экзамен по гисте 2 курс. 1. Понятие прогенеза и эмбриогенеза. Периоды и основные стадии эмбриогенеза у человека. Половые клетки человека, их структурногенетическая характеристика
 Скачать 2.3 Mb. Скачать 2.3 Mb.
|

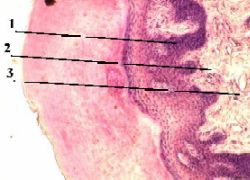
Межклеточные соединения (контакты)Плазмолемма многоклеточных животных организмов принимает активное участие в образовании специальных структур — межклеточных соединений (junctiones intercellulares), обеспечивающих межклеточные взаимодействия. Различают несколько типов таких структур (рис. 7). Простое межклеточное соединение, (junctio intercellularis simplex) — сближение плазмолемм соседних клеток на расстояние 15—20 нм. При этом происходит взаимодействие слоев гликокаликса соседних клеток. Разновидностью простого соединения является "пальцевидное", или соединение по типу замка. Плотное соединение (запирающая зона) (zonula occludens) — зона, где слои двух плазмолемм максимально сближены, здесь происходит как бы слияние участков плазмолемм двух соседних клеток. Роль плотного замыкающего соединения заключается в механическом соединении клеток друг с другом. Эта область непроницаема для макромолекул и ионов и, следовательно, она запирает, отграничивает межклеточные щели (и вместе с ними собственно внутреннюю среду организма) от внешней среды. Часто встречается, особенно в эпителии, особый тип соединения — пятно сцепления, или десмосома (desmosoma). Эта структура представляет собой небольшую площадку, иногда имеющую слоистый вид, диаметром до 0,5 мкм, где между мембранами располагается зона с высокой электронной плотностью. К плазмолемме в зоне десмосомы со стороны цитоплазмы прилегает участок электронноплотного вещества, так что внутренний слой мембраны кажется утолщенным. Под этим утолщением находится область тонких фибрилл, которые могут быть погружены в относительно плотный матрикс. Функциональная роль десмосом заключается главным образом в механической связи между клетками. Щелевидное соединение, или нексус (nexus), представляет собой область протяженностью 0,5—3 мкм, где плазмолеммы разделены промежутком в 2—3 нм. Со стороны цитоплазмы никаких специальных примембранных структур в данной области не обнаруживается, но в структуре плазмолемм соседних клеток друг против друга располагаются специальные белковые комплексы (коннексоны), которые образуют как бы каналы из одной клетки в другую. Этот тип соединения встречается во всех группах тканей. Функциональная роль щелевидного соединения заключается в переносе ионов и мелких молекул от клетки к клетке. Так, в сердечной мышце возбуждение, в основе которого лежит процесс изменения ионной проницаемости, передается от клетки к клетке через нексус. Синаптические соединения, или синапсы (synapsis). Этот тип соединений характерен для нервной ткани и встречается в специализированных участках контакта как между двумя нейронами, так и между нейроном и каким-либо иным элементом, входящим в состав рецептора или эффектора (например, нервно-мышечные, нервно-эпителиальные синапсы). Синапсы — участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому. Реснички — органоиды, аналогичные по строению и функцию с центриолями, т.е. имеют сходное строение и обеспечивают двигательную функцию. Ресничка представляет собой вырост цитоплазмы на поверхности клетки, покрытый цитолеммой. Вдоль этого выроста внутри располагаются 9 пар микротрубочек, расположенных параллельно друг к другу, образуя цилиндр; в центре этого цилиндра вдоль, а следовательно и в центре реснички, располагается еще 1 пара центральных микротрубочек. У основания этого выроста-реснички, перпендикулярно к ней, располагается еще одна аналогичная структура. 8.Микроворсинки — это выросты цитоплазмы на поверхности клеток, покрыты снаружи цитолеммой, увеличивают площадь поверхности клетки. Встречаются в эпителиальных клетках, обеспечивающих функцию всасывания (кишечник, почечные канальцы). 13. Органеллы цитоплазмы, понятие и классификация. Структурно-функциональная характеристика органелл, участвующих в биосинтезе внутриклеточных веществ и внутриклеточных защитных реакциях. Органоиды — постоянные структуры цитоплазмы, имеющие определенное строение и функции. Органоиды классифицируются по строению и по функцию. По строению различают: 1. Органоиды общего назначения (имеются в большем или меньшем количестве во всех клетках, обеспечивают функции необходимые всем клеткам): митохондрия, эндоплазматическая сеть, пластинчатый комплекс, лизосомы, клеточный центр, пероксисомы. 2. Органоиды специального назначения — (имеются только в клетках высокоспециализированных тканей и обеспечивают выполнение строгоспецифических функций этих тканей): в эпителиальных клетках — реснички, микроворсинки, тонофибриллы; в нейральных тканях — нейрофибриллы и базофильное вещество; в мышечных тканях — миофибриллы. По строению органиоды подразделяются: 1. Мембранные — эндоплазматическая сеть, митохондрии, пластинчатый комплекс, лизосомы, пероксисомы. 2. Немембранные — рибосомы, микротрубочки, центриоли, реснички. Строение и функции органоидов: 1. Митохондрии — структуры округлой, овальной и сильновытянутой эллепсоидной формы. Окружены двойной элементарной мембраной: наружная элементарная мембрана имеет ровную поверхность, внутренняя мембрана образует складки — кристы; полость внутри внутренней мембраны заполнена матриксом — гомогенная бесструктурная масса. Функция: митохондрии называют»энергетическими станциями» клетки, т.е. там происходит аккумулирование энергии в виде АТФ, выделяемое при «сжигании» белков, жиров, углеводов и др. веществ. Короче, митохондрии — поставщики энергии. 2. Эндоплазматическая сеть(ЭПС) — это система (сеть) внутриклеточных канальцев, стенки которых состоит из элементантарных биологических мембран. Различают ЭПС гранулярного типа (в стенки ЭПС вмонтированы гранулы = рибосомы) — с фукнцией синтеза белков, и агранулярного типа (канальцы без рибосом) — с функцией синтеза жиров, липидов и углеводов. 3. Пластинчатый комплекс (Гольджи) — система наслоенных друг на друга уплощенных цистерн, стенка которых состоит из элементарной биологической мембраны, и расположенных рядом пузырьков (везикул). Располагается обычно над ядром, и выполняет функцию — завершение процессов синтеза веществ в клетке, расфасовка продуктов синтеза по порциям в везикулы, ограниченных элементарной биологической мембраной. Везикулы в дальнейшем транспортируются в пределах данной клетки или выводятся экзоцитолизом за пределы клетки. 4.Лизосомы — структуры округлой или овальной формы, окружены элементарной биологической мембраной, содержащие внутри полный комплект протеолитических и других литических ферментов. Функция — обеспечивают внутриклеточное переваривание, т.е. последнюю фазу фаго(пино)цитоза. 5.Пироксисомы — мелкие структуры округлой или овальной формы, окруженные элементарной базальной мембраной, содержащие внутри пероксидазу, обеспечивающая обезвреживание перекисных радикалов — продуктов обмена веществ, подлежащих удалению из организма. 6.Клеточный центр — органоид обеспечивающий двигательную функцию (растаскивание хромосом) при делении клетки. Состоит из 2-х центриолей; каждая центриоля представляет собой цилиндрическое тело, стенка которого образована 9-ю парами микротрубочек расположенных по периферии цилиндра вдоль и 1-й парой микротрубочек в центре. Центриоли располагаются по отношению друг к другу перпендикулярно. При делении клетки центриоли располагаются на двух противоположных полюсах и обеспечивают растаскивание хромосом к полюсам. 14. Структурно-функциональная характеристика органелл, участвующих в процессах выведения веществ из клеток, во внутриклеточном пищеварении, защитных и обезвреживающих реакциях и энергопроизводстве. Строение и функции органоидов: 1. Митохондрии — структуры округлой, овальной и сильновытянутой эллепсоидной формы. Окружены двойной элементарной мембраной: наружная элементарная мембрана имеет ровную поверхность, внутренняя мембрана образует складки — кристы; полость внутри внутренней мембраны заполнена матриксом — гомогенная бесструктурная масса. Функция: митохондрии называют»энергетическими станциями» клетки, т.е. там происходит аккумулирование энергии в виде АТФ, выделяемое при «сжигании» белков, жиров, углеводов и др. веществ. Короче, митохондрии — поставщики энергии. 2. Эндоплазматическая сеть(ЭПС) — это система (сеть) внутриклеточных канальцев, стенки которых состоит из элементантарных биологических мембран. Различают ЭПС гранулярного типа (в стенки ЭПС вмонтированы гранулы = рибосомы) — с фукнцией синтеза белков, и агранулярного типа (канальцы без рибосом) — с функцией синтеза жиров, липидов и углеводов. 3. Пластинчатый комплекс (Гольджи) — система наслоенных друг на друга уплощенных цистерн, стенка которых состоит из элементарной биологической мембраны, и расположенных рядом пузырьков (везикул). Располагается обычно над ядром, и выполняет функцию — завершение процессов синтеза веществ в клетке, расфасовка продуктов синтеза по порциям в везикулы, ограниченных элементарной биологической мембраной. Везикулы в дальнейшем транспортируются в пределах данной клетки или выводятся экзоцитолизом за пределы клетки. 4.Лизосомы — структуры округлой или овальной формы, окружены элементарной биологической мембраной, содержащие внутри полный комплект протеолитических и других литических ферментов. Функция — обеспечивают внутриклеточное переваривание, т.е. последнюю фазу фаго(пино)цитоза. 5.Пироксисомы — мелкие структуры округлой или овальной формы, окруженные элементарной базальной мембраной, содержащие внутри пероксидазу, обеспечивающая обезвреживание перекисных радикалов — продуктов обмена веществ, подлежащих удалению из организма. 12. Пероксисомы — овальные тельца (0,5-1,5 мкм) окруженные элементарной мембраной, заполненные гранулярным матриксом с кристаллоподобными структурами; содержат каталазы для разрушения перекисных радикалов. Функция: обезвреживание перекисных радикалов, образующихся при метаболизме в клетках. Включения — непостоянные структуры цитоплазмы, могущие появляться или исчезать, в зависимости от функционального состояния клетки. Классификация включений: I. Трофические включения — отложенные в запас гранулы питательных веществ (белки, жиры, углеводы). В качестве примеров можно привести: гликоген в нейтрофильных гранулоцитах, в гепатоцитах, в мышечных волокнах; жировые капельки в гепатоцитах и липоцитах; белковые гранулы в составе желтка яйцеклеток и т. д. II. Пигментные включения — гранулы эндогенных или экзогенных пигментов. Примеры: меланин в меланоцитах кожи (для защиты от УФЛ), гемаглобин в эритроцитах (для транпортировки кислорода и углекислого газа), родопсин и йодопсин в палочках и колбочках сетчатки глаза (обеспечивают черно-белое и цветное зрение) и т.д. III. Секреторные включения — капельки (гранулы) секрета веществ, подготовленные для выделения из любых секреторных клеток (в клетках всех экзокринных и эндокринных желез). Пример: капельки молока в лактоцитах, зимогенные гранулы в панкреатоцитах и т.д. IV. Экскреторные включения — конечные (вредные) продукты обмена веществ, подлежащие удалению из организма. Пример: включения мочевины, мочевой кислоты, креатинина в эпителиоцитах почечных канальцев 15. Структурно-функциональная и химическая характеристика органелл, составляющих цитоскелет клетки, строение и значение центриолей, ресничек и жгутиков. Клеточный центр — органоид обеспечивающий двигательную функцию (растаскивание хромосом) при делении клетки. Состоит из 2-х центриолей; каждая центриоля представляет собой цилиндрическое тело, стенка которого образована 9-ю парами микротрубочек расположенных по периферии цилиндра вдоль и 1-й парой микротрубочек в центре. Центриоли располагаются по отношению друг к другу перпендикулярно. При делении клетки центриоли располагаются на двух противоположных полюсах и обеспечивают растаскивание хромосом к полюсам. 7.Реснички — органоиды, аналогичные по строению и функцию с центриолями, т.е. имеют сходное строение и обеспечивают двигательную функцию. Ресничка представляет собой вырост цитоплазмы на поверхности клетки, покрытый цитолеммой. Вдоль этого выроста внутри располагаются 9 пар микротрубочек, расположенных параллельно друг к другу, образуя цилиндр; в центре этого цилиндра вдоль, а следовательно и в центре реснички, располагается еще 1 пара центральных микротрубочек. У основания этого выроста-реснички, перпендикулярно к ней, располагается еще одна аналогичная структура. 8.Микроворсинки — это выросты цитоплазмы на поверхности клеток, покрыты снаружи цитолеммой, увеличивают площадь поверхности клетки. Встречаются в эпителиальных клетках, обеспечивающих функцию всасывания (кишечник, почечные канальцы). 9,Миофибриллы — состоят из сократительных белков актина и миозина, имеются в мышечных клетках и обеспечивают процесс сокращения. 10.Нейрофибриллы — встречаются в нейроцитах и представляют собой совокупность нейрофибрилл и нейротрубочек. В теле клетки располагаются беспорядочно, а в отростках — параллельно друг к другу. Выполняют функцию скелета нейроцитов (т.е. функция цитоскелета), а в отростках участвуют в транспортировке веществ от тела нейроцитов по отросткам на периферию.  16. Включения цитоплазмы: понятие, классификация, хим и морфофункциональная характеристика. Включения — непостоянные структуры цитоплазмы, могущие появляться или исчезать, в зависимости от функционального состояния клетки. Классификация включений: I. Трофические включения — отложенные в запас гранулы питательных веществ (белки, жиры, углеводы). В качестве примеров можно привести: гликоген в нейтрофильных гранулоцитах, в гепатоцитах, в мышечных волокнах; жировые капельки в гепатоцитах и липоцитах; белковые гранулы в составе желтка яйцеклеток и т. д. II. Пигментные включения — гранулы эндогенных или экзогенных пигментов. Примеры: меланин в меланоцитах кожи (для защиты от УФЛ), гемаглобин в эритроцитах (для транпортировки кислорода и углекислого газа), родопсин и йодопсин в палочках и колбочках сетчатки глаза (обеспечивают черно-белое и цветное зрение) и т.д. III. Секреторные включения — капельки (гранулы) секрета веществ, подготовленные для выделения из любых секреторных клеток (в клетках всех экзокринных и эндокринных желез). Пример: капельки молока в лактоцитах, зимогенные гранулы в панкреатоцитах и т.д. IV. Экскреторные включения — конечные (вредные) продукты обмена веществ, подлежащие удалению из организма. Пример: включения мочевины, мочевой кислоты, креатинина в эпителиоцитах почечных канальцев 17. Ядро клетки: строение, функции и хим состав. Взаимодействие структур ядра и цитоплазмы в процессе синтеза белка в клетках. Ядро — часть клетки, являющееся хранилищем наследственной информации. Окружено кариолеммой (два листка элементарной биомембраны), имеющей поры. В ядре содержится кариоплазма, основу которой составляет ядерный белковый матрикс (структурная сеть из негистоновых белков). В в ядерном белковом матриксе располагается хроматин — ДНК в комплексе с гистоновыми и негистоновыми белками. Хроматин может быть деконденцированным (разрыхленным, светлым) — эухроматин («эу»- хороший) и наоборот, конденсированным (плотно упакованным, темным) — гетерохроматин. Чем больше эухроматина, тем интенсивнее синтетические процессы в ядре и цитоплазме, и наоборот, преобладание гетерохроматина показывает на снижение синтетических процессов, на состояние метаболического покоя. Ядрышко — самая плотная, интенсивно окрашивающаяся структура ядра с диаметром 1-5 мкм — является производным хроматина, одним из его локусов. Функция: образование рРНК и рибосом. ЯДЕРНО-ЦИТОПЛАЗМАТИЧЕСКОЕ ВЗАИМОДЕЙСТВИЕ процессы взаимодействия ядра и цитоплазмы, обеспечивающие морфолого-функц. единство клетки. Под действием входящих из цитоплазмы в ядро регуляторов активности генов (обычно белков) происходит активация или же инактивация транскрипции тех или иных ядерных генов. В ядро поступают также предшественники и ферменты, необходимые для репликации ДНК, синтеза РНК, а также белки, входящие в состав хроматина, ядрышек и др. структур ядра. У простейших и нек-рых низших растений перед митозом в ядро поступают тубулины — белки, из к-рых строятся микротрубочки митотич. веретена. Из ядра в цитоплазму, вероятно, через поры, выходят продукты генной активности — разл. формы РНК и РНП, к-рые в дальнейшем обеспечивают синтез белка в цитоплазме и определяют его специфичность. Т. о., ядро управляет всеми белковыми синтезами и через них физиол. и морфологич. процессами в клетке, а цитоплазма регулирует (по принципу обратной связи) активность генетич. аппарата ядра и снабжает его материалами и энергией. В более широком смысле слова к Я.-ц. в. относятся также взаимодействия геномов ядра и митохондрий, ядра и пластид (межгеномные взаимодействия). Осн. метод изучения Я.-ц. в. — получение ядерно-цитоплазматич. гибридов путём пересадки ядер или слияния клеток. 18. Репродукция клеток и клеточных структур: способы репродукции, их характеристика, значение для ж/д организма.      19. Понятие о жизненном цикле клетки: его этапы и порфо-фун характеристика. Особенности жизненного цикла у различных видов клеток.      20. Неклеточные структуры организма: их моро-фун хар-ка, взаимоотношение клеток и неклеточных структур. Неклеточные структуры
2.3.1. Волокна соединительной ткани
2.3.2. Аморфное вещество хряща
21. Уровни организации живого. Определение ткани. Вклад Заварзина, Хлопина в учение о тканях. Классификация тканей. Структурные элементы тканей. Хар-ка симпластов и межклеточного вещества. Регенерация и изменчивость тканей. Ткани - это исторически (филогенетически) сложившиеся системы клеток и неклеточных структур, обладающих общностью строения, в ряде случаев - общностью происхождения, и специализированные на выполнении определенных функций. В любой системе все ее элементы упорядочены в пространстве и функционируют согласованно друг с другом; система в целом обладает при этом свойствами, не присущими ни одному из ее элементов, взятому в отдельности. Соответственно и в каждой ткани ее строение и функции несводимы к простой сумме свойств отдельных входящих в нее клеток. Ведущими элементами тканевой системы являются клетки (см. лекцию). Кроме клеток, различают клеточные производные и межклеточное вещество. К производным клеток относят симпласты (например, мышечные волокна, наружная часть трофобласта), синцитий (развивающиеся мужские половые клетки, пульпа эмалевого органа), а также постклеточные структуры (эритроциты, тромбоциты, роговые чешуйки эпидермиса и т. д.). Межклеточное вещество подразделяют на основное вещество и на волокна. Оно может быть представлено золем, гелем или быть минерализованным. Среди волокон различают обычно три вида: коллагеновые, ретикулярные, эластические Последовательная ступенчатая детерминация и коммитирование потенций однородных клеточных группировок — дивергентный процесс. В общем виде эволюционная концепция дивергентного развития тканей в филогенезе и в онтогенезе была сформулирована Н.Г.Хлопиным. Современные генетические концепции подтверждают правоту его представлений. Именно Н.Г.Хлопин ввел понятие о генетических тканевых типах. Концепция Хлопина хорошо отвечает на вопрос, как и какими путями происходило развитие и становление тканей, но не останавливается на причинах, определяющих пути развития. Причинные аспекты развития тканей раскрывает теория параллелизмов А.А.Заварзина. Он обратил внимание на сходство строения тканей, которые выполняют одинаковые функции у животных, принадлежащих даже к весьма удаленным друг от друга эволюционным группировкам. Вместе с тем известно, что, когда эволюционные ветви только расходились, у общих предков таких специализированных тканей еще не было. Следовательно, в ходе эволюции в разных ветвях филогенетического древа самостоятельно, как бы параллельно, возникали одинаково организованные ткани, выполняющие сходную функцию. Причиной этого является естественный отбор: если возникали какие-то организмы, у которых соответствие строения и функции клеток, тканей, органов нарушалось, они были и менее жизнеспособны. Теория Заварзина отвечает на вопрос, почему развитие тканей шло тем, а не иным путем, раскрывает казуальные аспекты эволюции тканей. Концепции А.А.Заварзина и Н.Г.Хлопина, разработанные независимо одна от другой, дополняют друг друга и были объединены А.А.Брауном и В.П.Михайловым: сходные тканевые структуры возникали параллельно в ходе дивергентного развития. Развитие тканей в эмбриогенезе происходит в результате дифференцировки клеток. Под дифференцировкой понимают изменения в структуре клеток в результате их функциональной специализации, обусловленные активностью их генетического аппарата. Различают четыре основных периода дифференцировки клеток зародыша — оотипическую, бластомерную, зачатковую и тканевую дифференцировку. Проходя через эти периоды клетки зародыша образуют ткани (гистогенез). КЛАССИФИКАЦИЯ ТКАНЕЙИмеется несколько классификаций тканей. Наиболее распространенной является так называемая морфофункциональная классификация, по которой насчитывают четыре группы тканей:
К тканям внутренней среды относятся соединительные ткани, кровь и лимфа. Эпителиальные ткани характеризуются объединением клеток в пласты или тяжи. Через эти ткани совершается обмен веществ между организмом и внешней средой. Эпителиальные ткани выполняют функции защиты, всасывания и экскреции. Источниками формирования эпителиальных тканей являются все три зародышевых листка — эктодерма, мезодерма и энтодерма. Ткани внутренней среды (соединительные ткани, включая скелетные, кровь и лимфа) развиваются из так называемой эмбриональной соединительной ткани — мезенхимы. Ткани внутренней среды характеризуются наличием большого количества межклеточного вещества и содержат различные клетки. Они специализируются на выполнении трофической, пластической, опорной и защитной функциях. Мышечные ткани специализированны на выполнении функции движения. Они развивается в основном из мезодермы (поперечно исчерченная ткань) и мезенхимы (гладкая мышечная ткань). Нервная ткань развивается из эктодермы и специализируется на выполнении регуляторной функции - восприятии, проведении и передачи информации. РЕГЕНЕРАЦИЯ ТКАНЕЙЗнание основ кинетики клеточных популяций необходимо для понимания теории регенерации, т.е. восстановления структуры биологического объекта после ее разрушения. Соответственно уровням организации живого различают клеточную (или внутриклеточную), тканевую, органную регенерацию. Предметом общей гистологии является регенерация на тканевом уровне. Различают регенерацию физиологическую, которая совершается постоянно в здоровом организме, и репаративную — вследствие повреждения. У разных тканей возможности регенерации неодинаковы. В ряде тканей гибель клеток генетически запрограммирована и совершается постоянно (в многослойном ороговевающем эпителии кожи, в однослойном каемчатом эпителии тонкой кишки, в крови). За счет непрерывного размножения, в первую очередь полустволовых клеток-предшественников, количество клеток в популяции пополняется и постоянно находится в состоянии равновесия. Наряду с запрограммированной физиологической гибелью клеток во всех тканях происходит и незапрограммированная — от случайных причин: травмирования, интоксикаций, воздействий радиационного фона. Хотя в ряде тканей запрограммированной гибели нет, но в течение всей жизни в них сохраняются стволовые и полу-стволовые клетки. В ответ на случайную гибель возникает их размножение и популяция восстанавливается. У взрослого человека в тканях, где стволовых клеток не остается, регенерация на тканевом уровне невозможна, она происходит лишь на клеточном уровне. Органы и системы организма являются многотканевыми образованиями, в которых различные ткани тесно взаимосвязаны и взаимообусловлены при выполнении ряда характерных функций. В процессе эволюции у высших животных и человека возникли интегрирующие и регулирующие системы организма — нервная и эндокринная. Все многотканевые компоненты органов и систем организма находятся под контролем этих регулирующих систем и, таким образом, осуществляется высокая интеграция организма как единого целого. В эволюционном развитии животного мира с усложнением организации возрастала интегрирующая и регулирующая роль нервной системы, в том числе и в нервной регуляции деятельности эндокринных желез. 23. Классификация тканей. Структурные элементы тканей. Понятие о стволовых клетках, популяциях и дифферонах. КЛАССИФИКАЦИЯ ТКАНЕЙИмеется несколько классификаций тканей. Наиболее распространенной является так называемая морфофункциональная классификация, по которой насчитывают четыре группы тканей:
К тканям внутренней среды относятся соединительные ткани, кровь и лимфа. Эпителиальные ткани характеризуются объединением клеток в пласты или тяжи. Через эти ткани совершается обмен веществ между организмом и внешней средой. Эпителиальные ткани выполняют функции защиты, всасывания и экскреции. Источниками формирования эпителиальных тканей являются все три зародышевых листка — эктодерма, мезодерма и энтодерма. Ткани внутренней среды (соединительные ткани, включая скелетные, кровь и лимфа) развиваются из так называемой эмбриональной соединительной ткани — мезенхимы. Ткани внутренней среды характеризуются наличием большого количества межклеточного вещества и содержат различные клетки. Они специализируются на выполнении трофической, пластической, опорной и защитной функциях. Мышечные ткани специализированны на выполнении функции движения. Они развивается в основном из мезодермы (поперечно исчерченная ткань) и мезенхимы (гладкая мышечная ткань). Нервная ткань развивается из эктодермы и специализируется на выполнении регуляторной функции - восприятии, проведении и передачи информации. ОСНОВЫ КИНЕТИКИ КЛЕТОЧНЫХ ПОПУЛЯЦИЙКаждая ткань имеет или имела в эмбриогенезе стволовые клетки — наименее дифференцированные и наименее коммитированные. Они образуют самоподдерживающуюся популяцию, их потомки способны дифференцироваться в нескольких направлениях под влиянием микроокружения (факторов дифференцировки), образуя клетки-предшественники и, далее, функционирующие дифференцированные клетки. Таким образом, стволовые клетки полипотентны. Они делятся редко, пополнение зрелых клеток ткани, если это необходимо, осуществляется в первую очередь за счет клеток следующих генераций (клеток-предшественников). По сравнению со всеми другими клетками данной ткани стволовые клетки наиболее устойчивы к повреждающим воздействиям. Хотя в состав ткани входят не только клетки, именно клетки являются ведущими элементами системы, т. е. определяют ее основные свойства. Их разрушение приводит к деструкции системы и, как правило, их гибель делает ткань нежизнеспособной, особенно если были затронуты стволовые клетки. Если одна из стволовых клеток вступает на путь дифференциации, то в результате последовательного ряда коммитирующих митозов возникают сначала полустволовые, а затем и дифференцированные клетки со специфической функцией. Выход стволовой клетки из популяции служит сигналом для деления другой стволовой клетки по типу некоммитирующего митоза. Общая численность стволовых клеток в итоге восстанавливается. В условиях нормальной жизнедеятельности она сохраняется приблизительно постоянной. Совокупность клеток, развивающихся из одного вида стволовых клеток, составляет стволовой дифферон. Часто в образовании ткани участвуют различные диффероны. Так, в состав эпидермиса, кроме кератиноцитов, входят клетки, развивающиеся в нейральном гребне и имеющие другую детерминацию (меланоциты), а также клетки, развивающиеся путем дифференциации стволовой клетки крови, т. е. принадлежащие уже к третьему дифферону (внутриэпидерминальные макрофаги, или клетки Лангерганса). Дифференцированные клетки наряду с выполнением своих специфических функций способны синтезировать особые вещества — кейлоны, тормозящие интенсивность размножения клеток-предшественников и стволовых клеток. Если в силу каких-либо причин количество дифференцированных функционирующих клеток уменьшается (например, после травмы), тормозящее действие кейлонов ослабевает и численность популяции восстанавливается. Кроме кейлонов (местных регуляторов), клеточное размножение контролируется гормонами; одновременно продукты жизнедеятельности клеток регулируют активность желёз внутренней секреции. Если какие-либо клетки под воздействием внешних повреждающих факторов претерпевают мутации, они элиминируются из тканевой системы вследствие иммунологических реакций. Выбор пути дифференциации клеток определяется межклеточными взаимодействиями. Влияние микроокружения изменяет активность генома дифференцирующейся клетки, активируя одни и блокируя другие гены. У клеток, уже дифференцированных и утративших способность к дальнейшему размножению, строение и функция тоже могут изменяться (например, у гранулоцитов начиная со стадии метамиелоцита). Такой процесс не приводит к возникновению различий среди потомков клетки и для него больше подходит название «специализация». 24. Морфо-функ хар-ка эпителиальных тканей, источники их развития. Классификация. Вклад Хлопина в изучение эпителиальных тканей. Особенности строения эпител клеток, поляризация, специальные органеллы межклеточные соединения. Строение и роль базальной мембраны. НГ Хлопин создал теорию дивергентного развития тканей в фило- и онтогенезе т.е. объяснил как и какими путями происходило развитие и становление тканей. Соответственно этой теории Хлопин выдвинул генетическую классификацию тканей. Согласно Хлопину из 8 зачатков — энтодермы, целомической выстилки, энтомезенхимы, миотомов, хорды, кожной эктодермы, нейроэктодермы, прехордальной пластинки — в ходе дивергентной дифференцировки путем расхождения признаков образуются все виды тканей; поэтому в основу этой классификации Т положены источники развития. СИСТЕМА ЭПИТЕЛИАЛЬНЫХ ТКАНЕЙ. Эпителиальные ткани (ЭТ) в фило- и онтогенезе образуются первыми, т.е. древнейшяя гистологическая система. Для ЭТ характерны следующие отличительные свойства: 1. Пограничность — ЭТ покрывают наружные поверхности органов и внутренние поверхности полостей, т.е. разграничивают внутреннюю среду организма от окружающей среды и среды полостей. 2. Состоит только из клеток, межклеточное вещество практически отсутствует. 3. Клетки лежат плотно друг к другу, образуя сплошной пласт. 4. Эп. всегда располагается на базальной мембране (углеводнобелково- липидный комплекс с тончайшими фибриллами) и им отграничивается от подлежащей рыхлой соединительной ткани. 5. Эп. не имеет собственных кровеносных сосудов, питается диффузно через базальную мембрану, за счет сосудов подлежащей рыхлой соединительной ткани. 6. Эпителию характерно гетерополярность — апикальные (верхушка) и базальные части клеток отличаются по строению и по функции; а в многослойном эпителии — отличие в строении и функции слоев. 7. Характерно повышенная регенераторная способность, обусловленная пограничностью — чаще чем другие ткани подвергается воздействию неблагоприятных факторов и чаще гибнут клетки, отсюда необходимость в высокой регенераторной способности. 8. Эпителиоциты могут иметь органоиды специального назначения: - реснички (эпителий воздухоносных путей); - микроворсинки ( эпителий кишечника и почек); - тонофибриллы ( эпителий кожи). 9. Функции: - защитная; - разграничительная; - участие в обмене веществ между организмом и окружающей средой; - секреторная. КЛАССИФИКАЦИЯ. Для системы ЭТ используется 2 классификации — морфофункциональная (по строению и функции) и гистогенетическая (по происхождению или источникам развития). Гистогенетическая классификация: 1. Эп. кожного типа (эктодермальные) — многослойный плоский ороговевающий и неороговевающий эп.; эп. слюнных, сальных, молочных и потовых желез; переходный эпителий мочеиспускательного канала; многорядный мерцательный эп. воздухоносных путей; альвеолярный эп. легких; эп. щитовидной и паращитовидной железы, тимуса и аденогипофиза. 2. Эпителии кишечного типа (энтеродермальный) — однослойный призматический эп. кишечного тракта; эп. печени и поджелудочной железы. 3. Эпителий почечного типа (нефродермальный) — эпителий нефрона. 4. Эпителий целомического типа (целодермальный) — однослойный плоский эпителий серозных покровов (брюшины, плевры, околосердечной сумки); эп. половых желез; эп. коры надпочечников. 5. Эпителий нейроглиального типа — эпиндимный эп. мозговых желудочков; эп. мозговых оболочек; пигментный эп. сетчатки глаза; обонятельный эп.; глиальный эп. органа слуха; вкусовой эп.; эп. передней камеры глаза; хромофобный эп. мозгового слоя надпочечников; периневральный эпителий. Морфофункциональная классификация (применяется чаще): I. Однослойный эпителий. 1. Однослойный однорядный эпителий. а) однослойный плоский; б) однослойный кубический; в) однослойный цилиндрический (призматический): - однослойный призматический каемчатый - однослойный призматический железистый - однослойный призматический мерцательный 2. Однослойный многорядный мерцательный эпителий. II. Многослойный эпителий. 1. Многослойный плоский неороговевающий 2. Многослойный плоский ороговевающий 3. Переходный В однослойном эп. все клетки без исключения непосредственно связаны, (контактируют) с базальной мембраной. В однослойном однорядном эпителии все клетки контактируют с базальной мембраной; имеют одинаковую высоту, поэтому ядра располагаются на одном уровне. Однослойный плоский эпителий — состоит из одного слоя резко уплощенных клеток полигональной формы (многоугольной); основание (ширина) клеток больше, чем высота (толщина); в клетках органоидов мало, встречаются митохондрии, одиночные микроворсинки, в цитоплазме видны пиноцитозные пузырьки. Однослойный плоский эпителий выстилает серозные покровы (брюшина, плевра, околосердечная сумка). В отношении эндотелия (клетки выстилающие кровеносные и лимфатические сосуды, полости сердца) среди гистологов единого мнения нет: одни относят эндотелий однослойному плоскому эпителию, другие — к соединительной ткани со специальными свойствами. Источники развития: эндотелий развивается из мезенхимы; однослойный плоский эпителий серозных покровов — из спланхнотомов (вентральная часть мезодермы). Функции: разграничительная, уменьшает трение внутренних органов путем выделения серозной жидкости. Однослойный кубический эпителий — на срезе у клеток диаметр (ширина) равен высоте. Встречается в выводных протоках экзокринных желез, в извитых почечных канальцах. Однослойный призматический (цилиндрический) эпителий — на срезе ширина клеток меньше чем высота. В зависимости от особенностей строения и функции различают: - однослойный призматический железистый, имееется в желудке, в канале шейки матки, специализирован на непрерывную выработку слизи; - однослойный призматический каемчатый, выстилает кишечник, на апикальной поверхности клеток имеется большое количество микроворсинок; специализирован на всасывание. - однослойный призматический реснитчатый, выстилает маточные трубы; на апикальной поверхности эпителиоциты имеют реснички. Регенерация однослойного однорядного эпителия происходит за счет стволовых (камбиальных) клеток, равномерно разбросанных среди других дифференцированных клеток. Однослойный многорядный мерцательный эпителий — все клетки контактируют с базальной мембраной, но имеют разную высоту и поэтому ядра располагаются на разных уровнях, т.е. в несколько рядов. Выстилает воздухоносные пути. В составе этого эпителия различают разновидности клеток: - короткие и длинные вставочные клетки (малодифференцированные и среди них стволовые клетки; обеспечивают регенерацию); - бокаловидные клетки — имеют форму бокала, плохо воспринимают красители (в препарате — белые), вырабатывают слизь; - реснитчатые клетки, на апикальной поверхности имеют мерцательные реснички. Функция: очистка и увлажнение проходящего воздуха. Многослойный эпителий — состоит из нескольких слоев клеток, причем с базальной мембраной контактирует только самый нижний ряд клеток. 1. Многослойный плоский неороговевающий эпителий — выстилает передний (ротовая полость, глотка., пищевод) и конечный отдел (анальный отдел прямой кишки) пищеварительной системы, роговицу. Состоит из слоев: а) базальный слой — цилиндрической формы эпителиоциты со слабобазофильной цитоплазмой, часто с фигурой митоза; в небольшом количестве стволовые клетки для регенерации; б) шиповатый слой — состоит из значительного количества слоев клеток шиповатой формы ( ), клетки активно делятся. в) покровные клетки — плоские, стареющие клетки, не делятся, с поверхности постепенно слущиваются. Источник развития: эктодерма. Прехордальная пластинка в составе энтодермы передний кишки. Функция: механ. защита. 2. Многослойный плоский ороговевающий эпителий — это эпителий кожи. Развивается из эктодермы, выполняет защитную функцию — защита от механических повреждений, лучевого, бактериального и химического воздействия, разграничивает организм от окружающей среды. Состоит из слоев: а) базальный слой — во многом похож на аналогичный слой многослойного неороговевающего эпителия; дополнительно: содержит до 10% меланоцитов — отросчатые клетки с включениями меланина в цитоплазме — обеспечивают защиту от УФЛ; имеется небольшое количество клеток Меркеля (входят в состав механорецепторов); дендритические клетки с защитной функцией путем фагоцитоза; в эпителиоцитах содержатся тонофибриллы (органоид спец. назначения — обеспечивают прочность). б) шиповатый слой — из эпителиоцитов с шиповидными выростами; встречаются дендроциты и лимфоциты крови; эпителиоциты еще делятся. в) зернистый слой — из нескольких рядов вытянутых уплощенно-овальных клеток с базофильными гранулами кератогиалина (предшественник рогового вещества — кератина) в цитоплазме; клетки не делятся. г) блестящий слой — клетки полностью заполнены элаидином (образуется из кератина и продуктов распада тонофибрилл), отражающим и сильно преломляющим свет; под микроскопом границ клеток и ядер не видно. д) слой роговых чешуек — состоит из роговых пластинок из кератина, содержащих пузырьки с жиром и воздухом, кератосомы (соответствуют лизосомам). С поверхности чешуйки слущиваются. 3. Переходный эпителий — выстилает полые органы, стенка которых способна сильному растяжению (лоханка, мочеточники, мочевой пузырь). Слои: - базальный слой (из мелких темных низкопризматических или кубических клеток — малодифференцированные и стволовые клетки, обеспечивают регенерацию; - промежуточный слой — из крупных грушевидных клеток, узкой базальной частью, контактирующий с базальной мембраной (стенка не растянута, поэтому эпителий утолщен); когда стенка органа растянута грушевидные клетки уменьшаются по высоте и располагаются среди базальных клеток. - покровные клетки — крупные куполообразные клетки; при растянутой стенки органа клетки уплощаются; клетки не делятся, постепенно слущиваются. Таким образом, строение переходного эпителия изменяется в зависимости от состояния органа: когда стенка не растянута, эпителий утолщен за счет «вытеснения» части клеток из базального слоя в промежуточный слой; при растянутой стенки толщина эпителия уменьшается за счет уплощения покровных клеток и перехода части клеток из промежуточного слоя в базальный. Источники развития: эп. лоханки и мочеточника — из мезонефрального протока (производное сегментных ножек), эп. мочевого пузыря — из энтодермы аллантоиса и энтодермы клоаки. Функция — защитная. ЖЕЛЕЗИСТЫЙ ЭПИТЕЛИЙ Железистый эп. (ЖЭ) специализирован на выработку секрета. ЖЭ образует железы: I. Эндокринные железы — не имеют выводных протоков, секрет выделяется непосредственно в кровь или лимфу; обильно кровоснабжаются; вырабатывают гормоны или биологически активные вещества, оказывающие сильное регулирующее влияние на органы и системы даже в небольших дозах. II. Экзокринные железы — имеют выводные протоки, выделяют секрет на поверхность эпителия ( на наружные поверхности или в полости). Состоят из концевых (секреторных) отделов и выводных протоков. Принципы классификации экзокринных желез: I. По строению выводных протоков: 1. Простые — выводной проток не ветвится. 2. Сложные — выводной проток ветвится. II. По строению (форме) секреторных отделов: 1. Альвеолярные — секреторный отдел в виде альвеолы, пузырька. 2. Трубчатые — секр. отдел в виде трубочки. 3. Альвеолярно-трубчатые (смешанная форма). III. По соотношению выводных протоков и секреторных отделов: 1. Неразветвленные — в один выводной проток открывается один секретор- ный отдел. 2. Разветвленные — в один выводной проток открывается несколько секре- торных отделов. IV. По типу секреции: 1. Мерокриновые — при секреции целосность клеток не нарушается. Харак- терно для большинства желез. 2. Апокриновые (апекс — верхушка, кринио — выделение) — при секреции частично разрушается (отрывается) верхушка клеток (пр.: молочные железы). 3. Голокриновые — при секреции клетка полностью разрушается. Пр.: сальные железы кожи. V. По локализации: 1. Эндоэпителиальные — одноклеточная железа в толще покровного эпителия. Пр.: бокаловидные клетки в эпителие кишечника и воздухонос. путей. 2. Экзоэпителиальные железы — секреторный отдел лежит вне эпителия, в подлежащих тканях. VI. По характеру секрета: -белковые,слизистые, слизисто-белковые, потовые, сальные, молочные и т.д. Фазы секреции: 1. Поступление в железистые клетки исходных материалов для синтеза секрета (аминокислоты, липиды, минеральные вещества и т.д.). 2. Синтез (в ЭПС) и накопление (в ПК) в железистых клетках секрета. 3. Выделение секрета. Для клеток железистого эпителия характерно наличие органелл: ЭПС гранулярного или агранулярного типа (в зависимости от характера секрета), пластинчатый комплекс, митохондрии. Регенерация железистого эпителия — в большинстве железах регенерация железистого эпителия происходит путем деления малодифференцированных (камбиальных) клеток. Отдельные железы (слюнные железы, поджелудочная железа) стволовых и малодифференцированных клеток не имеют и в них происходит внутриклеточная регенерация — т.е. обновление внутри клеток изношенных органоидов, при отсутствии способности к делению клеток. Строение базальной мембраны
Тип контакта базальной мембраны с эпителием: полудесмосома — сходна по строению с десмосомой, но это соединение клеток с межклеточными структурами. Так в эпителиях линкерные гликопротеиды (интегрины) десмосомы взаимодействуют с белками базальной мембраны. Функция — механическая. Базальные мембраны делят на:
Функции базальной мембраны
25. Морфо-фун х-ка покровного эпителия: классификация. Многослойные эпителии:различные виды, источники их развития, диффероны кожного эпителия. Физиологическая регенерация, локализация камбиальных элементов. | |||||||||||