экзамен по гисте 2 курс. 1. Понятие прогенеза и эмбриогенеза. Периоды и основные стадии эмбриогенеза у человека. Половые клетки человека, их структурногенетическая характеристика
 Скачать 2.3 Mb. Скачать 2.3 Mb.
|

 1. Понятие прогенеза и эмбриогенеза. Периоды и основные стадии эмбриогенеза у человека. Половые клетки человека, их структурно-генетическая характеристика. 1. Понятие прогенеза и эмбриогенеза. Периоды и основные стадии эмбриогенеза у человека. Половые клетки человека, их структурно-генетическая характеристика.Эмбриогенезу человека предшествует прогенез – процессы развития половых клеток, т.е., овогенез и сперматогенез. Первые 3 стадии развития половых клеток схожи и в тоже время имеют различия: I – стадия размножения: происходит размножение стволовых половых клеток путем деления митозом малодифференцированных половых клеток (светлые сперматогонии типа А и овогонии). Стадия размножения у лиц женского пола протекает еще в эмбриональном периоде, у лиц мужского пола – после полового созревания. II – стадия роста: происходит увеличение размеров половых клеток и подготовка к мейозу (происходит в стадии созревания) – синтез ДНК и перекомбинация генов в гомологичных хромосомах при кроссенговере. У лиц мужского пола происходит после полового созревания и при этом сперматогонии превращаются в сперматоциты I порядка, у лиц женского пола эта стадия протекает в 2 периода: “период малого роста” – в эмбриональном периоде происходит незначительное увеличение размеров половой клетки, синтез ДНК и перекомбинация генов в гомологичных хромосомах при кроссенговере, “период большого роста” – после полового созревания, при этом овоцит I порядка почти в 2 раза увеличивается в размерах, приобретает вторичную оболочку, накапливает желток (трофические включения). III – стадия созревания: происходит мейоз (2 быстро следующих друг за другом деления половых клеток без удвоения хромосом), в результате образуются половые клетки с уже с гаплоидным набором хромосом. У мужчин стадия созревания происходит после полового созревания, а у лиц женского пола – начинается еще в эмбриональном периоде и завершается после полового созревания. У мужчин в стадии созревания из одного сперматоцита I порядка образуется 4 сперматида; у женщин первое деление начинается еще в эмбриональном периоде, но остается незавершенным, продолжается и заканчивается после полового созревания и из одного овоцита I порядка образуется один овоцит II порядка и первое редукционное (направительное) тельце, а после второго деления – одна яйцеклетка и второе редукционное тельце. Первое редукционное тельце также может поделиться, поэтому в результате стадии созревания у женщин из одного овоцита I порядка образуется 1 яйцеклетка и 3 (или 2) редукционные тельца. IV – стадия формирования имеет место только при сперматогенезе, при этом сперматиды избавляются от излишнего количества цитоплазмы, покрываются гликокаликсом, приобретают характерный для них структуру (морфологию), т.е. превращаются в сперматозоиды. В результате гаметогенеза образуются половые мужские женские клетки, которые отличаются от соматических клеток по следующим признакам: 1. Набор хромосом: соматические клетки имеют диплоидный, половые клетки – гаплоидный набор хромосом. 2. Половые клетки имеют специальные приспособления для выполнения своих специфических функций: а) сперматозоиды – акрасому (видоизмененный пластинчатый комплекс) для проникновения через оболочки яйцеклетки и двигательный аппарат (хвостик, центриоли и митохондрии); б) яйцеклетки – I и II оболочки, желток (трофические включения). 3. Ядерно-цитоплазматическое отношение половых клеток резко отличается от соматических: в яйцеклетке – очень низкое (резко преобладает масса цитоплазмы), в сперматазоидах – очень высокое (преобладает масса ядра). 4. Жизненный цикл у половых сложный и многостадийный. 5. Биологическое назначение: из соматических клеток могут образоваться только подобные клетки, а из половых клеток – целый новый организм. Яйцеклетка человека имеет диаметр около 130 мкм, окружена первичной (оолеммa или собственная оболочка) и вторичной (блестящая или прозрачная зона — Zоnа ро11ucida=Zр) оболочками и слоем фолликулярных клеток. Блестящая зона состоит из гликопротеинов Zр1, Zр2 и ZрЗ (является специфическим рецептором для сперматозоидов) и гликозаминогликанов. Ядро содержит гаплоидный набор хромосом и хорошо выраженное ядрышко. В цитоплазме имеются гранулярный ЭПС, комплекс Гольджи, умеренное количество митохондрий, гранулы желтка (трофические включения), сразу под цитолеммой — кортикальные гранулы (гранулы диаметром около 1мкм. окружены элементарной .мембраной, содержат ферменты для кортикальной реакции). В яйцеклетки человека же’лтка мало, желток распределяется по цитоплазме равномерно, т.е. яйцеклетка олиголецнтальная и изолецитальная. Процесс внутриутробного развития у человека длится .в среднем 280 суток (10 лунных месяцев) и делится на 3 периода: 1. Начальный период (1-ая неделя). 2. Зародышевый период (2-8 недели). 3. Плодный период (с 9-й недели до рождения). Эмбриогенез включает в себя процессы с момента оплодотворения до рождения и включает следующие его дни. 1. Оплодотворение, в результате которого образуется зигота (одноклеточный зародыш), 2. Дробление зародыша с образованием бластулы. 3. Гаструляция — образование 3-х листкового зародыша. 4. Гистогенез, органогенез и ситемагенез — дифференцировка зародышевых листков в ткани органов, образование из органов систем органов. Оплодотворение — это сближение и слияние половых клеток с образованием одноклеточного зародыша — зиготы. У человека оплодотворение внутреннее, т.е. происходит и женских половых путях. В процессе оплодотворения выделяют: 1. Дистантное взаимодействие и сближение половых клеток. 2. Контактное взаимодействие половых клеток и активизация яйцеклетки. 3. Вхождение сперматазоида в в яйцеклетку и последующей синкарион (сингамия)- слияние женского и мужского пронуклеусов. Дистантное взаимодействие (взаимодействие на расстоянии) половых клеток начинается с момента попадания сперматозоидов в женские половые пути, т.е. когда сперматозоиды находятся еще во влагалище: женские половые клетки выделяют специфические вещества — гемогомоны, которые вызывают хемотаксис сперматозоидов — свойство сперматозоидов двигаться против градиента концентрации (туда, где выше концентрация) гемогомонов Хемотаксис сперматозоидов обуславливает их направленное движение к яйцеклетке. Продвижению сперматозоидов к яйцеклетке способствует также реотаксис сперматозоидов — свойство сперматозоидов всегда двигаться против тока жидкосж (слизь в женских половых путях течет по направлению: маточные трубы —> матка —> влагалище). Сперматозоиды двигаются со скоростью 30-50 мкм/сек. Поэгому уже через 30-60 минуг могут попадать в полость матки, а через 1,5-2 часа — ампулярной части маточной трубы. Сразу после эякуляции и в. ближайшие часы сперматозоиды еще не способны оплодотворить яйцеклетку, для этого должна произойти капацнтация сперматозоида -приобретение способности к оплодотворению под воздействием секретов женских половых путей. Для капацитации сперматозоидов в среднем требуется 7 часов. Контактное взаимодействие половых клеток и проникновение сперматозойда в яйцеклетку Многочисленные сперматозоиды окружают яйцеклетку и выделяют ферменты, под воздействием которых о г яйцеклетки отсоединяются фолликулярные клетки. Сперматозоиды синхронным биением жгутиков вращают яйцеклетку со скоростью 4 оборота в минуту (в течении до 12 часов !). При помощи специфических (строго комплементарных для каждого вида) рецепторов сперматозоид (фермент гликозилтрансфераза па повнерхностп акросомы) и яйцеклетка (сахар N-ацетилглюкозамин в составе ZрЗ фракции гликопротеинов блестящей зоны) «узнают» друг друга — т.е. происходит контакт, в результате у сперматозоида просходит акрасомальная реакция — выделение и воздействие на оболочки яйцеклетки ферментов акросомы (гиалуронидаза и трипсин) и головка сперматозоида проникает через блестящую зону и оолемму и цитоплазму яйцеклетки. У человека и яйцеклетку проникает только один сперматозоид — моноспермия. Проникновению других сперматозоидов (полиспермия) препятствует наступающая через» несколько секунд кортикальная реакция — мембраны кортикальных гранул яйцеклетки сливаются с оолеммой и содержимое гранул воздействует на блестящшую зону (И оболочку яйцеклетки); в результате этого блестящая оболочка утрачивает способность осуществлять специфический контакт со сперматозоидами и запускать у них акрасомальную реакцию, уплотняется и превращается в оболочку оплодотворения, непроницаемую для остальных сперматозоидов. Синкарион. После проникновения в яйцеклетку ядро сперматозоида увеличивается в размерах, хроматин десперализуется. Далее ядра яйцеклетки и сперматозоида (пронуклеусы) сближаются и сливаются (синкарион), а из клеточного цетра сперматозоида начинает формироваться веретено деления — в результате образуется зигота, т.е. одноклеточный зародыш. В зиготе начинается интенсивный синтез белка. — яйцеклетка актиивизируется, начинает готовится к следующей стадии эмбриогенезе — дроблению. Для активизации яйцеклетки проникновение в нее сперматозоида необязательно, достаточно кратковременного контакта между поверхностью яйцеклетки и сперматозоида. Яйцеклетку можно активизировать с помощью множества неспецифических химических и физических факторов (например укол иглы). Развитие яйцеклетки активацией без сперматозоида называется партеногенезом (пример: у некоторых насекомых, у пчел). В настоящее время разработаны методы оплодотворения яйцеклеток млекопитающих и человека in vitro — в пробирке, с последующей пересадкой в матку. Благодаря этой методике бесплодные женщины получили возможность рожать нормальных детей. Дробление — это деление зародыша, но это деление несколько отличается от обычного деления митозом: при дроблении отсутствует пресинтетнческая фаза G1, поэтому дочерние (слетки (бластомеры) после митоза не синтезируют белки и не растут — с каждым делением размеры образовавшихся бластомеров становится все меньше и меньше (отсюда и название «‘дробление»). В результате дробления восстанавливается нормальное ядерно-цитоплазматическое отношение (у оплодотворенной яйцеклетки оно очень низкое) и образуется бластула. Тип дробления зависит от количества и распределения желтка в яйцеклетке. Олиголецитальная изолецитальная яйцеклетка человека дробится по типу: полное неравномерное асинхронное. Полное — в дроблении участвуют все участки оплодотворенной яйцеклетки; неравномерное — образовавшиеся бластомеры не одинаковые, не равные: одни крупные и темные, располагаются в центре зародыша их совокупность называется змбриобластом — будущее тело, другие мелкие и светлые, окружают снаружи бластомеры эмбриобласта — их совокупность называется трофобластом — участвует при формировании плаценты; асинхронное = количество бластомеров увеличивается не по геометрической прогреcии (2-> 4-> 8 и т.д., т.е. кратное увеличение числа блаcтомеров). В то же время следует отметить, что первые бластомеры (по крайней мере до 8 первых бластомеров) потенциально не различаются. Доказательством этого утверждения является образование однояйцевых (идентичных) близнецов. Однояйцевые близнецы образуются из одной яйцеклетки в случае когда по каким-то причинам на стадии 2-х или 3-х бластомерного зародыша происходит их разделение и из каждого бластомера развивается самостоятельный отдельный организм. С другой стороны, если в эксперименте взять двухбластомерного зародыша и разрушить, один бластомер, то из второго бластомера может развиться вполне нормальный организм. Или еще; если взять двух эмбрионов на стадии до 8-бласгомерного этапа и их объединить в одну морулу, то можно получить новый организм, наследующий признаки не 2-х а 4-х родителей — такие экспериментально полученные животные называются химерами. Этими экспериментами было доказано, что бластомеры ранних стадий дробления (вплоть до 8-клеточной стадии) практически идентичны и обладают неограниченными потенциями. С помощью метода химер было доказано, что какие бластомеры дадут трофобласт, а какие — эмбриобласт зависит не от особенностей самих бластомеров, а лишь от места, какое они случайно заняли в моруле. Бластомеры расположенные снаружи морулы подвергаются воздействию одних условий, внутренние бластомеры оказываются в других условиях, что и определяет направление их дальнейшей дифференцировки. Дробление зиготы человека начинается в конце 1-х суток после оплодотворения в дистальной части маточной трубы и заканчивается в 7-е сутки в полости матки. На 2-3-и сутки зародыш находится в маточных трубах и имеет вид плотного узелка — морулы, в центральной части которого находятся крупные темные бластомеры — эмбриоблзст, а по периферии -мелкие светлые бластомеры = трофобласт. В 4-е сутки бластула находится в проксимальной части маточной трубы, т.е. вплотную подходит к полости матки и имеет вид пузырька. Бластомеры трофобласта всасывают секрет маточной трубы и сами секретируют жидкость, поэтому трофобласт растягивается и превращается в пузырек, заполненный жидкостью, а эмбриооласт прикрепляется на одном полюсе к трофобласту снутри. Такая бластула называется эпибластулой (или синонимы; бластоциста, стерробластула). На 5-е сутки бластоциста попадает в полость матки и остается там до 7-х суток, за это время несколько увеличивается в размерах (100 и более бластомеров). Гаструляция — это сложный процесс, где в результате размножения, роста, днфференцировкн н направленного перемещения клеток образуется 3-х листковый зародыш. Гаструляция происходит 7-17 сутки и осуществляется путем деламииацин или расщипления (7-14 сутки) и иммиграции или выселения (14-17 сутки). В 7-е сутки эмбриобласт расщепляется на 2 слоя: верхний слой — эпибласт или первичная эктодерма (содержит материал будущей эктодермы, мезодермы, хорды и части энтодермы) и нижний слой — гипобласт (будущая энтодерма после присоединения клеточного материала прехордальной пластинки из эпибласта). Почти одновременно с этим происходит выселение клеток из эпи- и гипооласта — внезародышевая мезенхима, которая выстилает внутреннюю поверхность трофобласта. Далее в течение 2-й недели эпибласт и гипобласт начинают прогибаться в противоположных направлениях и превращаются в пузырьки: из эпибласта образуется амниотнческий пузырек, нз гипобласта — желточный пузырек. Эти 2 пузырька окружаются внезародышевой мезенхимой. Соприкасающиеся поверхности амниотнческого и желточного пузырька имеют вид диска (или щитка) и соответственно называются зародышевым эпибластом и зародышевым гипобластом, а вместе — зародышевым щитком. Остальные участки амниогического и желточного пузырька называются внезародышевым эпи- и гипобластом. В начале 3-й недели (14-17 сутки) происходит иммиграция (выселение) клеток из эпибласта, причем это происходит в 2 фазы: в I фазе идет подготовка к выселению — клеточный материал подлежащий иммиграции перемещается (медленно двигающиеся клетки: с будущего краниального конца к каудальному концу по центру эпибласта, а быстродвигаюшиеся клетки: вначале тоже с краниального полюса к каудальному полюсу, но по краю э:.’нблпста, а затем поворачиваются и идут но центру эпибласта к краниальному концу) и собирается вместе и образует на поверхности эпибласта 3 структуры: прехордальную пластинку, I узелок и I полоску; II фаза — собственно выселение материала этих 3- структур. I узелок выселяется и между имеющимися двумя листками образует первый осевой орган — хорду. Материал прехордальной пластинки выселяется и присоединяется к гипобласту, с этого момента нижним листок будет называться энтодермой. Клетки 1 полоски выселяется, занимает положение между двумя имеющимися листками и образует средний листок — мезодерму. Оставшаяся часть эпибласта после выселения клеток 3-х структур будет называться эктодермой. В следующей стадии начинается дифференцировка зародышевых листков в ткани (гистогенез) органов (органогенез) и формирование из органов систем органов (системогенез). При этом следует выделить 20-21 сутки эмбриогенеза — в эти сроки происходят следующие важные процессы: 1 Из всех трех зародышевых листков, но преимущественно нз мезодермы, выселяются клетки, заполняют пространства между тремя зародышевыми листками, т.е. формируется зародышевая мезенхима. 2. Мезодерма дифференцируется на составные части (томы) сомиты, сегментные ножки и спланхнотомы. 3 Трехлистковый плоским зародыш сворачивается «в трубку» — формируется туловище (энтодерма сворачиваясь в трубку превращается к I кишку, окруженную слоем мезенхимы и сплапхнатомами; эктодерма покрывает туловище снаружи). 4. При сворачивании зародыша » в трубку» внезародышевые части организма обособляются от тела зародыша и формируются провизорные органы желточный мешок, гипнотическая оболочка. Материал зародышевых листков дифференнцируется в ткани, органы и системы органов ( более подробно о развитии конкретных органов и систем смотри в лекциях по частной гистологии): I Эктодерма: - нервная ткань органов нервной системы; - эпидермис кожи и его производные (сальные, потовые, молочные железы, ногти, волосы); - эпителии роговицы и хрусталик глаза, эпителий преддверья ротовой полости и анального отдела прямой кишки; II Мезодерма: -дерматомы -> дерма кожи; - миотомы —> скелетная мускулатура; - склеротомы —> осевой скелет (кости и хрящи позвоночного столба; - сегментные ножки (нефрототомы) —► эпителий мочеполовой системы; - спланхиотомы -> мезотелий серозных покровов (брюшины, плевры и околосердечной сумки), эпителий гонад (клетки Сертоли яичка и фолликулярные клетки яичников), корковая часть надпочечников, миокард и эпикард; III Энтодерма: - эпителий и железы (включая печень и поджелудочную железу) пищеварительном и дыхательной системы; IV Мезенхима: ткани внутренней среды (кровь и лимфа, все виды волокнистых соединительных тканей — рыхлая волокнистая соединительная ткань, плотная волокнистая оформленная и неоформленная волокнистая соединительные ткани, соединительные ткани со специальными свойствами, костные и хрящевые ткани) и гладкая мышечная ткань. 2. Основные стадии эмбриогенеза. Понятие оплодотворения. Характеристика оплодотворения у человека: морфология, необходимые условия. Понятие зиготы. 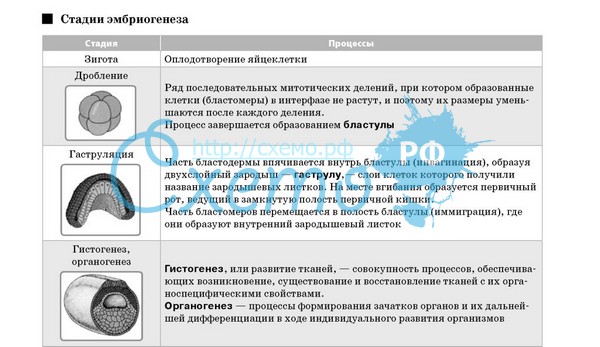 Оплодотворение — это сближение и слияние половых клеток с образованием одноклеточного зародыша — зиготы. У человека оплодотворение внутреннее, т.е. происходит и женских половых путях. В процессе оплодотворения выделяют: 1. Дистантное взаимодействие и сближение половых клеток. 2. Контактное взаимодействие половых клеток и активизация яйцеклетки. 3. Вхождение сперматазоида в в яйцеклетку и последующей синкарион (сингамия)- слияние женского и мужского пронуклеусов. Дистантное взаимодействие (взаимодействие на расстоянии) половых клеток начинается с момента попадания сперматозоидов в женские половые пути, т.е. когда сперматозоиды находятся еще во влагалище: женские половые клетки выделяют специфические вещества — гемогомоны, которые вызывают хемотаксис сперматозоидов — свойство сперматозоидов двигаться против градиента концентрации (туда, где выше концентрация) гемогомонов Хемотаксис сперматозоидов обуславливает их направленное движение к яйцеклетке. Продвижению сперматозоидов к яйцеклетке способствует также реотаксис сперматозоидов — свойство сперматозоидов всегда двигаться против тока жидкосж (слизь в женских половых путях течет по направлению: маточные трубы —> матка —> влагалище). Сперматозоиды двигаются со скоростью 30-50 мкм/сек. Поэгому уже через 30-60 минуг могут попадать в полость матки, а через 1,5-2 часа — ампулярной части маточной трубы. Сразу после эякуляции и в. ближайшие часы сперматозоиды еще не способны оплодотворить яйцеклетку, для этого должна произойти капацнтация сперматозоида -приобретение способности к оплодотворению под воздействием секретов женских половых путей. Для капацитации сперматозоидов в среднем требуется 7 часов. Контактное взаимодействие половых клеток и проникновение сперматозойда в яйцеклетку Многочисленные сперматозоиды окружают яйцеклетку и выделяют ферменты, под воздействием которых о г яйцеклетки отсоединяются фолликулярные клетки. Сперматозоиды синхронным биением жгутиков вращают яйцеклетку со скоростью 4 оборота в минуту (в течении до 12 часов !). При помощи специфических (строго комплементарных для каждого вида) рецепторов сперматозоид (фермент гликозилтрансфераза па повнерхностп акросомы) и яйцеклетка (сахар N-ацетилглюкозамин в составе ZрЗ фракции гликопротеинов блестящей зоны) «узнают» друг друга — т.е. происходит контакт, в результате у сперматозоида просходит акрасомальная реакция — выделение и воздействие на оболочки яйцеклетки ферментов акросомы (гиалуронидаза и трипсин) и головка сперматозоида проникает через блестящую зону и оолемму и цитоплазму яйцеклетки. У человека и яйцеклетку проникает только один сперматозоид — моноспермия. Проникновению других сперматозоидов (полиспермия) препятствует наступающая через» несколько секунд кортикальная реакция — мембраны кортикальных гранул яйцеклетки сливаются с оолеммой и содержимое гранул воздействует на блестящшую зону (И оболочку яйцеклетки); в результате этого блестящая оболочка утрачивает способность осуществлять специфический контакт со сперматозоидами и запускать у них акрасомальную реакцию, уплотняется и превращается в оболочку оплодотворения, непроницаемую для остальных сперматозоидов. Синкарион. После проникновения в яйцеклетку ядро сперматозоида увеличивается в размерах, хроматин десперализуется. Далее ядра яйцеклетки и сперматозоида (пронуклеусы) сближаются и сливаются (синкарион), а из клеточного цетра сперматозоида начинает формироваться веретено деления — в результате образуется зигота, т.е. одноклеточный зародыш. В зиготе начинается интенсивный синтез белка. — яйцеклетка актиивизируется, начинает готовится к следующей стадии эмбриогенезе — дроблению. Для активизации яйцеклетки проникновение в нее сперматозоида необязательно, достаточно кратковременного контакта между поверхностью яйцеклетки и сперматозоида. Яйцеклетку можно активизировать с помощью множества неспецифических химических и физических факторов (например укол иглы). Развитие яйцеклетки активацией без сперматозоида называется партеногенезом (пример: у некоторых насекомых, у пчел). В настоящее время разработаны методы оплодотворения яйцеклеток млекопитающих и человека in vitro — в пробирке, с последующей пересадкой в матку. Благодаря этой методике бесплодные женщины получили возможность рожать нормальных детей. Дробление — это деление зародыша, но это деление несколько отличается от обычного деления митозом: при дроблении отсутствует пресинтетнческая фаза G1, поэтому дочерние (слетки (бластомеры) после митоза не синтезируют белки и не растут — с каждым делением размеры образовавшихся бластомеров становится все меньше и меньше (отсюда и название «‘дробление»). В результате дробления восстанавливается нормальное ядерно-цитоплазматическое отношение (у оплодотворенной яйцеклетки оно очень низкое) и образуется бластула. Тип дробления зависит от количества и распределения желтка в яйцеклетке. Олиголецитальная изолецитальная яйцеклетка человека дробится по типу: полное неравномерное асинхронное. Полное — в дроблении участвуют все участки оплодотворенной яйцеклетки; неравномерное — образовавшиеся бластомеры не одинаковые, не равные: одни крупные и темные, располагаются в центре зародыша их совокупность называется змбриобластом — будущее тело, другие мелкие и светлые, окружают снаружи бластомеры эмбриобласта — их совокупность называется трофобластом — участвует при формировании плаценты; асинхронное = количество бластомеров увеличивается не по геометрической прогреcии (2-> 4-> 8 и т.д., т.е. кратное увеличение числа блаcтомеров). В то же время следует отметить, что первые бластомеры (по крайней мере до 8 первых бластомеров) потенциально не различаются. Доказательством этого утверждения является образование однояйцевых (идентичных) близнецов. Однояйцевые близнецы образуются из одной яйцеклетки в случае когда по каким-то причинам на стадии 2-х или 3-х бластомерного зародыша происходит их разделение и из каждого бластомера развивается самостоятельный отдельный организм. С другой стороны, если в эксперименте взять двухбластомерного зародыша и разрушить, один бластомер, то из второго бластомера может развиться вполне нормальный организм. Или еще; если взять двух эмбрионов на стадии до 8-бласгомерного этапа и их объединить в одну морулу, то можно получить новый организм, наследующий признаки не 2-х а 4-х родителей — такие экспериментально полученные животные называются химерами. Этими экспериментами было доказано, что бластомеры ранних стадий дробления (вплоть до 8-клеточной стадии) практически идентичны и обладают неограниченными потенциями. С помощью метода химер было доказано, что какие бластомеры дадут трофобласт, а какие — эмбриобласт зависит не от особенностей самих бластомеров, а лишь от места, какое они случайно заняли в моруле. Бластомеры расположенные снаружи морулы подвергаются воздействию одних условий, внутренние бластомеры оказываются в других условиях, что и определяет направление их дальнейшей дифференцировки. Дробление зиготы человека начинается в конце 1-х суток после оплодотворения в дистальной части маточной трубы и заканчивается в 7-е сутки в полости матки. На 2-3-и сутки зародыш находится в маточных трубах и имеет вид плотного узелка — морулы, в центральной части которого находятся крупные темные бластомеры — эмбриоблзст, а по периферии -мелкие светлые бластомеры = трофобласт. В 4-е сутки бластула находится в проксимальной части маточной трубы, т.е. вплотную подходит к полости матки и имеет вид пузырька. Бластомеры трофобласта всасывают секрет маточной трубы и сами секретируют жидкость, поэтому трофобласт растягивается и превращается в пузырек, заполненный жидкостью, а эмбриооласт прикрепляется на одном полюсе к трофобласту снутри. Такая бластула называется эпибластулой (или синонимы; бластоциста, стерробластула). На 5-е сутки бластоциста попадает в полость матки и остается там до 7-х суток, за это время несколько увеличивается в размерах (100 и более бластомеров). Гаструляция — это сложный процесс, где в результате размножения, роста, днфференцировкн н направленного перемещения клеток образуется 3-х листковый зародыш. Гаструляция происходит 7-17 сутки и осуществляется путем деламииацин или расщипления (7-14 сутки) и иммиграции или выселения (14-17 сутки). В 7-е сутки эмбриобласт расщепляется на 2 слоя: верхний слой — эпибласт или первичная эктодерма (содержит материал будущей эктодермы, мезодермы, хорды и части энтодермы) и нижний слой — гипобласт (будущая энтодерма после присоединения клеточного материала прехордальной пластинки из эпибласта). Почти одновременно с этим происходит выселение клеток из эпи- и гипооласта — внезародышевая мезенхима, которая выстилает внутреннюю поверхность трофобласта. Далее в течение 2-й недели эпибласт и гипобласт начинают прогибаться в противоположных направлениях и превращаются в пузырьки: из эпибласта образуется амниотнческий пузырек, нз гипобласта — желточный пузырек. Эти 2 пузырька окружаются внезародышевой мезенхимой. Соприкасающиеся поверхности амниотнческого и желточного пузырька имеют вид диска (или щитка) и соответственно называются зародышевым эпибластом и зародышевым гипобластом, а вместе — зародышевым щитком. Остальные участки амниогического и желточного пузырька называются внезародышевым эпи- и гипобластом. В начале 3-й недели (14-17 сутки) происходит иммиграция (выселение) клеток из эпибласта, причем это происходит в 2 фазы: в I фазе идет подготовка к выселению — клеточный материал подлежащий иммиграции перемещается (медленно двигающиеся клетки: с будущего краниального конца к каудальному концу по центру эпибласта, а быстродвигаюшиеся клетки: вначале тоже с краниального полюса к каудальному полюсу, но по краю э:.’нблпста, а затем поворачиваются и идут но центру эпибласта к краниальному концу) и собирается вместе и образует на поверхности эпибласта 3 структуры: прехордальную пластинку, I узелок и I полоску; II фаза — собственно выселение материала этих 3- структур. I узелок выселяется и между имеющимися двумя листками образует первый осевой орган — хорду. Материал прехордальной пластинки выселяется и присоединяется к гипобласту, с этого момента нижним листок будет называться энтодермой. Клетки 1 полоски выселяется, занимает положение между двумя имеющимися листками и образует средний листок — мезодерму. Оставшаяся часть эпибласта после выселения клеток 3-х структур будет называться эктодермой. В следующей стадии начинается дифференцировка зародышевых листков в ткани (гистогенез) органов (органогенез) и формирование из органов систем органов (системогенез). При этом следует выделить 20-21 сутки эмбриогенеза — в эти сроки происходят следующие важные процессы: 1 Из всех трех зародышевых листков, но преимущественно нз мезодермы, выселяются клетки, заполняют пространства между тремя зародышевыми листками, т.е. формируется зародышевая мезенхима. 2. Мезодерма дифференцируется на составные части (томы) сомиты, сегментные ножки и спланхнотомы. 3 Трехлистковый плоским зародыш сворачивается «в трубку» — формируется туловище (энтодерма сворачиваясь в трубку превращается к I кишку, окруженную слоем мезенхимы и сплапхнатомами; эктодерма покрывает туловище снаружи). 4. При сворачивании зародыша » в трубку» внезародышевые части организма обособляются от тела зародыша и формируются провизорные органы желточный мешок, гипнотическая оболочка. Материал зародышевых листков дифференнцируется в ткани, органы и системы органов ( более подробно о развитии конкретных органов и систем смотри в лекциях по частной гистологии): I Эктодерма: - нервная ткань органов нервной системы; - эпидермис кожи и его производные (сальные, потовые, молочные железы, ногти, волосы); - эпителии роговицы и хрусталик глаза, эпителий преддверья ротовой полости и анального отдела прямой кишки; II Мезодерма: -дерматомы -> дерма кожи; - миотомы —> скелетная мускулатура; - склеротомы —> осевой скелет (кости и хрящи позвоночного столба; - сегментные ножки (нефрототомы) —► эпителий мочеполовой системы; - спланхиотомы -> мезотелий серозных покровов (брюшины, плевры и околосердечной сумки), эпителий гонад (клетки Сертоли яичка и фолликулярные клетки яичников), корковая часть надпочечников, миокард и эпикард; III Энтодерма: - эпителий и железы (включая печень и поджелудочную железу) пищеварительном и дыхательной системы; IV Мезенхима: ткани внутренней среды (кровь и лимфа, все виды волокнистых соединительных тканей — рыхлая волокнистая соединительная ткань, плотная волокнистая оформленная и неоформленная волокнистая соединительные ткани, соединительные ткани со специальными свойствами, костные и хрящевые ткани) и гладкая мышечная ткань. |