1 Открытие днк и доква ее генетической роли
 Скачать 347.71 Kb. Скачать 347.71 Kb.
|

|
1) Открытие днк и док-ва ее генетической роли Г. Мендель (1865) высказал первые предположения об организации наследственного материала. На основании результатов своих экспериментов на горохе он пришел к заключению, что наследственный материал дискретен, т.е. представлен отдельными наследственными задатками, отвечающими за развитие определенных признаков организмов. По утверждению Менделя, в наследственном материале организмов, размножающихся половым путем, развитие отдельного признака обеспечивается парой аллельных задатков, пришедших с половыми клетками от обоих родителей. При образовании гамет в каждую из них попадает лишь один из пары аллельных задатков, поэтому гаметы всегда «чисты». В 1909 г. В. Иогансен назвал «наследственные задатки» Менделя генами ДНК как химическое вещество была выделена Иоганном Фридрихом Мишером в 1869 году из остатков клеток, содержащихся в гное. Он выделил вещество, в состав которого входят азот и фосфор. Вначале новое вещество получило название нуклеин, а позже, когда Мишер определил, что это вещество обладает кислотными свойствами, вещество получило название нуклеиновая кислота. Биологическая функция новооткрытого вещества была неясна, и долгое время ДНК считалась запасником фосфора в организме. Более того, даже в начале XX века многие биологи считали, что ДНК не имеет никакого отношения к передаче информации, поскольку строение молекулы, по их мнению, было слишком однообразным и не могло содержать закодированную информацию. В 1928 г. Ф. Гриффитом был поставлен опыт на пневмококках, в котором наблюдалось изменение (трансформация) некоторых наследственных свойств одного бактериального штамма под влиянием материала, полученного из убитых клеток другого штамма. Химическая природа вещества, трансформирующего наследственные свойства бактерий, была установлена лишь в 1944 г. О. Эйвери, доказавшим его принадлежность к нуклеиновым кислотам (ДНК). Другими доказательствами участия ДНК в обеспечении наследственности и изменчивости являются: 1. постоянство содержания ДНК во всех типах соматических клеток организма; 2. соответствие содержания ДНК плоидности клеток (в соматических клетках ее вдвое больше, чем в половых, в полиплоидных клетках оно соответствует количеству наборов хромосом); 3. явление генетической рекомбинации у бактерий при их конъюгации, в ходе которой осуществляется проникновение части ДНК из одной клетки в другую и изменение свойств последней; 4. изменение наследственных свойств бактериальных клеток путем переноса ДНК от одного штамма к другому с помощью ДНК-фага — явление трансдукции; 5. инфицирующая активность изолированной нуклеиновой кислоты вирусов Важным результатом целенаправленного изучения нуклеиновых кислот было создание Дж. Уотсоном и Ф. Криком (1953) пространственной модели молекулы ДНК. 2) Строение нулеотида ДНК (тут крч и по рнк тож) Это полимеры, состоящие из мономеров — нуклеотидов, включающих три компонента: сахар (пентозу), фосфат и азотистое основание (пурин или пиримидин). К первому атому углерода в молекуле пентозы С-1' присоединяется азотистое основание (аденин, гуанин, цитозин, тимин или урацил), а к пятому атому углерода С-5' с помощью эфирной связи — фосфат; у третьего атома углерода С-3' всегда имеется гидроксильная группа — ОН (рис. 3.1). Соединение нуклеотидов в макромолекулу нуклеиновой кислоты происходит путем взаимодействия фосфата одного нуклеотида с гидроксилом другого так, что между ними устанавливается фосфодиэфирная связь (рис. 3.2). В результате образуется полинуклеотидная цепь. Остов цепи состоит из чередующихся молекул фосфата и сахара. К молекулам пентозы в положении С-1' присоединено одно из перечисленных выше азотистых оснований 3) Типы нуклеотидов Когда нуклеотиды полимеризуются или объединяются вместе, они образуют нуклеиновую кислоту, такую как ДНК или РНК. Каждый нуклеотидный фосфат присоединяется к другому сахару, образуя сахар-фосфатную основу с азотистыми основаниями. Нуклеозид является частью нуклеотида, который состоит только из сахара и основания. Таким образом, мы можем говорить о нуклеотиде как о нуклеозиде и фосфатах: Нуклеозид монофосфат представляет собой нуклеотид, который включает в себя один фосфат. Нуклеозид дифосфат представляет собой нуклеотид, который включает в себя два фосфата. Нуклеозид трифосфат представляет собой нуклеотид, который содержит три фосфата. Нуклеотиды являются строительными блоками ДНК и РНК. В одной молекуле ДНК насчитывается 10-25 тыс. нуклеотидов четырех типов, различающихся по азотистому основанию: в адениловый (А) нуклеотид входит аденин, в гуаниловый (Г) — гуанин, в тимидиловый (Т) — тимин, в цитидиловый (Ц) — цитозин. В состав РНК входят нуклеотиды четырех типов, различающиеся по азотистому основанию: аденин входит в состав аденилово- го нуклеотида (А), гуанин — гуанилового (Г), урацил — уриди- лового (У), цитозин — цитидилового (Ц). 4) Первичная, вторичная и третичная структуры молекулы ДНК см тетр 5) Основные св-ва ДНК 2 основополагающий свойства - наследственность и изменчивость (еще ясен красен репликация и репарация) мб еще участие в матричном синтезе рнк (транскрипция), крч можно еще по тетрадке посмореть Перечисленные особенности химической структуры и свойств ДНК обусловливают выполняемые ею функции. ДНК хранит, передает и реализует наследственную информацию Функциональные возможности генетического материала обеспечиваются протеканием четырех генетических процессов– репликацией и обеспечиваются протеканием четырех генетических процессов– репликацией и репарацией ДНК, транскрипцией и генетической рекомбинацией. 6) Механизм репликации Это свойство обеспечивается особенностями химической организации молекулы ДНК, состоящей из двух комплементарных цепей. В процессе репликации на каждой полинуклеотидной цепи материнской молекулы ДНК синтезируется комплементарная ей цепь. В итоге из одной двойной спирали ДНК образуются две идентичные двойные спирали. Такой способ удвоения молекул, при котором каждая дочерняя молекула содержит одну материнскую и одну вновь синтезированную цепь, называют полуконсервативным Инициация репликации осуществляется в особых участках ДНК, обозначаемых ori (от англ. origin —начало). Они включают последовательность, состоящую из 300 нуклеотидных пар, узнаваемую специфическими белками. Двойная спираль ДНК в этих локусах разделяется на две цепи, при этом, как правило, по обе стороны от точки начала репликации образуются области расхождения полинуклеотидных цепей — репликационные вилки, которые движутся в противоположных от локуса ori направлениях. Между репликационными вилками образуется структура, называемая репликаци-онным глазком, где на двух цепях материнской ДНК образуются новые полинуклеотидные цепи С помощью фермента геликазы, разрывающего водородные связи, двойная спираль ДНК расплетается в точках начала репликации. Образующиеся при этом одинарные цепи ДНК связываются специальными дестабилизирующими белками, которые растягивают остовы цепей, делая их азотистые основания доступными для связывания с комплементарными нуклеотидами, находящимися в нуклеоплазме. На каждой из цепей, образующихся в области репликационной вилки, при участии фермента ДНК-полимеразы осуществляется синтез комплементарных цепей 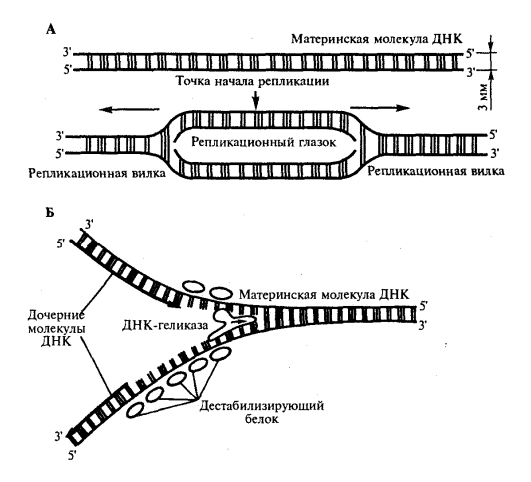 3.8 3.8В процессе синтеза репликационные вилки движутся вдоль материнской спирали в противоположных направлениях, захватывая все новые зоны Разделение спирально закрученных цепей родительской ДНК ферментом геликазой вызывает появление супервитков перед репликационной вилкой. Это объясняется тем, что при расхождении каждых 10 пар нуклеотидов, образующих один виток спирали, родительская ДНК должна совершить один полный оборот вокруг своей оси. Следовательно, для продвижения репликационной вилки вся молекула ДНК перед ней должна была бы быстро вращаться, что потребовало бы большой затраты энергии. В действительности это не наблюдается благодаря особому классу белков, называемых ДНК-топоизомеразами. Топоизомераза разрывает одну из цепей ДНК, что дает ей возможность вращаться вокруг второй цепи. Это ослабляет накопившееся напряжение в двойной спирали ДНК К высвобождающимся водородным связям нуклеотидных последовательностей разделенных родительских цепей присоединяются свободные нуклеотиды из нуклеоплазмы, где они присутствуют в виде дезоксирибонуклеозидгрифосфатов: дАТФ, дГТФ, дЦТФ, дТТФ. Комплементарный нуклеозидтрифосфат образует водородные связи с определенным основанием материнской цепи ДНК. Затем при участии фермента ДНК-полимеразы он связывается фосфодиэфирной связью с предшествующим нуклеотидом вновь синтезируемой цепи, отдавая при этом неорганический пирофосфат (рис. 3.10). Поскольку ДНК-полимераза присоединяет очередной нуклеотид к ОН-группе в 3'-положении предшествующего нуклеотида, цепь постепенно удлиняется на ее 3'- конце. Особенностью ДНК-полимеразы является ее неспособность начать синтез новой полинуклеотидной цепи путем простого связывания двух нуклеозидтрифосфатов: необходим 3'-ОН-конец какой-либо полинуклеотидной цепи, спаренной с матричной цепью ДНК, к которой ДНК-полимераза может лишь добавлять новые нуклеотиды. Такую полинук-леотиднуй цепь называют затравкой или праймером. Роль затравки для синтеза полинуклеотидных цепей ДНК в ходе репликации выполняют короткие последовательности РНК, образуемые при участии фермента РНК-праймазы (рис. 3.11). Указанная особенность ДНК-полимеразы означает, что матрицей при репликации может служить лишь цепь ДНК, несущая спаренную с ней затравку, которая имеет свободный 3'-ОН-конец 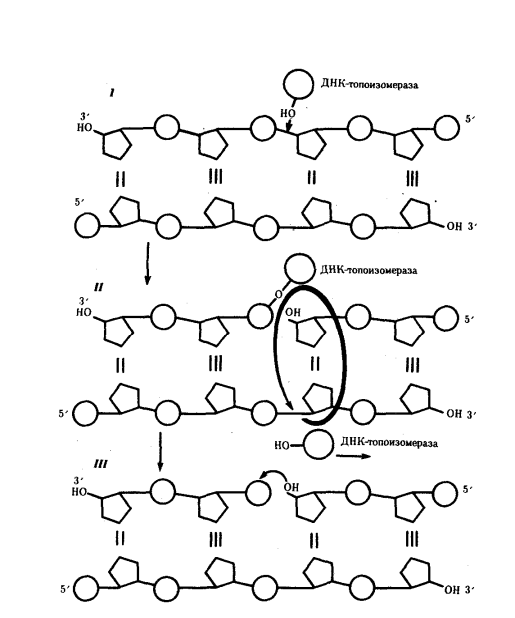 3.9 Разрыв одной из цепей ДНК с помощью фермента ДНК-топоизомеразы I — ДНК-топоизомераза образует ковалентаую связь с одной из фосфатных групп ДНК (верхняя цепь); II — в результате разрыва фосфодиэфирной связи в одной полинуклеотидной цепи вокруг соответствующей ей связи другой цепи осуществляется вращение, которое снимает напряжение, вызванное расхождением двух цепей ДНК в области репликационной вилки; III — после снятия напряжения в спирали ДНК происходит спонтанное отделение ДНК-топоизомеразы и восстановление фосфодиэфирной связи в цепи ДНК Способность ДНК-полимеразы осуществлять сборку полинуклеотида в направлении от 5'- к 3' -концу при антипараллельном соединении двух цепей ДНК означает, что процесс репликации должен протекать на них по-разному. Действительно, если на одной из матриц (3' → 5') сборка новой цепи происходит непрерывно от 5'- к 3'-концу и она постепенно удлиняется на 3'-конце, то другая цепь, синтезируемая на матрице (5' → 3'), должна была бы расти от 3'- к 5'-концу. Это противоречит направлению действия фермента ДНК-полимеразы 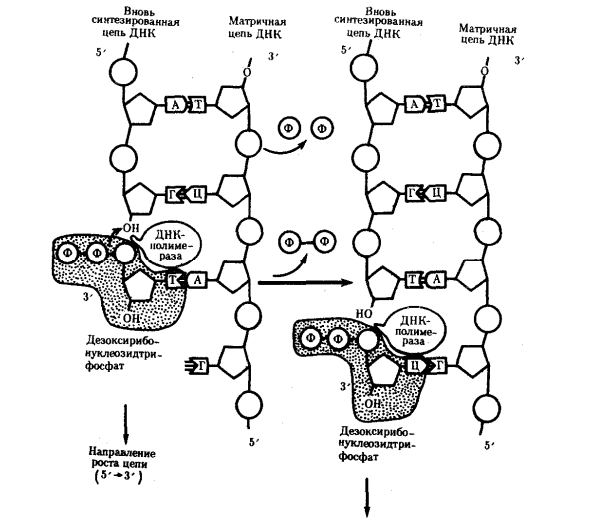 3.10 Присоединение очередного нуклеотида к дочерней цепи ДНК, синтезируемой при участии ДНК-полимеразы: ФФ—пирофосфат В настоящее время установлено, что синтез второй цепи ДНК осуществляется короткими фрагментами (фрагменты Оказаки) также в направлении от 5'- к 3'- концу (по типу шитья «назад иголкой»). У прокариот фрагменты Оказаки содержат от 1000 до 2000 нуклеотидов, у эукариот они значительно короче (от 100 до 200 нуклеотидов). Синтезу каждого такого фрагмента предшествует образование РНКзатравки длиной около 10 нуклеотидов. Вновь образованный фрагмент с помощью фермента ДНК-лигазы соединяется с предшествующим фрагментом после удаления его РНК-затравки (рис. 3.12, А). В связи с указанными особенностями репликационная вилка является асимметричной. Из двух синтезируемых дочерних цепей одна строится непрерывно, ее синтез идет быстрее и эту цепь называют лидирующей. Синтез другой цепи идет медленнее, так как она собирается из отдельных фрагментов,требующих образования, а затем удаления РНК-затравки. Поэтому такую цепь называют запаздывающей (отстающей). Хотя отдельные фрагменты образуются в направлении 5' → 3', в целом эта цепь растет в направлении 3' → 5' (рис. 3.12, А). В виду того, что от локуса ori как правило начинаются две репликационные вилки, идущие в противоположных направлениях, синтез лидирующих цепей в них идет на разных цепях материнской ДНК (рис 3.12, Б). Конечным результатом процесса репликации является образование двух молекул ДНК, нуклеотидная последовательность которых идентична таковой в материнской двойной спирали ДНК. 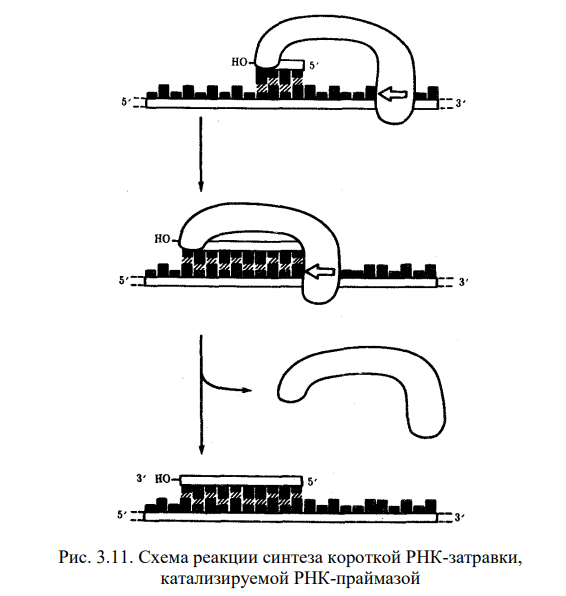 Рассмотренная последовательность событий, происходящих в ходе репликативного синтеза, предполагает участие целой системы ферментов: геликазы, топоизомеразы, дестабилизирующих белков, ДНК-полимеразы и других, совместно действующих в области репликационной вилки (рис 3.13). Репликация ДНК у про- и эукариот в основных чертах протекает сходно, однако, скорость синтеза у эукариот (около 100 нуклеотидов/с) на порядок ниже, чем у прокариот (1000 нуклеотидов/с). Причиной этого может быть образование ДНК эукариот достаточно прочных соединений с белками (см. гл 3.5.2. ), что затрудняет ее деспирали-зацию, необходимую для осуществления репликативного синтеза. Фрагмент ДНК от точки начала репликации до точки ее окончания образует единицу репликации — репликон. Однажды начавшись в точке начала (локус on), репликация продолжается до тех пор, пока весь репликон не будет дуплицирован. Кольцевые молекулы ДНК прокариотических клеток имеют один локус on и представляют собой целиком отдельные репликоны. Эукариотические хромосомы содержат большое число репликонов. В связи с этим удвоение молекулы ДНК, расположенной вдоль эукариотической хромосомы, начинается в нескольких точках. В разных репликонах удвоение может идти в разное время или одновременно. 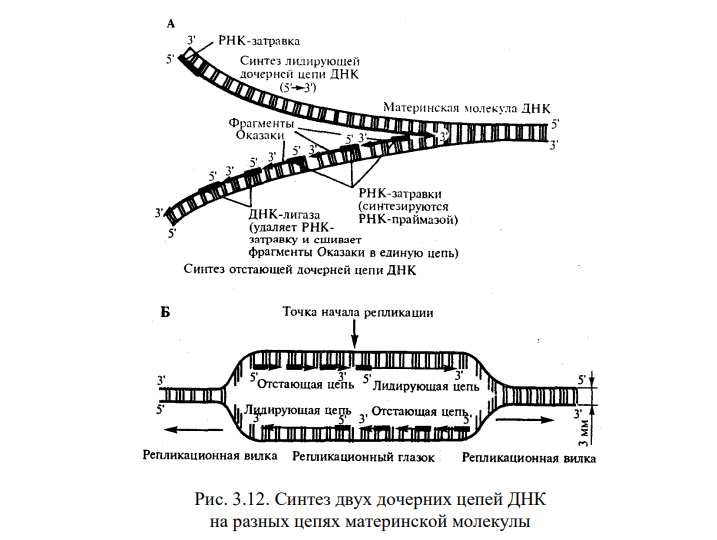 А. В связи с антипараллельностью цепей ДНК синтез дочерних цепей идет по- разному, на верхней материнской цепи дочерняя цель синтезируется непрерывно— лидирующая цепь, на нижней материнской цепи дочерняя цепь собирается из фрагментов Оказаки —отстающая цепь Б. Синтез лидирующих цепей в раэнонаправленных вилках происходит на разных цепях материнской ДНК 7) Видовая специфичность молекул ДНК Генетический код — это последовательность, согласно которой закодированная информация будет преобразована живыми клетками в белки. С первого взгляда генетические коды очень похожи между всеми организмами, и легко изображаются в виде простой таблицы из 64 элементов. Но на самом деле видовая специфичность молекул ДНК у разных видов живых существ уникальна. Генетический код значительно отличается даже внутри одного вида, лишь частично совпадая у родственников. И только однояйцевые близнецы имеют полностью идентичные хромосомы. Каждому животному, как и растению, свойственно определённое количество хромосом с вполне конкретной последовательностью молекул ДНК. Иногда, вследствие мутаций (случайных изменений в структуре ДНК), организм получает свойства, не являющиеся нормой для данного вида. Опираясь на эти знания, учёные начали определять сначала в криминалистике, а позже и в области семейного права, принадлежность биологического материала (волоса, крови, слюны) конкретному человеку. Видовая специфичность молекул днк положена в основу современных методов диагностики инфекций – ПЦР (полимеразная цепная реакция). Проведение ПЦР-исследования позволяет по фрагменту ДНК, содержащемуся в крови или на слизистых оболочках человека, определить вид микроорганизма, вызвавший инфекционное заражение. 8) Уровни организации наследственного материала у эукариот Современные представления о природе генетического аппарата позволяют выделить три уровня его организации: генный, хромосомный и геномный. На каждом из них проявляются основные свойства материала наследственности и изменчивости и определенные закономерности его передачи и функционирования. Элементарной структурой ГЕННОГО уровня организации служит ген. На этом уровне изучается структура молекулы ДНК, биосинтез белка и др. Благодаря относительной независимости генов возможно дискретное (раздельное) и независимое наследование (III закон Менделя) и изменение (мутации) отдельных признаков. Гены клеток эукариот распределены по хромосомам, образуя ХРОМОСОМНЫЙ уровень организации наследственного материала. Этот уровень организации служит необходимым условием сцепления генов и перераспределения генов родителей у потомков при половом размножении (кроссинговер). Вся совокупность генов организма в функциональном отношении ведет себя как целое и образует единую систему, называемую ГЕНОМОМ. Один и тот же ген в разных генотипах может проявлять себя по-разному. Геномный уровень организации объясняет взаимодействие генов как в одной, так и в разных хромосомах. 9) Понятие о гене с молекулярной точки зрения. Свойства гена Ген представляет собой элементарную единицу функции наследственного материала. Это означает, что фрагмент молекулы ДНК, соответствующий отдельному гену и определяющий благодаря содержащейся в нем биологической информации возможность развития конкретного признака, является далее неделимым в функциональном отношении. Сведения о генных мутациях, изложенные выше, указывают на значение изменений химической структуры, затрагивающих не весь ген, а отдельные его участки, вследствие чего появляются новые варианты признака. Минимальное количество наследственного материала, способное, изменяясь, приводить к появлению вариантов признака, соответствует элементарной единице мутационного процесса и называется мутоном. Рассмотренные выше примеры генных мутаций свидетельствуют о том, что достаточно заменить одну пару комплементарных оснований в гене, чтобы изменились свойства кодируемого им белка. Таким образом, мутон соответствует одной паре комплементарных нуклеотидов. |