Влияние натриево-сульфатного засоления почв на рост пшеницы. ВЛИЯНИЕ НАТРИЕВО-СУЛЬФАТНОГО ЗАСОЛЕНИЯ. 1 Растения в условиях стресса и механизмы адаптации
 Скачать 490.5 Kb. Скачать 490.5 Kb.
|

|
Содержание
Введение Засоленные почвы занимают около 30% всей суши. По некоторым данным [Удовенко, 1978; Блэк, 1973] в бывшем СССР засолено около 10%, а в отдельных районах - до 90% всей посевной площади. В последнее время кроме почв первичного засоления, развивающихся естественным путем в силу природных явлений, увеличивается и доля почв вторичного засоления, развивающихся не без помощи человека. Масштабы вторичного засоления значительны — несколько десятков тысяч гектаров в год. Есть серьезные опасения, что со временем вторичное засоление может стать важным фактором, ограничивающим сельскохозяйственное производство. Борьба с засолением почв приобрела народнохозяйственное значение, поэтому в последнее время вопросам солеустойчивости растений уделяется огромное внимание [Третьяков, 2000]. Наряду с правильной агротехникой и мелиоративными путями ослабления уровня отрицательного воздействия засоления почвы, важное значение имеет задача повышения устойчивости самих растений к стрессовым условиям [Жученко, 1988]. Целью работы являлось исследование влияние натриево-сульфатного засоления почв на рост пшеницы В задачи работы входило: 1. исследование влияния натриево-сульфатного засоления на рост корней и калеоптилей пщеницы. 2. исследование влияния натриево-сульфатного засоления на накопление биомассы пщеницы. Глава 1. Литературный обзор 1.1. Засоление почв За последние 50 лет уровень Мирового океана поднялся на 10 см. Эта тенденция, по предсказаниям ученых, будет продолжаться и дальше. Следствием этого является возрастающий дефицит пресной воды, а доля засоленных вод увеличивается. Таким образом, к обширным территориям в той или иной степени засоления будут прибавляться новые. Однако кроме почв первичного засоления, развивающихся естественным путем в силу природных явлений, увеличивается и доля почв вторичного засоления, развивающихся не без помощи человека, чаще всего при нерациональном орошении. Масштабы вторичного засоления значительны — несколько десятков тысяч гектаров в год. Есть серьезные опасения, что со временем вторичное засоление может стать важным фактором, ограничивающим сельскохозяйственное производство. Борьба с засолением почв приобрела народнохозяйственное значение, поэтому в последнее время вопросам солеустойчивости растений уделяется огромное внимание [Третьяков, 2000]. Засоление связано главным образом с повышенным содержанием натрия в почве. В зависимости от преимущественного накопления отдельных солей натрия засоление может быть сульфатным, хлоридным, содовым или смешанным. Наиболее вредное влияние оказывают ионы Na+ и С1-. Засоление приводит к созданию в почве низкого (резко отрицательного) водного потенциала, поэтому поступление воды в растение сильно затруднено [Якушкина, 1993]. Засоление почв может происходить самыми различными способами. Одним из них является неумеренный, бессистемный полив при отсутствии дренажа. Засоление почв имеет значительные масштабы и представляет опасность для поливного земледелия. Вторичное засоление ранее не засоленных почв может происходить двояким путем. Во-первых, при неглубоком залегании минерализованные грунтовые воды, поднимаясь по капиллярам почвы и испаряясь, оставляют соли у поверхности. Во-вторых, при неумеренном поливе происходит подъем грунтовых вод, заболачивание и засоление почвы солями, растворенными в этих водах. Оросительные системы иногда при этом полностью выходят из строя. Засоление может наблюдаться и при глубоком залегании грунтовых вод, если в почвогрунте лежат уплотненные слои, служащие водоупором. Образующаяся под этими слоями верховодка способствует капиллярному подъему и выносу солей в пахотный слой. Явление засоления почв имеет широкое распространение и наблюдается во многих странах орошаемого земледелия Азии, Африки, Америки. В результате вторичного засоления десятки миллионов гектаров из некогда продуктивных земель превратились в малопродуктивные засоленные почвы и соляные пустыни. Ежегодно в разных странах гибнут 200 - 300 тыс. га поливных земель от засоления и заболачивания. Для предотвращения вторичного засоления земель большое значение имеет дренаж, который необходим при близком залегании минерализованных грунтовых вод. Применяют промывание засоленных почв. Для полной ликвидации засоления рекомендуется строительство каналов в лотках и бетонных ложах [Охрана природы, 1987]. Растения в условиях стресса и механизмы адаптации. При любых экстремальных воздействиях в растительном организме происходят изменения различных физиологических параметров, что обусловлено взаимосвязанностью отдельных процессов в растении и саморегулируемостью его метаболизма в целом [Елисеева, 1987; Жученко, 1987]. Клетки растений реагируют сходными изменениями на различные стрессы [Удовенко, 1978]. В основе такой неспецифической реакции клетки лежит изменение конформационной подвижности белковых молекул. Неспецифическая реакция растительного организма проявляется и на других, более высоких уровнях организации биологических систем [Мирославов, 1997]. Выделяют первичные изменения, обусловленные непосредственным действием стресса на растение и вторичные, вызванные первичными нарушениями метаболических функций [Зауралов, 1989]. К первичным относятся: изменения осморегуляции клетки, биоэнергетических процессов, структурной целостности и состава мембран, а также изменение структурного состояния и обусловленная им функциональная активность ядерной ДНК [Строганов, 1962]. К вторичным относится большая часть физиологических изменений в клетке и целом организме. Их общей чертой является причинная зависимость от физиологических процессов, первично изменяющихся в результате стресса. Функционально наиболее важное значение среди вторичных отклонений имеет торможение белкового синтеза, увеличение концентрации фитогормонов, подавляющих деление и растяжение клеток [Удовенко, 1979]. Существуют определенные закономерности в изменении физиологических параметров под воздействием стресса, которые можно охарактеризовать двумя типами кривых (рис. 1) [Жученко, 1987]. Обнаружено, что при стабильном и длительном экстремальном воздействии уровень многих параметров меняется экспоненциально и в дальнейшем сохраняется на отличном от оптимальной нормы уровне (рис.1а). Другая же группа параметров изменяется по одновершинной кривой, сначала существенно отклоняясь от нормы, а затем постепенно возвращаясь к исходному уровню или близко к нему (рис.1б). Р 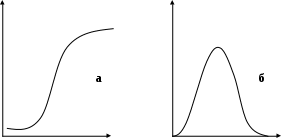 ис. 1. Изменение физиологических параметров под воздействием cтресса: а – по логарифмическому, б – по одновершинному типу По первому типу изменяются параметры, характеризующие первичные нарушения и интегральные изменения. По второму типу в экстремальных условиях меняются параметры вторичных отклонений. Они характеризуют либо соотношение скоростных процессов, противоположно направленных (например, синтез-гидролиз), или относительно независимых между собой (накопление биомассы, поглощение минеральных ионов), либо состояние постоянно обновляемых активно функционирующих биологических структур (внутриклеточных мембран и органелл) [Жученко, 1987]. Предполагается, что именно изменения второго типа реально отражают сущность саморегулируемости метаболизма живой клетки и тем самым непосредственно характеризуют адаптацию растительного организма к стрессовым условиям [Альтергот, 1982]. Важнейшими механизмами адаптации растений к неблагоприятным факторам являются снижение скорости катаболических процессов и более полная мобилизация потенциальных возможностей различных реакций, компенсаторные изменения сопряженных процессов, а также включение шунтовых механизмов [Удовенко, 1979]. Установлено, что в нормальных условиях интенсивность различных процессов значительно ниже максимально возможного и при неблагоприятных условиях может возрастать до такого уровня. Например, у растений на засоленном фоне в течение некоторого времени скорость фотосинтеза остается на том же уровне, что и на пресном [Белецкий, 1990]. Обнаружены некоторые компенсаторные изменения сопряженных процессов. В частности, для компенсации дефицита энергетического баланса организма у растений в условиях солевого стресса обычно усиливается интенсивность дыхания при снижении его энергетической эффективности. У слабоустойчивых растений, которые существенно угнетаются солями, такого стимулирования дыхания не наблюдается или оно незначительно, и обнаружен резкий дефицит их энергетического баланса. Увеличение числа изоферментов в условиях стресса, повышение активности одних ферментов при подавлении других, катализирующих превращение тех же субстратов, активизация пентозофосфатного пути дыхания при ослаблении гликолитического свидетельствует о включении различных шунтовых механизмов при неблагоприятных для растения условиях окружающей среды [Жученко, 1988]. Отмечено, что наиболее эффективная организменная адаптация растений к длительно действующим неблагоприятным факторам осуществляется через новообразование более устойчивых к стрессу клеток и органов в процессе роста. То есть ростовые реакции, очевидно, являются интегрирующими при определении адаптации растений к стрессовым условиям [Кузнецов и др., 1991; Гамбарова, 1997]. Таким образом, адаптации растений к различным неблагоприятным факторам окружающей среды являются неспецифическими и определяются сложным комплексом процессов, координируемых системой саморегуляции растений. О почвах говорят, что они засолены, если они содержат избыток растворимых солей в корнеобитаемом слое [Блэк, 1973]. Главными катионными компонентами растворимых солей в засоленных почвах являются натрий, кальций, магний, а главными анионными компонентами - сульфаты, хлориды и карбонаты. Второстепенными компонентами могут быть калий, бикарбонаты, нитраты [Строганов, 1962]. Чаще всего засоление представлено ионами натрия в сочетании с ионами хлора, сульфата или карбоната. Почвы с карбонатно-натриевым засолением практически непригодны для произрастания большинства растений. Поэтому наиболее подробно изучается влияние на растения хлорно - и сульфатно-натриевого засоления [Удовенко, 1978]. Уровень накопления иона соли в растительном организме при данной концентрации его в среде обусловливается совокупностью двух определяющих факторов: сорбционной емкости и растворяющей способности (или коэффициента растворимости по отношению к воде) цитоплазмы [Блэк, 1973; Строганов, 1962]. Ионы хлора аккумулируются в основном в надземных органах, прежде всего в молодых листьях. Натрий также в большей степени аккумулируется в надземных органах, особенно в стеблях. Концентрации сульфат - иона оказываются наиболее высокими в корнях [Удовенко, 1977]. Неодинаково распределение ионов при засолении у сортов с разной степенью солеустойчивости: более устойчивые формы локализуют соли преимущественно в корнях, представляющих для них своеобразный барьер, а у неустойчивых основная масса ионов накапливается в надземных органах. При засолении претерпевают изменения разнообразные физиологические параметры, что обусловлено взаимосвязанностью физиологических функций в самокоординируемой системе растительного организма. Эти изменения имеют динамический характер, отражающий последовательно идущие в клетках фазы раздражения, повреждения и адаптации организма к засолению [Альтергот, 1982]. Четкое представление о характере этих изменений необходимо для разработки эффективных способов повышения солеустойчивости растений. Физиологические процессы солеустойчивости растений лежат в пределах осмотического давления 0,3-1,8 мПа, что соответствует концентрации хлорида натрия 0,42-2,52% [Елисеева, 1987]. Засоление оказывает серьезное ингибиторное влияние прежде всего на рост растений. В связи с этим выделяют ионные, осмотические и токсичные эффекты воздействия солей на растение [Строганов, 1962; Жученко, 1988]. При засолении у растений повышается осмотический потенциал клеточного сока, вследствие накопления в цитоплазме гидрофильных, осмотически активных ионов солей [Белянская, Исханов, Шамина, 1991]. Наряду с изменением уровня осмотического потенциала при засолении повышается и проницаемость протоплазмы клеток. Установлено, что при этом выделяются кальций и белки. Одновалентные катионы солей разрушают связь кальция со структурными и растворимыми белками клетки, обуславливая этим изменение свойств цитоплазмы. Добавление кальция в среду восстанавливает проницаемость протоплазмы клеток [Калинин, 1980]. Также постоянно наблюдаемым эффектом воздействия солей на клетки является отхождение протоплазмы от клеточных стенок. При этом происходит разобщение плазмодесм и нарушение межклеточных связей [Удовенко, 1977]. Помимо осмотического действия, обуславливающего и обезвоживание, прямое влияние солей заключается в сдвиге ионного равновесия, которое изменяет все внутриклеточные процессы, а также неблагоприятно действует на протоплазму, органоиды и биополимеры клетки [Филатова, Мельникова, 1987]. В условиях засоления у большинства культивируемых видов растений изменяется интенсивность дыхания, снижается интенсивность фотосинтеза [Белецкий, 1990], синтеза белков и нуклеиновых кислот [Филатова, Мельникова, 1987], фосфорного обмена [Строганов, 1962], поглощения элементов минерального питания и их метаболического использования в надземных органах [Хориков, 1990; Климашевский, 1991], а также доступность воды для растения (состояние «физиологической засухи»), что в целом приводит к заметному снижению роста [Удовенко, 1977]. Одной из причин торможения ростовых процессов у растений при засолении является также заметное изменение содержания и соотношения эндогенных стимуляторов и ингибиторов роста [Подлиток, 1987; Рейтарова, Черников, 1991]. К одной из главных причин повреждения растений в условиях засоления Б.П.Строганов [1962], относит нарушение процессов азотного обмена. При этом в частности, замедляется синтез белков, усиливается гидролиз запасных белков, что приводит к появлению токсичных диаминов (таких, как путресцин и кадаверин), вызывающих появление некрозов у растений [Филатова, Мельникова, 1987; Жученко, 1988]. Согласно Жученко [1980], при солевом стрессе возможны повреждения мембран клеток как следствие изменения липидного комплекса, денатурации и агрегации белков, нарушения обменных процессов, возникновения осмотического стресса и стресса, обусловленного «голоданием» растений. Считается, что эти нарушения носят вторичный, косвенный характер. Эффект торможения процессов роста и развития растений при засолении проявляется, начиная с прорастания, наклевывания семян, роста проростка и сохраняется в течение всего периода вегетации [Удовенко, 1978]. Торможение синтетических процессов, в первую очередь белка и нуклеиновых кислот, и понижение энергообеспеченности организма ведет не только к резкому сокращению ростовых процессов, но и создает у растений диспропорцию синтеза и гидролиза. Если уровень засоления субстрата достигает летального порога или превышает его, то размеры нарушений постоянно возрастают, прогрессивно увеличивается диспропорция между продуктами синтеза и распада в клетках и организм погибает. Однако, когда степень засоления ниже, растения выживают, но при этом интенсивность синтетических реакций в них стабилизируется на новом, пониженном уровне. Таким образом, приспособление к засолению (в том числе и к многим другим экстремальным воздействиям, как считают многие исследователи) в целом выражается в переходе всех основных реакций метаболизма на новый, менее интенсивный уровень [Удовенко, 1975]. За счет этого, с одной стороны, восстанавливается нормальное равновесие между интенсивностью метаболических процессов, что обеспечивает дальнейшее существование организма, как сложной и скоординированной биологической системы. С другой стороны, понижается чувствительность организма к отклоняющимся от оптимальной нормы условиям среды, что объясняет и причину появления у растений так называемой сопряженной солеустойчивости, и сущность явления закаливания. . Солеустойчивость, основные механизмы защиты, методы оценки. По устойчивости к засолению среди видового и экотипического разнообразия наиболее часто выделяют две группы растений: галофиты и гликофиты [Удовенко, 1977; Блэк, 1973]. Среди культурных растений практически все виды являются гликофитами [Калинин, 1980]. При оценке солеустойчивости исследователи различают два аспекта - биологический и агрономический [Удовенко, 1977; Строганов, 1962]. С биологической точки зрения под солеустойчивостью вида или сорта понимают тот предел засоления, при котором растения еще способны полностью завершить онтогенетический цикл развития и воспроизвести всхожие семена. Выше этого предела растения гибнут раньше завершения цикла онтогенеза или образуют невсхожие семена с недоразвитым зародышем [Жученко, 1987]. При агрономическом подходе к солеустойчивости растений обращают внимание насколько данный сорт или вид снижает свою продуктивность в конкретных условиях засоления (при определенном его уровне) по сравнению с его урожайностью без засоления [Удовенко, Гончарова, 1982]. Известно, что ионы соли находятся в протоплазме в двух различных состояниях: адсорбционно - связанном белками и другими полимерными молекулами и свободно-растворенном в жидкой фазе цитоплазмы. Очевидно, что при одном и том же уровне содержания иона в клетке эффект воздействия его на метаболизм будет различным по величине при разном соотношении количеств иона в этих состояниях. Именно этим, считает Г. В. Удовенко [1977], определяется уровень устойчивости растений к избытку засоляющих ионов. Большинство авторов в настоящее время выделяют три вероятных способа защиты от солевого стресса: 1. «Осмотическая защита» - поддержание тургора клетки активацией биосинтеза осмопротекторов, балансирование внутриклеточного осмотического давления аккумуляцией осмотически активных веществ, активация ферментов с защитными свойствами [Белянская, Исханов, Шамина, 1991; Жученко, 1987; Блэк, 1973]; 2. «Вакуолярная защита» - предотвращение токсичности действия ионов путем компартментализации в метаболически инертных структурах [Удовенко, 1977; Дридзе, Майсурян, 1993] 3. «Мембранная защита» - изменения в структуре клеточной мембраны, препятствующие проникновению токсичных ионов в клетку или обеспечивающие их выброс за пределы клетки с помощью «протонной помпы» [Строганов, 1962; Климашевский, 1991]. Только сочетание всех трех вариантов защиты от стрессовой ситуации позволяет клетке проявить максимальную резистентность к воздействию соли [Дридзе, Майсурян, 1993]. Свойство солеустойчивости на организменном уровне может обеспечиваться за счет накопления концентрации солей в тканях и частях растения, не несущих большой метаболической нагрузки и выполняющих осмотические функции без значительного воздействия на ферментативную активность. В условиях высокого содержания хлорида натрия в субстрате некоторые виды растений освобождаются от избытка ионов с помощью железистых клеток [Жученко, 1987]. Растения других видов экскретируют излишнее количество солей через специальные выделительные клетки, расположенные на верхней и нижней поверхностях листьях, поддерживая, таким образом концентрацию ионов в тканях на оптимальном уровне [Филатова, Мельникова, 1987]. Устойчивость к засолению разных сельскохозяйственных культур и сортов меняется в процессе онтогенеза. При этом на ранних его этапах, а также в период формирования репродуктивных органов, растение обычно характеризуется наименьшей солеустойчивостью [Бессонова, 1995]. Например, ячмень, сахарная свекла и хлопчатник относятся к числу наиболее толерантных к засолению культур, но их прорастающие семена и проростки весьма чувствительны к засолению [Хориков, 1990]. Таким образом, солеустойчивость является результатом совокупного действия ряда защитных механизмов, детерминированных в клетке, скоростью адаптивных реакций, а также многовариантностью путей реализации онтогенетического развития растительного организма. Существует большое количество прямых и косвенных физиологических методов оценки солеустойчивости растений, основанных на учете различных параметров. Большая группа методов включает оценку прорастания семян в солевых растворах [Удовенко, Гончарова, 1982]. Тестирующим признаком служит депрессия накопления биомассы и депрессия роста проростков в солевом растворе по сравнению с проростками пресного контроля [Иванов, Удовенко, 1970; Семушина, Лукьянова, 1979]. Многие исследователи подчеркивают, что детальная информация о степени солеустойчивости различных растений в первый период роста может способствовать рентабельному возделыванию их на засоленных землях [Быстров, Коваль, 1996; Иванов, Удовенко, 1970]. Метод проростков позволяет изучать способы повышения биологической солеустойчивости сортов; сопоставлять солеустойчивость семян различных видов и сортов, выделять наиболее устойчивые из них для возделывания на полях с учетом уровня засоления; определять уровень солестойкости данного сорта; выявлять устойчивые формы растений для целей селекции; устанавливать предел засоления, при котором возможно прорастание семян, точно разграничивать сорта по интенсивности роста проростков при засолении [Иванов, Удовенко, 1970]. При этом долгое время стоял вопрос о возможности диагностики солеустойчивости сортообразцов путем их оценки в первую фазу роста. В результате многочисленных исследований солеустойчивости культурных растений физиологическим и вегетационным методом, Г. В. Удовенко [1975] обнаружена пригодность метода проростков для сравнительной характеристики солеустойчивости культурных растений. Вегетационный и вегетационно-полевой методы оценки солеустойчивости растений являются интегральными и наиболее точными, но в то же время наиболее длительными и трудоемкими. В этом случае растения выращивают по обычной методике вегетационного или полевого опыта на двух фонах - незасоленном [контроль] и засоленном. Солеустойчивость растений определяют по степени снижения урожайности сорта или культуры на засоленном фоне по сравнению с урожайностью его на контроле [Удовенко, Гончарова, 1982]. |