экзамен по гисте 2 курс. 1. Понятие прогенеза и эмбриогенеза. Периоды и основные стадии эмбриогенеза у человека. Половые клетки человека, их структурногенетическая характеристика
 Скачать 2.3 Mb. Скачать 2.3 Mb.
|

Поперечнополосатые мышечные тканиИмеется две основные разновидности поперечнополосатых (исчерченных) тканей — скелетная мышечная ткань и сердечная мышечная ткань. Скелетная мышечная тканьГистогенезИсточником развития элементов скелетной (соматической) поперечнополосатой мышечной ткани являются клетки миотомов — миобласты. Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтонных мышц. Другие клетки мигрируют из миотомов в мезенхиму. Они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела. В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные симпласты — мышечные трубочки (миотубы). В них происходит дифференцировка специальных органелл — миофибрилл. В это время в миотубах отмечается хорошо развитая гранулярная эндоплазматическая сеть. Миофибриллы сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра, напротив, из центральных отделов смещаются к периферии. Клеточные центры и микротрубочки при этом полностью исчезают. Гранулярная эндоплазматическая сеть редуцируется в значительной степени. Такие дефинитивные структуры называют миосимпластами. Клетки другой линии остаются самостоятельными и дифференцируются в миосателлитоциты (или миосателлиты). Эти клетки располагаются на поверхности миосимпластов. СтроениеОсновной структурной единицей скелетной мышечной ткани является мышечное волокно, состоящее из миосимпласта и миосателлитоцитов, покрытых общей базальной мембраной. Длина всего волокна может измеряться сантиметрами при толщине всего 50—100 мкм. Комплекс, состоящий из плазмолеммы миосимпласта и базальной мембраны, называют сарколеммой. Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под сарколеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч. У полюсов ядер располагаются органеллы общего значения — аппарат Гольджи и небольшие фрагменты гранулярной эндоплазматической сети. Миофибриллы заполняют основную часть миосимпласта и расположены продольно. Саркомер — это структурная единица миофибриллы. Каждая миофибрилла имеет поперечные темные и светлые диски, имеющие неодинаковое лучепреломление (анизотропные A-диски и изотропные I-диски). Каждая миофибрилла окружена продольно расположенными и анастомозирующими между собой петлями агранулярной эндоплазматической сети — саркоплазматической сети, или саркоплазматического ретикулума. Соседние саркомеры имеют общую пограничную структуру — Z-линию (или телофрагму). Она построена в виде сети из белковых фибриллярных молекул, среди которых существенную роль играет альфа-актинин. С этой сетью связаны концы тонких, актиновых, филаментов. От соседних Z-линий актиновые филаменты направляются к центру саркомера, но не доходят до его середины. Филаменты актина объединены с Z-линией и нитями миозина фибриллярными нерастяжимыми молекулами небулина. Посередине темного диска саркомера располагается сеть, построенная из миомезина. Она образует в сечении М-линию, или мезофрагму. В узлах этой М-линии закреплены концы толстых, миозиновых филаментов. Другие их концы направляются в сторону Z-линий и располагаются между филаментами актина, но до самих Z-линий тоже не доходят. Вместе с тем эти концы фиксированы по отношению к Z-линиям растяжимыми гигантскими белковыми молекулами титина. Молекулы миозина имеют длинный хвост и на его конце две головки. При повышении концентрации ионов кальция в области присоединения головок (в своеобразном шарнирном участке) молекула миозина изменяет свою конфигурацию. При этом (поскольку между миозиновыми филаментами расположены актиновые) головки миозина связываются с актином (при участии вспомогательных белков — тропомиозина и тропонина). Затем головка миозина наклоняется и тянет за собой актиновую молекулу в сторону М-линии. Z-линии сближаются, саркомер укорачивается. 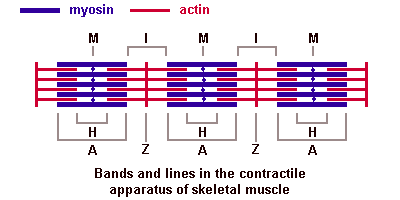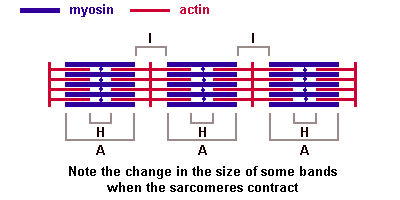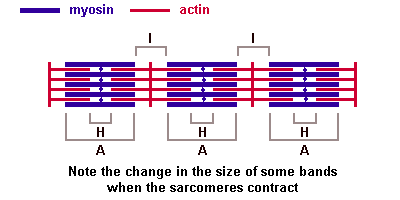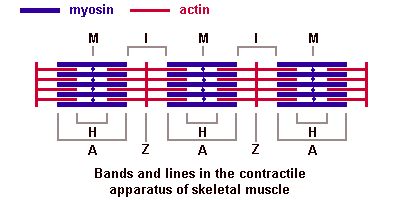 Альфа-актининовые сети Z-линий соседних миофибрилл связаны друг с другом промежуточными филаментами. Они подходят к внутренней поверхности плазмолеммы и закрепляются в кортикальном слое цитоплазмы, так что саркомеры всех миофибрилл располагаются на одном уровне. Это и создает при наблюдении в микроскоп впечатление поперечной исчерченности всего волокна. Источником ионов кальция служат цистерны агранулярной эндоплазматической сети. Они вытянуты вдоль миофибрилл около каждого саркомера и образуют саркоплазматическую сеть. Именно в ней аккумулируются ионы кальция, когда миосимпласт находится в расслабленном состоянии. На уровне Z-линий (у амфибии) или на границе А- и I-дисков (у млекопитающих) канальцы сети меняют направление и располагаются поперечно, образуя расширенные терминальные или (латеральные) L-цистерны. С поверхности миосимпласта плазмолемма образует длинные трубочки, идущие поперечно в глубину клетки (Т-трубочки) на уровне границ между темными и светлыми дисками. Когда клетка получает сигнал о начале сокращения, этот сигнал перемещается по плазмолемме в виде потенциала действия и распространяется отсюда на мембрану Т-трубочек. Поскольку эта мембрана сближена с мембранами саркоплазматической сети, состояние последних меняется, кальций освобождается из цистерн сети и взаимодействует с актино-миозиновыми комплексами (они сокращаются). Когда потенциал действия исчезает, кальций снова аккумулируется в цистернах саркоплазматического ретикулума и сокращение миофибрилл прекращается. Для развития усилия сокращения нужна энергия. Она освобождается за счет АТФ- АДФ-превращений. Роль АТФазы выполняет миозин. Источником АТФ служат главным образом митохондрии, поэтому они и располагаются непосредственно между миофибриллами. Большую роль в деятельности миосимпластов играют включения миоглобина и гликогена.Гликоген служит источником энергии, необходимой не только для совершения мышечной работы, но и поддержания теплового баланса всего организма. Миоглобин связывает кислород, когда мышца расслаблена и через мелкие кровеносные сосуды свободно протекает кровь. Во время сокращения мышцы сосуды сдавливаются, а запасенный кислород освобождается из миоглобина и участвует в биохимических реакциях. Миосателлитоциты - это малодифференцированные клетки, являющиеся источником регенерации мышечной ткани. Они прилежат к поверхности миосимпласта, так что их плазмолеммы соприкасаются. Миосателлитоциты одноядерны, их ядра овальной формы и мельче, чем в симпластах. Они обладают всеми органеллами общего значения (в том числе и клеточным центром). Типы мышечных волокон. Разные мышцы (как органы) функционируют в неодинаковых биомеханических условиях. Поэтому и мышечные волокна в составе разных мышц обладают разной силой, скоростью и длительностью сокращения, а также утомляемостью. Ферменты в них обладают разной активностью и представлены в различных изомерных формах. Заметно различие в них содержания дыхательных ферментов — гликолитических и окислительных. По соотношению миофибрилл, митохондрий и миоглобина различают белые, красные и промежуточные волокна. По функциональным особенностям мышечные волокна подразделяют на быстрые, медленные и промежуточные. Наиболее заметно мышечные волокна различаются особенностями молекулярной организации миозина. Среди различных его изоформ существуют две основных — «быстрая» и «медленная». При постановке гистохимических реакций их различают по АТФазной активности. С этими свойствами коррелирует и активность дыхательных ферментов. Обычно в быстрых волокнах преобладают гликолитические процессы, они более богаты гликогеном, в них меньше миоглобина, поэтому их называют также белыми. В медленных волокнах, напротив, выше активность окислительных ферментов, они богаче миоглобином, выглядят более красными. Свойства мышечных волокон меняются при изменении нагрузок — спортивных, профессиональных, а также в экстремальных условиях (таких как невесомость). При возврате к обычной деятельности такие изменения обратимы. При некоторых заболеваниях (мышечные атрофии, дистрофии, последствия денервации) мышечные волокна с разными исходными свойствами изменяются неодинаково. Это позволяет уточнять диагноз, для чего исследуют биоптаты скелетных мышц. Регенерация скелетной мышечной тканиЯдра миосимпластов делиться не могут, так как у них отсутствуют клеточные центры. Камбиальными элементами служат миосателлитоциты. Пока организм растет, они делятся, а дочерние клетки встраиваются в концы симпластов. По окончании роста размножение миосателлитоцитов затухает. После повреждения мышечного волокна на некотором протяжении от места травмы оно разрушается и его фрагменты фагоцитируются макрофагами. Восстановление любых тканей организма может осуществляется за счет двух механизмов: гипертрофии и гиперплазии. Под гипертрофией подразумевают компенсаторное увеличение объема самого симпласта, в т.ч. за счет увеличения количества миофибрилл. В симпласте активизируются гранулярная эндоплазматическая сеть и аппарат Гольджи. Происходит синтез веществ, необходимых для восстановления саркоплазмы и миофибрилл, а также сборка мембран, так что восстанавливается целостность плазмолеммы. Поврежденный конец миосимпласта при этом утолщается, образуя мышечную почку. Под гиперплазией понимают пролиферацию миосателлитоцитов. Сохранившиеся рядом с повреждением миосателлитоциты делятся. Одни из них мигрируют к мышечной почке и встраиваются в нее, другие сливаются (так же, как миобласты при гистогенезе) и образуют миотубы, которые затем входят в состав вновь образованных мышечных волокон или формируют новые волокна. 40. Мышца как орган: строение, васкуляризация, эф и аф иннервация, связь мышцы с сухожилием. Скелетная мышца как органПередача усилий сокращения на скелет осуществляется посредством сухожилий или прикрепления мышц непосредственно к надкостнице. На конце каждого мышечного волокна плазмолемма образует глубокие узкие впячивания. В них со стороны сухожилия или надкостницы проникают тонкие коллагеновые волокна. Последние спирально оплетаются ретикулярными волокнами. Концы волокон направляются к базальной мембране, входят в нее, поворачивают назад и по выходе снова оплетают коллагеновые волокна соединительной ткани. Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистойсоединительной ткани — эндомизий. Коллагеновые волокна наружного листка базальной мембраны вплетаются в него, что способствует объединению усилий при сокращении миосимпластов. Более толстые прослойки рыхлой соединительной ткани окружают по нескольку мышечных волокон, образуя перимизий и разделяя мышцу на пучки. Несколько пучков объединяются в более крупные группы, разделенные более толстыми соединительнотканными прослойками. Соединительную ткань, окружающую поверхность мышцы, называют эпимизием. Васкуляризация. Артерии вступают в мышцу и распространяются по прослойкам соединительной ткани, постепенно истончаясь. Ветви 5—6-го порядка образуют в перимизии артериолы. В эндомизии расположены капилляры. Они идут вдоль мышечных волокон, анастомозируя друг с другом. Венулы, вены и лимфатические сосуды проходят рядом с приносящими сосудами. Как обычно, рядом с сосудами много тканевых базофилов, принимающих участие в регуляции проницаемости сосудистой стенки. Иннервация. В мышцах выявлены миелинизированные эфферентные (двигательные), афферентные (чувствительные), а также немиелинизированные вегетативные нервные волокна. Отросток нервной клетки, приносящий импульс от моторного нейрона спинного мозга, ветвится в перимизии. Каждая его ветвь проникает сквозь базальную мембрану, и у поверхности симпласта на плазмолемме образует терминали, участвуя в организации так называемой моторной бляшки, или нервно-мышечного соединения. При поступлении нервного импульса из терминали выделяется ацетилхолин — медиатор, который вызывает возбуждающий потенциал действия, распространяющееся отсюда по плазмолемме миосимпласта. Итак, каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров, образуя комплекс, именуемый мионом. Группа же мышечных волокон, иннервируемых одним мотонейроном, называется нервно-мышечной единицей. Характерно, что мышечные волокна, принадлежащие к одной нервно-мышечной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам. Чувствительные нервные окончания располагаются не на рабочих мышечных волокнах, а связаны со специализированными мышечными волокнами в так называемых мышечных веретенах, которые расположены в перимизии. Волокна в таких чувствительных мышечных веретенах именуются интрафузальными волокнами, а обычные рабочие мышечные волокна – экстрафузальными. Интрафузальные мышечные волокна веретен значительно тоньше рабочих. Существует два их вида — волокна с ядерной сумкой и волокна с ядерной цепочкой. Каждое мышечное волокно веретена спирально обвито терминалью чувствительного нервного волокна. В результате сокращения или расслабления рабочих мышечных волокон изменяется натяжение соединительнотканной капсулы веретена, соответственно изменяется тонус интрафузальных мышечных волокон. Вследствие этого возбуждаются чувствительные нервные окончания, обвивающие их, и в области терминалей возникают афферентные нервные импульсы. На каждом миосимпласте располагается также своя моторная бляшка. Поэтому интрафузальные мышечные волокна постоянно находятся в напряжении, подстраиваясь к длине мышечного брюшка в целом. 41. Сердечная мышечная ткань. Регенерация. Сердечная мышечная тканьГистогенез и виды клеток. Источники развития сердечной поперечнополосатой мышечной ткани — симметричные участки висцерального листка спланхнотома в шейной части зародыша — так называемые миоэпикардиалъные пластинки. Из них дифференцируются также клетки мезотелия эпикарда. В ходе гистогенеза возникает 3 вида кардиомиоцитов:
Рабочие (сократительные) кардиомиоциты образуют свои цепочки. Укорачиваясь, они обеспечивают силу сокращения всей сердечной мышцы. Рабочие кардиомиоциты способны передавать управляющие сигналы друг другу. Синусные (пейсмекерные) кардиомиоциты способны автоматически в определенном ритме сменять состояние сокращения на состояние расслабления. Они воспринимают управляющие сигналы от нервных волокон, в ответ на что изменяют ритм сократительной деятельности. Синусные (пейсмекерные) кардиомиоциты передают управляющие сигналы переходным кардиомиоцитам, а последние — проводящим. Проводящие кардиомиоциты образуют цепочки клеток, соединенных своими концами. Первая клетка в цепочке воспринимает управляющие сигналы от синусных кардиомиоцитов и передает их далее — другим проводящим кардиомиоцитам. Клетки, замыкающие цепочку, передают сигнал через переходные кардиомиоциты рабочим. Секреторные кардиомиоциты выполняют особую функцию. Они вырабатывают гормон -натрийуретический фактор, участвующий в процессах регуляции мочеобразования и в некоторых других процессах. Сократительные кардиомиоциты имеют удлиненную (100—150 мкм) форму, близкую к цилиндрической. Их концы соединяются друг с другом, так что цепочки клеток составляют так называемые функциональные волокна (толщиной до 20 мкм). В области контактов клеток образуются так называемые вставочные диски. Кардиомиоциты могут ветвиться и образуют трехмерную сеть. Их поверхности покрыты базальной мембраной, в которую снаружи вплетаются ретикулярные и коллагеновые волокна. Ядро кардиомиоцита (иногда их два) овальное и лежит в центральной части клетки. У полюсов ядра сосредоточены немногочисленные органеллы общего значения. Миофибриллы слабо обособлены друг от друга, могут расщепляться. Их строение аналогично строению миофибрилл миосимпласта скелетного мышечного волокна. От поверхности плазмолеммы в глубь кардиомиоцита направлены Т-трубочки, находящиеся на уровне Z-линии. Их мембраны сближены, контактируют с мембранами гладкой эндоплазматической (т.е. саркоплазматической) сети. Петли последней вытянуты вдоль поверхности миофибрилл и имеют латеральные утолщения (L-системы), формирующие вместе с Т-трубочками триады или диады. В цитоплазме имеются включения гликогена и липидов, особенно много включений миоглобина. Механизм сокращения кардиомиоцитов такой же, как у миосимпласта. Кардиомиоциты соединяются друг с другом своими торцевыми концами. Здесь образуются так называемые вставочные диски: эти участки выглядят как тонкие пластинки при увеличении светового микроскопа. Фактически же концы кардиомиоцитов имеют неровную поверхность, поэтому выступы одной клетки входят во впадины другой. Поперечные участки выступов соседних клеток соединены друг с другом интердигитациями и десмосомами. К каждой десмосоме со стороны цитоплазмы подходит миофибрилла, закрепляющаяся концом в десмоплакиновом комплексе. Таким образом, при сокращении тяга одного кардиомиоцита передается другому. Боковые поверхности выступов кардиомиоцитов объединяются нексусами (или щелевыми соединениями). Это создает между ними метаболические связи и обеспечивает синхронность сокращений. Возможности регенерации сердечной мышечной ткани. При длительной усиленной работе (например, в условиях постоянно повышенного артериального давления крови) происходит рабочая гипертрофия кардиомиоцитов. Стволовых клеток или клеток-предшественников в сердечной мышечной ткани не обнаружено, поэтому погибающие кардиомиоциты (в частности, при инфаркте миокарда) не восстанавливаются, а замещаются элементами соединительной ткани. 42. Хар-ка нервной ткани. Источники развития. Нейроциты: функции, строение, морфолог и функциональн классификация. Нервная ткань — это система взаимосвязанных нервных клеток и нейроглии, обеспечивающих специфические функции восприятия раздражений, возбуждения, выработки импульса и его передачи. Она является основой строения органов нервной системы, обеспечивающих регуляцию всех тканей и органов, их интеграцию в организме и связь с окружающей средой. В нервной ткани выделяют два типа клеток – нервные и глиальные. Нервные клетки(нейроны, или нейроциты) — основные структурные компоненты нервной ткани, выполняющие специфическую функцию. Нейроглия обеспечивает существование и функционирование нервных клеток, осуществляя опорную, трофическую, разграничительную, секреторную и защитную функции. Развитие Нейруляция (схема). А — стадия нервной пластинки; Б - стадия нервного желобка; В - стадия нервной трубки. 1 - нервный желобок; 2 - нервный валик; 3 - кожная эктодерма; 4 - хорда; 5 - сомитная мезодерма; 6 - нервный гребень (ганглиозная пластинка); 7 - нервная трубка; 8 - мезенхима; 9 - эндодерма. 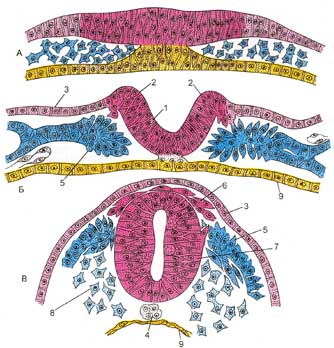 Нервная ткань развивается из дорсальной эктодермы. У 18-дневного эмбриона человека эктодерма по средней линии спины дифференцируется и утолщается, формируя нервную пластинку, латеральные края которой приподнимаются, образуя нервные валики, а между валиками формируется нервный желобок. Передний конец нервной пластинки расширяется, образуя позднее головной мозг. Латеральные края продолжают подниматься и растут медиально, пока не встретятся и не сольются по средней линии в нервную трубку, которая отделяется от лежащей над ней кожной эктодермы. Полость нервной трубки сохраняется у взрослых в виде системы желудочков головного мозга и центрального канала спинного мозга. Часть клеток нервной пластинки не входит в состав ни нервной трубки, ни кожной эктодермы, а образует скопления по бокам от нервной трубки, которые сливаются в рыхлый тяж, располагающийся между нервной трубкой и кожной эктодермой, — этонервный гребень (или ганглиозная пластинка). Из нервной трубки в дальнейшем формируются нейроны и макроглия центральной нервной системы. Нервный гребень дает начало нейронам чувствительных и автономных ганглиев, клеткам мягкой мозговой и паутинной оболочек мозга и некоторым видам глии: нейролеммоцитам (шванновским клеткам), клеткам-сателлитам ганглиев. Из нервного гребня развиваются также клетки мозгового вещества надпочечников, меланоциты кожи, часть клеток APUD-системы, сенсорные клетки каротидных телец. В формировании ганглиев V, VII, IX и X пар черепных нервов принимают участие, кроме нервного гребня, также нейрогенные плакоды, представляющие собой утолщения эктодермы по бокам формирующейся нервной трубки в краниальном отделе зародыша. Нервная трубка на ранних стадиях эмбриогенеза представляет собой многорядный нейроэпителий, состоящий из вентрикулярных, или нейроэпителиальных клеток. В дальнейшем в нервной трубке дифференцируется 4 концентрических зоны:
Вентрикулярная (эпендимная), внутренняя, зона состоит из делящихся клеток цилиндрической формы. Вентрикулярные (или матричные) клетки являются предшественниками нейронов и клеток макроглии. Субвентрикулярная зона состоит из клеток, сохраняющих высокую пролиферативную активность и являющихся потомками матричных клеток. Промежуточная (плащевая, или мантийная) зона состоит из клеток, переместившихся из вентрикулярной и субвентрикулярной зон — нейробластов и глиобластов. Нейробласты утрачивают способность к делению и в дальнейшем дифференцируются в нейроны. Глиобласты продолжают делиться и дают начало астроцитам и олигодендроцитам. Способность к делению не утрачивают полностью и зрелые глиоциты. Новообразование нейронов прекращается в раннем постнатальном периоде. Поскольку число нейронов в головном мозге составляет примерно 1 триллион, очевидно, в среднем в течение всего пренатального периода в 1 мин формируется 2,5 миллиона нейронов. Из клеток плащевого слоя образуются серое вещество спинного и часть серого вещества головного мозга. Маргинальная зона (или краевая вуаль) формируется из врастающих в нее аксонов нейробластов и макроглии и дает начало белому веществу. В некоторых областях головного мозга клетки плащевого слоя мигрируют дальше, образуя кортикальные пластинки — скопления клеток, из которых формируется кора большого мозга и мозжечка (т.е. серое вещество). По мере дифференцировки нейробласта, изменяется субмикроскопическое строение его ядра и цитоплазмы. Специфическим признаком начавшейся специализации нервных клеток следует считать появление в их цитоплазме тонких фибрилл — пучков нейрофиламентов и микротрубочек. Количество нейрофиламентов, содержащих белок — нейрофиламентный триплет, в процессе специализации увеличивается. Тело нейробласта постепенно приобретает грушевидную форму, а от его заостренного конца начинает развиваться отросток — аксон. Позднее дифференцируются другие отростки — дендриты. Нейробласты превращаются в зрелые нервные клетки — нейроны. Между нейронами устанавливаются контакты (синапсы). В процессе дифференцировки нейронов из нейробластов различают до-медиаторный и медиаторный периоды. Для домедиаторного периода характерно постепенное развитие в теле нейробласта органелл синтеза — свободных рибосом, а затем эндоплазматической сети. В медиаторном периоде у юных нейронов появляются первые пузырьки, содержащие нейромедиатор, а в дифференцирующихся и зрелых нейронах отмечаются: значительное развитие органелл синтеза и секреции, накопление медиаторов и поступление их в аксон, образование синапсов. Несмотря на то, что формирование нервной системы завершается только в первые годы после рождения, известная пластичность центральной нервной системы сохраняется до старости. Эта пластичность может выражаться в появлении новых терминалей и новых синаптических связей. Нейроны центральной нервной системы млекопитающих способны формировать новые ветви и новые синапсы. Пластичность проявляется в наибольшей степени в первые годы после рождения, но частично сохраняется и у взрослых — при изменении уровней гормонов, обучении новым навыкам, травме и других воздействиях. Хотя нейроны постоянны, их синаптические связи могут модифицироваться в течение всей жизни, что может выражаться, в частности, в увеличении или уменьшении их числа. Пластичность при малых повреждениях мозга проявляется в частичном восстановлении функций. В популяции нейронов, начиная с ранних стадий развития нервной системы и в течение всего онтогенеза, имеет место массовая гибель клеток. Эта запрограммированная физиологическая гибель клеток наблюдается как в центральной, так и в периферической нервной системе. У человека ежегодно погибает около 10 млн нервных клеток. Нейроны, или нейроциты, — специализированные клетки нервной системы, ответственные за получение, обработку и передачу сигнала (на: другие нейроны, мышечные или секреторные клетки). Нейрон является морфологически и функционально самостоятельной единицей, но с помощью своих отростков осуществляет синаптический контакт с другими нейронами, образуярефлекторные дуги — звенья цепи, из которой построена нервная система. В зависимости от функции в рефлекторной дуге различают три типа нейронов:
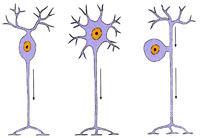
Афферентные (или рецепторные, чувствительные) нейроны воспринимают импульс, эфферентные (или двигательные) передают его на ткани рабочих органов, побуждая их к действию, а ассоциативные (или вставочные) осуществляют связь между нейронами. Подавляющее большинство нейронов (99,9%) - ассоциативные. Нейроны отличаются большим разнообразием форм и размеров. Например, диаметр тел клеток-зерен коры мозжечка 4—6 мкм, а гигантских пирамидных нейронов двигательной зоныкоры большого мозга — 130—150 мкм. Нейроны состоят из тела (или перикариона) и отростков: одного аксона и различного числа ветвящихся дендритов. По количеству отростков различают три типа нейронов:
Униполярные нейроны имеют только аксон (у высших животных и человека обычно не встречаются). Биполярные - имеют аксон и один дендрит. Мультиполярные нейроны (подавляющее большинство нейронов) имеют один аксон и много дендритов. Разновидностью биполярных нейронов является псевдо-униполярный нейрон, от тела которого отходит один общий вырост — отросток, разделяющийся затем на дендрит и аксон. Псевдоуниполярные нейроны присутствуют в спинальных ганглиях, биполярные — в органах чувств. Большинство нейронов - мультиполярные. Их формы чрезвычайно разнообразны. Аксон и его коллатерали оканчиваются, разветвляясь на несколько веточек, называемых телодендронами, последние заканчиваются терминальными утолщениями. Трехмерная область, в которой ветвятся дендриты одного нейрона, называетсядендритным полем нейрона. Дендриты представляют собой истинные выпячивания тела клетки. Они содержат те же органеллы, что и тело клетки: глыбки хроматофильной субстанции (т.е. гранулярной эндоплазматической сети и полисом), митохондрии, большое количество нейротубул (или микротрубочек) и нейрофиламентов. За счет дендритов рецепторная поверхность нейрона увеличивается в 1000 и более раз. Аксон — это отросток, по которому импульс передается от тела клетки. Он содержит митохондрии, нейротубулы и нейрофиламенты, а также гладкую эндоплазматическую сеть. Подавляющее большинство нейронов человека содержит одно округлое светлое ядро, расположенное в центре клетки. Двуядерные и тем более многоядерные нейроны встречаются крайне редко. Плазмолемма нейрона является возбудимой мембраной, т.е. обладает способностью генерировать и проводить импульс. Ее интегральными белками являются белки, функционирующие как ионно-избирательные каналы, и рецепторные белки, вызывающие реакции нейронов на специфические стимулы. В нейроне мембранный потенциал покоя равен —60 —70 мВ. Потенциал покоя создается за счет выведения Na+ из клетки. Большинство Na+- и К+-каналов при этом закрыты. Переход каналов из закрытого состояния в открытое регулируется мембранным потенциалом. В результате поступления возбуждающего импульса на плазмолемме клетки происходит частичная деполяризация. Когда она достигает критического (порогового) уровня, натриевые каналы открываются, позволяя ионам Na+ войти в клетку. Деполяризация усиливается, и при этом открывается еще больше натриевых каналов. Калиевые каналы также открываются, но медленнее и на более продолжительный срок, что позволяет К+ выйти из клетки и восстановить потенциал до прежнего уровня. Через 1—2 мс (т.н. рефрактерный период) каналы возвращаются в нормальное состояние, и мембрана может вновь отвечать на стимулы. Итак, распространение потенциала действия обусловлено вхождением в нейрон ионов Na+, которые могут деполяризовать соседний участок плазмолеммы, что в свою очередь создает потенциал действия на новом месте. При окрашивании нервной ткани анилиновыми красителями в цитоплазме нейронов выявляется хроматофильная субстанция в виде базофильных глыбок и зерен различных размеров и форм (другие названия хроматофильной субстанции - тигроид, тельца Ниссля). Базофильные глыбки локализуются в перикарионах и дендритах нейронов, но никогда не обнаруживаются в аксонах и их конусовидных основаниях — аксональных холмиках. Базофилия глыбок объясняется высоким содержанием рибонуклеопротеидов. Каждая глыбка хроматофильной субстанции состоит из цистерн гранулярной эндоплазматической сети, свободных рибосом и полисом. Для поддержания целостности нейронов и выполнения ими функций нейронам требуется огромное количество белков. Для аксонов, не имеющих органелл белкового синтеза, характерен постоянный ток цитоплазмы от перикариона к терминалям со скоростью 1—3 мм в сутки. Возрастные изменения нейронов сопровождаются накоплением липофусцина, разрушением крист митохондрий. Липофусцин — «пигмент старения» — желто-бурого цвета липопротеидной природы, представляющий собой остаточные тельца (т.е. телолизосомы) с продуктами непереваренных структур. 43. Нервные волокна: определение, строение и функции миели и безмиелин волокон. Регенерация. |