней G1 фазе. Делению клетки (митозу или мейозу) предшествует удвоение хромосом, которое происходит в периоде S клеточного цикла (рис.1). Период обозначают первой буквой слова synthesis - синтез ДНК. С момента окончания периода S до завершения метафазы ядро содержит в четыре раза больше ДНК, чем ядро сперматозоида или яйцеклетки, а каждая хромосома состоит из двух идентичных сестринских хроматид. Во время митоза хромосомы конденсируются и в конце профазы или начале метафазы становятся различимыми при оптической микроскопии. Для цитогенетического анализа обычно используют препараты именно метафазных хромосом. В начале анафазы центромеры гомологичных хромосом разъединяются, и хроматиды расходятся к противоположным полюсам митотического веретена. После того как к полюсам отойдут полные наборы хроматид (с этого момента их называют хромосомами), вокруг каждого из них образуется ядерная оболочка, формируя ядра двух дочерних клеток (разрушение ядерной оболочки материнской клетки произошло в конце профазы). Дочерние клетки вступают в период G1, и только при подготовке к следующему делению они переходят в период S и в них происходит репликация ДНК. Клетки со специализированными функциями, длительное время не вступающие в митоз или вообще утратившие способность к делению, находятся в состоянии, называемом периодом G0. Большинство клеток в организме диплоидные - то есть имеют два гаплоидных набора хромосом (гаплоидный набор - это число хромосом в гаметах, у человека он составляет 23 хромосомы, а диплоидный набор хромосом - 46). В гонадах предшественники половых клеток сначала претерпевают ряд митотических делений, а затем вступают в мейоз - процесс образования гамет, состоящий из двух последовательных делений. В мейозе гомологичные хромосомы спариваются (отцовская 1-я хромосома с материнской 1-й хромосомой и т. д.), после чего в ходе так называемого кроссинговера происходит рекомбинация, то есть обмен участками между отцовской и материнской хромосомами. В результате качественно изменяется генетический состав каждой из хромосом. В первом делении мейоза расходятся гомологичные хромосомы (а не сестринские хроматиды, как в митозе), вследствие чего образуются клетки с гаплоидным набором хромосом, каждая из которых содержит по 22 удвоенные аутосомы и одной удвоенной половой хромосоме. Между первым и вторым делениями мейоза нет периода S (рис.2, справа), а в дочерние клетки во втором делении расходятся сестринские хроматиды. В итоге образуются клетки с гаплоидным набором хромосом, в которых вдвое меньше ДНК, чем в диплоидных соматических клетках в периоде G1, и в 4 раза меньше - чем в соматических клетках по окончании периода S. При оплодотворении число хромосом и содержание ДНК у зиготы становится таким же, как в соматической клетке в периоде G1. Период S в зиготе открывает путь к регулярному делению, характерному для соматических клеток. Фазы Клеточный цикл эукариот разделяют на четыре фазы. В стадии конденсированные метафазные хромосомы поровну распределяются между дочерними клетками. Митоз был первой идентифицированной фазой клеточного цикла, а все остальные события, происходящие в клетке между двумя митозами, были названы интерфазой. Развитие исследований на молекулярном уровне позволило выделить в интерфазе стадию синтеза ДНК, получившую название S-фазы (synthesis). Эти две ключевые стадии клеточного цикла не переходят непосредственно одна в другую. После окончания митоза до начала синтеза ДНК имеет место G1-фаза клеточного цикла (gap), кажущаяся пауза в активности клетки, во время которой внутриклеточные синтетические процессы подготавливают репликацию генетического материала. Второй перерыв в видимой активности (фаза G2) наблюдается после окончания синтеза ДНК перед началом митоза. В фазе G2 клетка осуществляет контроль за точностью произошедшей редупликации ДНК и исправляет обнаруженные сбои. В ряде случаев выделяют пятую фазу клеточного цикла (G0), когда после завершения деления клетка не вступает в следующий клеточный цикл длительное время остается в состоянии покоя. Фазы клеточного цикла не имеют четких временных и функциональных границ, однако при переходе от одной фазы к другой происходит упорядоченное переключение синтетических процессов, позволяющее на молекулярном уровне дифференцировать эти внутриклеточные события. 39. Митоз как основной тип деления.открытый и закрытый митоз. Различные типы митоза эукариот. Наиболее простой тип митоза – плевромитоз. При закрытом плевромитозе (расхождение хромосом происходит без нарушения ядерной оболочки) в качестве центров организации микротрубочек (ЦОМТ) участвуют не центриоли, а полярные тельца неопределенной морфологии, от которых отходят микротрубочки. Этих телец два, они расходятся друг от друга, не теряя связи с ядерной оболочкой, и в результате этого образуются два полуверетена, связанные с хромосомами. Весь процесс образования митотического аппарата и расхождения хромосом происходит в этом случае под ядерной оболочкой. Такой тип митоза встречается среди простейших, он широко распространен у грибов. Встречаются формы полузакрытого плевромитоза, когда на полюсах сформированного веретена ядерная оболочка разрушается. Другой формой митоза является ортомитоз. В этом случае ЦОМТ располагаются в цитоплазме, где идет образование двухполюсного веретена. Существуют три формы ортомитоза: открытый (обычный митоз), полузакрытый и закрытый. При полузакрытом ортомитозе образуется бисимметричное веретено с помощью расположенных в цитоплазме ЦОМТ, ядерная оболочка сохраняется в течение всего митоза, за исключением полярных зон. В качестве ЦОМТ здесь могут обнаруживаться массы гранулярного материала или даже центриоли. Эта форма митоза встречается у зеленых, бурых, красных водорослей, у некоторых низших грибов. При закрытом ортомитозе полностью сохраняется ядерная оболочка, под которой образуется настоящее веретено. Микротрубочки формируются в кариоплазме, реже отрастают от внутриядерного ЦОМТ, не связанного (в отличие от плевромитоза) с ядерной оболочкой. Такого типа митозы характерны для деления микронуклеусов инфузорий, но встречаются и у других простейших. При открытом ортомитозе ядерная оболочка полностью распадается. Этот тип деления клеток характерен для животных организмов, некоторых простейших и для клеток высших растений. Эта форма митоза в свою очередь представлена астральным и анастральным типами (рис. 303). Главной особенностью митоза вообще является возникновение веретена деления, образующегося в связи с разнообразными по своему строению ЦОМТ. Основные стадии митоза: 1.Редупликация (самоудвоение) генетической информации материнской клетки и равномерное распределение ее между дочерними клетками. Это сопровождается изменениями структуры и морфологии хромосом, в которых сосредоточено более 90% информации эукариотической клетки 2.Митотический цикл состоит из четырех последовательных периодов: пресинтетического (или постмитотического) G1, синтетического S, постсинтетического (или премитотического) G2 и собственно митоза. Они составляют автокаталитическую интерфазу (подготовительный период). Фазы клеточного цикла: 1) пресинтетическая (G1) (2n2c, где n-число хромосом, c- число молекул). Идет сразу после деления клетки. Синтеза ДНК еще не происходит. Клетка активно растет в размерах, запасает вещества, необходимые для деления: белки (гистоны, структурные белки, ферменты), РНК, молекулы АТФ. Происходит деление митохондрий и хлоропластов (т. е. структур, способных к ауторепродукции). Восстанавливаются черты организации интерфазной клетки после предшествующего деления; 2) синтетическая (S) (2n4c). Происходит удвоение генетического материала путем репликации ДНК. Она происходит полуконсервативным способом, когда двойная спираль молекулы ДНК расходится на две цепи и на каждой из них синтезируется комплементарная цепочка. В итоге образуются две идентичные двойные спирали ДНК, каждая из которых состоит из одной новой и старой цепи ДНК. Количество наследственного материала удваивается. Кроме этого, продолжается синтез РНК и белков. Также репликации подвергается небольшая часть митохонд-риальной ДНК (основная же ее часть реплицируется в G2 период); 3) постсинтетическая (G2) (2n4c). ДНК уже не синтезируется, но происходит исправление недочетов, допущенных при синтезе ее в S период (репарация). Также накапливаются энергия и питательные вещества, продолжается синтез РНК и белков (преимущественно ядерных). S и G2 непосредственно связаны с митозом, поэтому их иногда выделяют в отдельный период — препрофазу. После этого наступает собственно митоз, который состоит из четырех фаз. Процесс деления включает в себя несколько последовательных фаз и представляет собой цикл. Его продолжительность различна и составляет у большинства клеток от 10 до 50 ч. При этом у клеток тела человека продолжительность самого митоза составляет 1—1,5 ч, G2-периода интерфазы — 2—3 ч, S-периода интерфазы — 6—10 ч. 39. Стадии митоза. Процесс митоза принято подразделять на четыре основные фазы: профазу, метафазу, анафазу и телофазу (рис. 1–3). Так как он непрерывен, смена фаз осуществляется плавно — одна незаметно переходит в другую. В профазе увеличивается объем ядра, и вследствие спирализации хроматина формируются хромосомы. К концу профазы видно, что каждая хромосома состоит из двух хроматид. Постепенно растворяются ядрышки и ядерная оболочка, и хромосомы оказываются беспорядочно расположенными в цитоплазме клетки. Центриоли расходятся к полюсам клетки. Формируется ахроматиновое веретено деления, часть нитей которого идет от полюса к полюсу, а часть — прикрепляется к центромерам хромосом. Содержание генетического материала в клетке остается неизменным (2n4c). В метафазе хромосомы достигают максимальной спирализации и располагаются упорядоченно на экваторе клетки, поэтому их подсчет и изучение проводят в этот период. Содержание генетического материала не изменяется (2n4c). В анафазе каждая хромосома «расщепляется» на две хроматиды, которые с этого момента называются дочерними хромосомами. Нити веретена, прикрепленные к центромерам, сокращаются и тянут хроматиды (дочерние хромосомы) к противоположным полюсам клетки. Содержание генетического материала в клетке у каждого полюса представлено диплоидным набором хромосом, но каждая хромосома содержит одну хроматиду (4n4c). В телофазе расположившиеся у полюсов хромосомы деспирализуются и становятся плохо видимыми. Вокруг хромосом у каждого полюса из мембранных структур цитоплазмы формируется ядерная оболочка, в ядрах образуются ядрышки. Разрушается веретено деления. Одновременно идет деление цитоплазмы. Дочерние клетки имеют диплоидный набор хромосом, каждая из которых состоит из одной хроматиды (2n2c). Нетипичные формы митоза К нетипичным формам митоза относятся амитоз, эндомитоз, политения. 1. Амитоз — это прямое деление ядра. При этом сохраняется морфология ядра, видны ядрышко и ядерная мембрана. Хромосомы не видны, и их равномерного распределения не происходит. Ядро делится на две относительно равные части без образования митотического аппарата (системы микротрубочек, центриолей, структурированных хромосом). Если при этом деление заканчивается, возникает двухъядерная клетка. Но иногда перешнуровывается и цитоплазма. Такой вид деления существует в некоторых дифференцированных тканях (в клетках скелетной мускулатуры, кожи, соединительной ткани), а также в патологически измененных тканях. Амитоз никогда не встречается в клетках, которые нуждаются в сохранении полноценной генетической информации, — оплодотворенных яйцеклетках, клетках нормально развивающегося эмбриона. Этот способ деления не может считаться полноценным способом размножения эукариотических клеток. 2. Эндомитоз. При этом типе деления после репликации ДНК не происходит разделения хромосом на две дочерние хроматиды. Это приводит к увеличению числа хромосом в клетке иногда в десятки раз по сравнению с диплоидным набором. Так возникают полиплоидные клетки. В норме этот процесс имеет место в интенсивно функционирующих тканях, например, в печени, где полиплоидные клетки встречаются очень часто. Однако с генетической точки зрения эндомитоз представляет собой геномную соматическую мутацию. 3. Политения. Происходит кратное увеличение содержания ДНК (хромонем) в хромосомах без увеличения содержания самих хромосом. При этом количество хромонем может достигать 1000 и более, хромосомы при этом приобретают гигантские размеры. При политении выпадают все фазы митотического цикла, кроме репродукции первичных нитей ДНК. Такой тип деления наблюдается в некоторых высокоспециализированных тканях (печеночных клетках, клетках слюнных желез двукрылых насекомых). По-литенные хромосомы дрозофил используются для построения цитологических карт генов в хромосомах. Биологическое значение митоза.Оно состоит в том, что митоз обеспечивает наследственную передачу признаков и свойств в ряду поколений клеток при развитии многоклеточного организма. Благодаря точному и равномерному распределению хромосом при митозе все клетки единого организма генетически одинаковы.Митотическое деление клеток лежит в основе всех форм бесполого размножения как у одноклеточных, так и у многоклеточных организмов. Митоз обусловливает важнейшие явления жизнедеятельности: рост, развитие и восстановление тканей и органов и бесполое размножение организмов. 40.митоз,общие черты и отличия.Особенности митоза у растений и у животных:
41.Мейоз значение, характеристика фаз, отличие от митоза. В отличие от митоза этот механизм в итоге приводит к образованию двух клеток с набором хромосом в 2 раза меньше исходного. Таким образом процесс мейоза служит переходом от диплоидной фазы к гаплоидной, причем в первую очередь речь идет о делении ядра, а уже во вторую - всей клетки. Восстановление же полного набора хромосом происходит в результате дальнейшего слияния гамет. В связи с уменьшением количества хромосом этот метод еще определяют как редукционное деление клетки. Мейоз и его фазы изучали такие известные ученые, как В. Флеминг, Э. Страсбургрер, В. И. Беляев и другие. Исследование этого процесса в клетках как растений, так и животных, продолжается до сих пор - настолько он сложен. Изначально этот процесс считался вариантом митоза, однако практически сразу после открытия он все-таки был выделен как отдельный механизм. Характеристика мейоза и его теоретическое значение были впервые в достаточной степени описаны Августом Вайсманом еще в 1887 году. С тех пор изучение процесса редукционного деления сильно продвинулось, но сделанные выводы пока не были опровергнуты. Мейоз не следует путать с гаметогенезом, хотя оба эти процесса тесно связаны. В образовании половых клеток участвуют оба механизма, однако между ними есть ряд серьезных отличий. Мейоз происходит в две стадии деления, каждая из которых состоит из 4 основных фаз, между ними есть короткий перерыв. Длительность всего процесса зависит от количества ДНК в ядре и структуры хромосомной организации. В целом он гораздо более продолжителен в сравнении с митозом. Кстати, одна из основных причин значительного видового разнообразия - именно мейоз. Набор хромосом в результате редукционного деления разбивается надвое, так что появляются новые комбинации генов, в первую очередь потенциально повышающие приспособляемость и адаптивность организмов, в итоге получающих те или иные наборы признаков и качеств. Фазы мейоза Как уже было упомянуто, редукционное клеточное деление условно делят на две стадии. Каждая из этих стадий разделена еще на 4. И первая фаза мейоза - профаза I в свою очередь подразделяется еще на 5 отдельныхэтапов 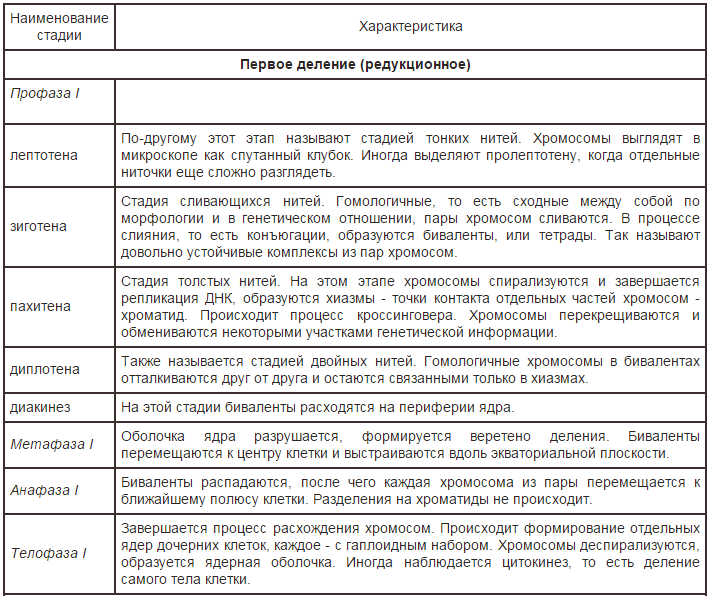  Отличия мейоза от митоза 1. В митозе одно деление, а в мейозе – два (из-за этого получается 4 клетки). 2. В профазе первого деления мейоза происходит конъюгация (тесное сближение гомологичных хромосом) и кроссинговер (обмен участками гомологичных хромосом), это приводит к перекомбинации (рекомбинации) наследственной информации. 3. В анафазе первого деления мейоза происходит независимое расхождение гомологичных хромосом (к полюсам клетки расходятся двуххроматидные хромосомы). Это приводит к рекомбинации и редукции. 4. В интерфазе между двумя делениями мейоза удвоения хромосом не происходит, поскольку они и так двойные. 5. После митоза получается две клетки, а после мейоза – четыре. 6. После митоза получаются соматические клетки (клетки тела), а после мейоза – половые клетки (гаметы – сперматозоиды и яйцеклетки; у растений после мейоза получаются споры). 7. После митоза получаются одинаковые клетки (копии), а после мейоза – разные (происходит рекомбинация наследственной информации). 8. После митоза количество хромосом в дочерних клетках остается таким же, как было в материнской, а после мейоза уменьшается в 2 раза (происходит редукция числа хромосом; если бы её не было, то после каждого оплодотворения число хромосом возрастало бы в два раза; чередование редукции и оплодотворения обеспечивает постоянство числа хромосом |