экзамен по гисте 2 курс. 1. Понятие прогенеза и эмбриогенеза. Периоды и основные стадии эмбриогенеза у человека. Половые клетки человека, их структурногенетическая характеристика
 Скачать 2.3 Mb. Скачать 2.3 Mb.
|

Нервные окончанияНервные волокна заканчиваются концевыми аппаратами — нервными окончаниями. Различают три группы нервных окончаний:
Межнейрональные синапсыСинапсы — это структуры, предназначенные для передачи импульса с одного нейрона на другой или на мышечные и железистые структуры. Синапсы определяют направление проведения импульса. Если раздражать аксон электрическим током, импульс пойдет в обоих направлениях; но импульс, идущий в сторону тела нейрона и его дендритов, не может быть передан на другие нейроны. Только импульс, достигающий терминалей аксона, с помощью синапсов может передать возбуждение на другой нейрон, мышечную или железистую клетку. В зависимости от способа передачи импульса синапсы могут быть химическими или электрическими (электротоническими). 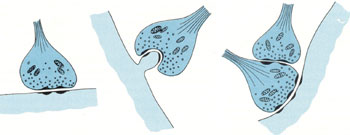 В зависимости от локализации окончаний терминальных веточек аксона, межнейрональные синапсы различают: аксо-дендритические, аксо-соматические, аксо-аксональные. Химические синапсы передают импульс на другую клетку с помощью специальных биологически активных веществ — нейромедиаторов, или нейротрансмиттеров, находящихся в синаптических пузырьках. Терминаль аксона представляет собой пресинаптическую часть, а область второго нейрона, или другой иннервируемой клетки, с которой она контактирует, — постсинаптическую часть. В пресинаптической части находятся синаптические пузырьки, многочисленные митохондрии и отдельные нейрофиламенты. Форма и содержимое синаптических пузырьков связаны с функцией синапса. Если передача импульса совершается с помощью медиатора ацетилхолина, - синапсы называют холинергическими, если медиатором служит норадреналин - адренергическими. В зависимости от передаваемого сигнала, нейромедиаторы, и соответственно синапсы, могут быть возбуждающими или тормозными. Такие нейромедиаторы, как дофамин, глицин и гамма-аминомасляная кислота (ГАМК) являются медиаторами тормозящих синапсов. Область синаптического контакта между двумя нейронами состоит из пресинаптической мембраны, синаптической щели и постсинаптической мембраны. Пресинаптическая мембрана — это мембрана клетки, передающей импульс. В этой области локализованы кальциевые каналы, способствующие слиянию синаптических пузырьков с пресинаптической мембраной и выделению медиатора в синаптическую щель. Синаптическая щельмежду пре- и постсинаптической мембранами имеет ширину 20—30 нм. Мембраны прочно прикреплены друг к другу в синаптической области филаментами, пересекающими синаптическую щель. 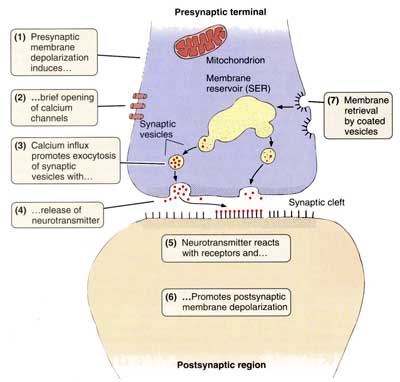 Постсинаптическая мембрана — это участок плазмолеммы клетки, воспринимающий медиаторы и генерирующий импульс. Она снабжена рецепторными зонами для восприятия соответствующего нейромедиатора. В целом процессы в синапсе происходят в следующем порядке: 1. Волна деполяризации доходит до пресинаптической мембраны. 2. При этом открываются кальциевые каналы, и ионы Са2+ входят в терминаль. 3. Повышение концентрации ионов Са2+ в терминали вызывает экзоцитоз нейромедиатора, и медиатор попадает в синаптическую щель. 4. Далее, нейромедиатор диффундирует через синаптическую щель и связывается со специфическими рецепторными участками на постсинаптической мембране, что вызывает молекулярные изменения в постсинаптической мембране, приводящие к открытию ионных каналов и созданию постсинаптических потенциалов, обусловливающих реакции возбуждения или торможения. Электрические, или электротонические, синапсы в нервной системе млекопитающих встречаются относительно редко. В области таких синапсов цитоплазмы соседних нейронов связаны щелевидными соединениями, обеспечивающими прохождение ионов из одной клетки в другую, а следовательно, электрическое взаимодействие этих клеток. Эти синапсы способствуют синхронизации нейральной активности. Эффекторные нервные окончанияСреди эффекторных нервных окончаний различают двигательные и секреторные. Двигательные нервные окончания — это концевые аппараты аксонов двигательных клеток соматической или вегетативной нервной системы. При их участии нервный импульс передается на ткани рабочих органов. 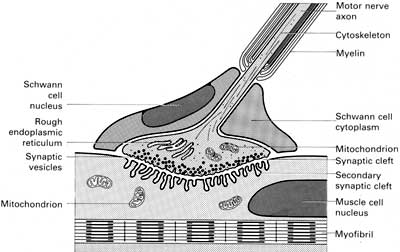 Двигательные окончания в поперечнополосатых мышцах называются нервно-мышечными окончаниями. Они представляют собой окончания аксонов клеток двигательных ядер передних рогов спинного мозга или моторных ядер головного мозга. Нервно-мышечное окончание состоит из концевого ветвления осевого цилиндра нервного волокна и специализированного участка мышечного волокна. Миелиновое нервное волокно, подойдя к мышечному волокну, теряет миелиновый слой и погружается в него, вовлекая за собой его плазмолемму и базальную мембрану. Мембрана мышечного волокна образует многочисленные складки, формирующие вторичные синаптические щели эффекторного окончания. В области окончания мышечное волокно не имеет типичной поперечной исчерченности и характеризуется обилием митохондрий, скоплением круглых или слегка овальных ядер. Терминальные ветви нервного волокна в нервно-мышечном соединении характеризуются обилием митохондрий и многочисленными пресинаптическими пузырьками, содержащими характерный для этого вида окончаний медиатор — ацетилхолин. При возбуждении ацетилхолин поступает через пресинаптическую мембрану в синаптическую щель нахолинорецепторы постсинаптической (мышечной) мембраны, вызывая ее возбуждение (волну деполяризации).  Постсинаптическая мембрана моторного нервного окончания содержит фермент ацетилхолинэстеразу, разрушающий медиатор и ограничивающий этим срок его действия. Двигательные нервные окончания в гладкой мышечной ткани представляют собой чёткообразные утолщения (или варикозы) нервного волокна, идущего среди неисчерченных гладких миоцитов. Варикозы содержат адренергические или холинергическиепресинаптические пузырьки. Нейролеммоциты в области варикозов часто отсутствуют, и волокно проходит «обнаженным». Сходное строение имеют секреторные нервные окончания (нейрожелезистые). Они представляют собой концевые утолщения терминали или утолщения по ходу нервного волокна, содержащие пресинаптические пузырьки, главным образом холинергические. Рецепторные нервные окончанияЭти нервные окончания —рецепторы — рассеяны по всему организму и воспринимают различные раздражения как из внешней среды, так и от внутренних органов. Соответственно выделяют две большие группы рецепторов: экстерорецепторы и интерорецепторы. К экстерорецепторам(внешним) относятся: слуховые, зрительные, обонятельные, вкусовые и осязательные рецепторы. К интерорецепторам(внутренним) относятся: висцеро-рецепторы (сигнализирующие о состоянии внутренних органов) и проприорецепторы (или рецепторы опорно-двигательного аппарата). В зависимости от специфичности раздражения, воспринимаемого данным видом рецептора, все чувствительные окончания делят на механорецепторы, барорецепторы, хеморецепторы,терморецепторы и некоторые другие. По особенностям строения чувствительные окончания подразделяют на свободные нервные окончания, т.е. состоящие только из конечных ветвлений осевого цилиндра, и несвободные, содержащие в своем составе все компоненты нервного волокна, а именно ветвления осевого цилиндра и клетки глии. Несвободные окончания, кроме того, могут быть покрыты соединительнотканной капсулой, и тогда они называются инкапсулированными. Несвободные нервные окончания, не имеющие соединительнотканной капсулы, называются неинкапсулированными. Свободные нервные окончания обычно воспринимают холод, тепло и боль. Такие окончания характерны для эпителия. В этом случае миелиновые нервные волокна подходят к эпителиальному пласту, теряют миелин, а осевые цилиндры проникают в эпителий и распадаются там между клетками на тонкие терминальные ветви. Инкапсулированные рецепторы соединительной ткани при всем их разнообразии всегда состоят из ветвления осевого цилиндра и глиальных клеток. Снаружи такие рецепторы покрыты соединительнотканной капсулой. Примером подобных окончаний могут служить весьма распространенные у человека пластинчатые тельца (или тельца Фатера-Пачини). В центре такого тельца располагается внутренняя луковица, или колба, образованная видоизмененными леммоцитами. Миелиновое чувствительное нервное волокно теряет около пластинчатого тельца миелиновый слой, проникает во внутреннюю луковицу и разветвляется. Снаружи тельце окружено слоистой капсулой, состоящей из фибробластов и спирально ориентированных волокон. Заполненные жидкостью пространства между пластинками содержат коллагеновые микрофибриллы. Давление на капсулу передается через заполненные жидкостью пространства между пластинками на внутреннюю луковицу и воспринимается безмиелиновыми волокнами во внутренней луковице. Пластинчатые тельца воспринимают давление и вибрацию. Они присутствуют в глубоких слоях дермы (особенно в коже пальцев), в брыжейке и внутренних органах. К чувствительным инкапсулированным окончаниям относятся осязательные тельца —тельца Мейснера. Эти структуры имеют овоидную форму, располагаются в верхушках соединительнотканных сосочков кожи. Осязательные тельца состоят из видоизмененных нейролеммоцитов — тактильных клеток, расположенных перпендикулярно длинной оси тельца. Части тактильных клеток, содержащие ядра, расположены на периферии, а уплощенные части, обращенные к центру, формируют пластинчатые отростки, интердигитирующие с отростками противоположной стороны. Тельце окружено тонкой капсулой. Миелиновое нервное волокно входит в основание тельца снизу, теряет миелиновый слой и формирует ветви, извивающиеся между тактильными клетками. Коллагеновые микрофибриллы и волокна связывают тактильные клетки с капсулой, а капсулу с базальным слоем эпидермиса, так что любое смещение эпидермиса передается на осязательное тельце Мейснера. К инкапсулированным нервным окончаниям относятся также рецепторы мышц и сухожилий: это нервно-мышечные веретена и нервно-сухожильные веретена. Нервно-мышечные веретена являются сенсорными органами в скелетных мышцах, которые функционируют как рецептор на растяжение. Веретено состоит из нескольких исчерченных мышечных волокон — интрафузальных волокон, заключенных в растяжимую соединительнотканную капсулу. Между капсулой и интрафузальными волокнами имеется заполненное жидкостью пространство. Интрафузальные волокна имеют актиновые и миозиновые миофиламенты только на концах, которые и сокращаются. Рецепторной частью интрафузального мышечного волокна является центральная, несокращающаяся часть. К интрафузальным мышечным волокнам подходят афферентные нервные волокна. При расслаблении (или растяжении) мышцы увеличивается и длина интрафузальных волокон, что регистрируется рецепторами. Одни окончания реагируют на изменение длины мышечного волокна и на скорость этого изменения, другие — реагируют только на изменение длины. При внезапном растяжении в спинной мозг поступает сильный сигнал, вызывающий резкое сокращение мышцы, с которой поступил сигнал, — динамический рефлекс на растяжение. При медленном, длительном растяжении волокна возникает статический сигнал на растяжение. Этот сигнал может поддерживать мышцу в состоянии сокращения в течение нескольких часов. Интрафузальные волокна имеют также эфферентную иннервацию. К ним подходят тонкие моторные волокна, оканчивающиеся аксо-мышечными синапсами на концах мышечного волокна. Вызывая сокращение концевых участков интрафузального волокна, они усиливают растяжение центральной рецепторной его части, повышая реакцию рецептора. Нервно-сухожильные веретена обычно располагаются в месте соединения мышцы с сухожилием. Коллагеновые пучки сухожилия, связанные с 10—15 мышечными волокнами, окружены соединительнотканной капсулой. К нервно-сухожильному веретену подходит толстое миелиновое волокно, которое теряет миелин и образует терминали, ветвящиеся между пучками коллагеновых волокон сухожилия. Сигнал с нервно-сухожильных веретен, вызванный напряжением мышцы, возбуждает тормозные нейроны спинного мозга. Последние тормозят соответствующие двигательные нейроны, предотвращая перерастяжение мышцы. 45. Рефлекторные дуги. Понятие о рефлекторной дуге Нервная ткань входит в состав нервной системы, функционирующей по рефлекторному принципу, морфологическим субстратом которого является рефлекторная дуга. Рефлекторная дуга представляет собой цепь нейронов, связанных друг с другом синапсами и обеспечивающих проведение нервного импульса от рецептора чувствительного нейрона до эфферентного окончания в рабочем органе. Самая простая рефлекторная дуга состоит из двух нейронов — чувствительного и двигательного. В подавляющем большинстве случаев между чувствительными и двигательными нейронами включены вставочные, или ассоциативные, нейроны. У высших животных рефлекторные дуги состоят обычно из многих нейронов и имеют значительно более сложное строение. Конкретные нервные связи будут рассмотрены при изучении нервной системы. 47. Хар-ка нервной системы. Нервы и спинно-мозговые ганлии. Регенерация нервов. |