Биология экзамен. 1 Раздел I. Общая характеристика жизни
 Скачать 0.75 Mb. Скачать 0.75 Mb.
|

|
Первичная структура – полинуклеотидная цепь, мономеры (нуклеотиды) соединяются фосфоди-эфирными связями (сборка цепи за счет фермента полимеразы). Наращивание цепи идет в направлении 5/ --------3/ Вторичная структура ДНК – две полинуклеотидные цепи (антипараллельны), связанные водородными связями Третьичная структура – трехмерная структура ДНП Функции ДНК Молекула ДНК человека — носитель генетической информации, которая записана в виде последовательности нуклеотидов с помощью генетического кода. В результате описанной выше репликации ДНК происходит передача генов ДНК от поколения к поколению. Свойства ДНК: Способность кодировать информацию (генетический код). Способность воспроизводить информацию (репликация). Способность реализовать информацию (трансляция). Способность правильно сохранять информацию (репарация). Способность передавать информацию (транскрипция). Способность изменять информацию (мутация и генетическая рекомбинация). Самовоспроизведение — способность живого организма, его органа, ткани, клетки или клеточного органоида или включения к образованию себе подобного.Репликация (от лат. replicatio — возобновление) — процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты на матрице родительской молекулы ДНК. В ходе последующего деления материнской клетки каждая дочерняя клетка получает по одной копии молекулы ДНК, которая является идентичной ДНК исходной материнской клетки. Этот процесс обеспечивает точную передачу генетической информации из поколения в поколение. Репликацию ДНК осуществляет сложный ферментный комплекс, состоящий из 15—20 различных белков, называемый реплисомой. Репликация ДНК — ключевое событие в ходе деления клетки. Принципиально, чтобы к моменту деления ДНК была реплицирована полностью и при этом только один раз. Это обеспечивается определёнными механизмами регуляции репликации ДНК. Репликация проходит в три этапа: 1. инициация репликации 2. элонгация 3. терминация репликации. Регуляция репликации осуществляется в основном на этапе инициации. Это достаточно легко осуществимо, потому что репликация может начинаться не с любого участка ДНК, а со строго определённого, называемого сайтом инициации репликации. Репликация начинается в сайте инициации репликации с расплетания двойной спирали ДНК, при этом формируется репликационная вилка — место непосредственной репликации ДНК. В репликационной вилке ДНК копирует крупный белковый комплекс (реплисома), ключевым ферментом которого является ДНК-полимераза. Репликационная вилка движется со скоростью порядка 100 000 пар нуклеотидов в минуту у прокариот и 500—5000 — у эукариот. матричный— последовательность синтезируемой цепи ДНК однозначно определяется последовательностью материнской цепи в соответствии с принципом комплементарности; · полуконсервативный— одна цепь молекулы ДНК, образовавшейся в результате репликации, является вновь синтезированной, а вторая — материнской; идёт в направлении от 5’-конца новой молекулы к 3’-концу; · полунепрерывный— одна из цепей ДНК синтезируется непрерывно, а вторая — в виде набора отдельных коротких фрагментов (фрагментов Оказаки); начинается с определённых участков ДНК, которые называются сайтами инициации репликации. Этапы РЕПЛИКАЦИИ: 1 - Разделение материнской цепи на 2 матричные нити (работает фермент ГЕЛИКАЗА) 2 - Дестабилизирующие белки располагаются вдоль каждой полинуклеотидной цепи (роль: растяжение нити и доступность для + комплементарных нуклеотидов) РЕПАРАЦИЯ – коррекция нарушений соединения под влиянием реакционно- способных веществ или УФ. Разновидности: - ЭКСЦИЗИОННАЯ или дорепликативная (с вырезками) - ПОСТРЕПЛИКАТИВНАЯ (путем рекомбинации) - СВЕТОВАЯ РЕПАРАЦИЯ (самопроизвольное устранение нарушений под действием видимого света). РЕПАРАЦИЯ - При наличии большого объема поражений включается система индуцируемых ферментов репарации (SOS система: восстановление может идти без соблюдения принципа комплементарности, что ведет к стойким изменениям – мутациям) - При значительном повреждении – блокада репликации ДНК. 4. Ген, его свойства. Ген как функциональная единица наследственности. Классификация генов. Особенности организации генов у про- и эукариот. Генетический код как способ записи наследственной информации, его свойства. Цистрон, его структура. Элементарной функциональной единицей наследственности является ГЕН Первые представления о сложной структуре гена возникли в 20- х годах прошлого столетия. Советские генетики А.С. Серебровский и Н.П.Дубинин выдвинули предположение о дискретной структуре гена Американский ученый Бензер предложил назвать часть гена цистроном Американский ученый Гильберт в 1978 г. Установил , что ген эукариот состоит из информативных и неинформативных участков Свойства гена: дискретность — несмешиваемость генов; стабильность — способность сохранять структуру; лабильность — способность многократно мутировать; множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм; аллельность — в генотипе диплоидных организмов только две формы гена; специфичность — каждый ген кодирует свой признак; плейотропия — множественный эффект гена; экспрессивность — степень выраженности гена в признаке; пенетрантность — частота проявления гена в фенотипе; амплификация — увеличение количества копий гена[ Классификация генов: Структурные гены : Независимые гены – их транскрипция не связана с функциональными генами, а напрямую регулируется гормонами Повторяющиеся гены (тандемные) – так устроены гены, несущие информацию о РНК Кластерные гены – группы различных генов, объединенных одной функцией Функциональные гены: Оператор – относится к группе акцепторов. Определяет время, с которого начинается транскрипция Промотор –участок ДНК, включает 80-90 нп. Способен связываться с ДНК – зависимой РНК – полимеразой. Полимераза узнает участок ТАТААТ, который называется блок Прибнова. В этом месте ДНК плотно не упаковывается. Промотор определяет место, с которого начинается транскрипция Энхансер – увеличивает скорость транскрипции Сайленсер – снижает скорость транскрипции Спейсер – неинформативный участок геном Псевдогены – НП, в которых полимераза не работает, в связи с мутацией Терминатор - ген, на котором заканчивается транскрипция. Находится на 3’ конце. Включает палиндром
Код наследственности – способ зашифровки в молекуле ДНК наследственной информации о структуре и функции белков Свойства кода (М.Ниренберг, 1963 г.): Колинеарность - параллелизм. Нуклеотидная последовательность ДНК соответствует аминокислотной последовательности белка Триплетность –каждая аминокислота кодируется тройкой нуклеотидов – триплетом. Из четырех нуклеотидов путем различных сочетаний можно получить 64 триплета - кодона. Неперекрываемость – перекрываемость – при неперекрываемости один и тот же нуклеотид не может одновременно принадлежать двум кодонам: Перекрываемость – заключается в том, что с одного и того же участка ДНК может считываться информация для образования двух и более белков в зависимости от начальной точки считывания Вырожденность – экспериментально установлено, что при триплетности все 64 кодона имеют значение в экспрессии генов. Из них 61 кодон кодирует аминокислоты, а 3 кодона являются стоп – кодонами: УГА,УАГ,УАА. Универсальность – кодирование аминокислот происходит одинаково на всех уровнях организации живой системы Квазиуниверсальность – некоторые кодоны в разных генетических системах кодируют различные аминокислоты Цистрон: Ген как функциональную единицу предложено называть цистроном. Именно цистрон определяет последовательность аминокислот в каждом специфическом белке. Цистрон, в свою очередь, подразделяется на предельно малые в линейном измерении единицы - реконы, способные к рекомбинации при кроссинговере. Выделяют, кроме того, мутоны - наименьшие части гена, способные к изменению (мутированию). Размеры рекона и мутона могут равняться одной или нескольким парам нуклеотидов, цистрона - сотням и тысячам нуклеотидов. 5. Этапы реализации генетической информации. Транскрипция и посттранскрипционные процессы. Регуляция 6. Трансляция и посттрансляционные процессы. Структура и виды РНК, роль РНК в процессе реализации наследственной информации. Регуляция РНК — полимер, мономерами которой являются рибонуклеотиды. В отличие от ДНК, РНК образована не двумя, а одной полинуклеотидной цепочкой (исключение — некоторые РНК-содержащие вирусы имеют двухцепочечную РНК). Нуклеотиды РНК способны образовывать водородные связи между собой. Цепи РНК значительно короче цепей ДНК. Мономер РНК — нуклеотид (рибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания РНК также относятся к классам пиримидинов и пуринов. Пиримидиновые основания РНК — урацил, цитозин, пуриновые основания — аденин и гуанин. Моносахарид нуклеотида РНК представлен рибозой. Выделяют три вида РНК: 1) информационная (матричная) РНК — иРНК (мРНК), 2) транспортная РНК — тРНК, 3) рибосомная РНК — рРНК. Все виды РНК представляют собой неразветвленные полинуклеотиды, имеют специфическую пространственную конформацию и принимают участие в процессах синтеза белка. Информация о строении всех видов РНК хранится в ДНК. Процесс синтеза РНК на матрице ДНК называется транскрипцией. Транспортные РНК содержат обычно 76 (от 75 до 95) нуклеотидов; молекулярная масса — 25 000–30 000. На долю тРНК приходится около 10% от общего содержания РНК в клетке.Функции тРНК: 1) транспорт аминокислот к месту синтеза белка, к рибосомам, 2) трансляционный посредник. В клетке встречается около 40 видов тРНК, каждый из них имеет характерную только для него последовательность нуклеотидов. Однако у всех тРНК имеется несколько внутримолекулярных комплементарных участков, из-за которых тРНК приобретают конформацию, напоминающую по форме лист клевера. У любой тРНК есть петля для контакта с рибосомой (1), антикодоновая петля (2), петля для контакта с ферментом (3), акцепторный стебель (4), антикодон (5). Аминокислота присоединяется к 3'-концу акцепторного стебля.Антикодон — три нуклеотида, «опознающие» кодон иРНК. Следует подчеркнуть, что конкретная тРНК может транспортировать строго определенную аминокислоту, соответствующую ее антикодону. Специфичность соединения аминокислоты и тРНК достигается благодаря свойствам фермента аминоацил-тРНК-синтетаза. Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000. На долю рРНК приходится 80–85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках. Функции рРНК: 1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом; 2) обеспечение взаимодействия рибосомы и тРНК; 3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания, 4) формирование активного центра рибосомы. Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000 000). На долю иРНК приходится до 5% от общего содержания РНК в клетке. Функции иРНК: 1) перенос генетической информации от ДНК к рибосомам, 2) матрица для синтеза молекулы белка, 3) определение аминокислотной последовательности первичной структуры белковой молекулы. Фаза инициации, или начало синтеза пептида, заключается в объединении двух находящихся до этого порознь в цитоплазме субчастиц рибосомы на определенном участке мРНК и присоединении к ней первой аминоацил-тРНК. Этим задается также рамка считывания информации, заключенной в мРНК (рис. 3.32). В молекуле любой мРНК вблизи ее 5'-конца имеется участок, комплементарный рРНК малой субчастицы рибосомы и специфически узнаваемый ею. Рядом с ним располагается инициирующий стартовый кодон АУТ, шифрующий аминокислоту метионин. Малая субчастица рибосомы соединяется с мРНК таким образом, что стартовый кодон АУТ располагается в области, соответствующей П-участку. При этом только инициирующая тРНК, несущая метионин, способна занять место в недостроенном П-участке малой субчастицы и комплементарно соединиться со стартовым кодоном. После описанного события происходит объединение большой и малой субчастиц рибосомы с образованием ее пептидильного и аминоацильного участков (рис. 3.32). 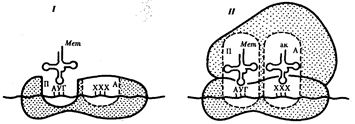 Рис. 3.32. Инициация белкового синтеза: I — соединение малой субчаспщы рибосомы с мРНК, присоединение к стартовому кодону несущей метионин тРНК, которая располагается в недостроенном П-участке; II — соединение большой и малой субчастиц рибосомы с образованием П- и А-участков; следующий этап связан с размещением в А-участке аминоацил-тРНК, соответствующей расположенномув нем кодону мРНК,—начало элонгации; ак — аминокислота К концу фазы инициации П-участок занят аминоацил-тРНК, связанной с метионином, тогда как в А-участке рибосомы располагается следующий за стартовым кодон. Описанные процессы инициации трансляции катализируются особыми белками — факторами инициации, которые подвижно связаны с малой субчастицей рибосомы. По завершении фазы инициации и образования комплекса рибосома — мРНК — инициирующая аминоацил-тРНК эти факторы отделяются от рибосомы. Фаза элонгации, или удлинения пептида, включает в себя все реакции от момента образования первой пептидной связи до присоединения последней аминокислоты. Она представляет собой циклически повторяющиеся события, при которых происходит специфическое узнавание аминоацил-тРНК очередного кодона, находящегося в А-участке, комплементарное взаимодействие между антикодоном и кодоном. Благодаря особенностям трехмерной организации тРНК. (см. разд. 3.4.3.1) при соединении ее антикодона с кодоном мРНК. транспортируемая ею аминокислота располагается в А-участке, поблизости от ранее включенной аминокислоты, находящейся в П-участке. Между двумя аминокислотами образуется пептидная связь, катализуемая особыми белками, входящими в состав рибосомы. В результате предыдущая аминокислота теряет связь со своей тРНК и присоединяется к аминоацил-тРНК, расположенной в А-участке. Находящаяся в этот момент в П-участке тРНК высвобождается и уходит в цитоплазму (рис. 3.33). Перемещение тРНК, нагруженной пептидной цепочкой, из А-участка в П-участок сопровождается продвижением рибосомы по мРНК на шаг, соответствующий одному кодону. Теперь следующий кодон приходит в контакт с А-участком, где он будет специфически «опознан» соответствующей аминоацил-тРНК, которая разместит здесь свою аминокислоту. Такая последовательность событий повторяется до тех пор, пока в А-участок рибосомы не поступит кодон-терминатор, для которого не существует соответствующей тРНК. 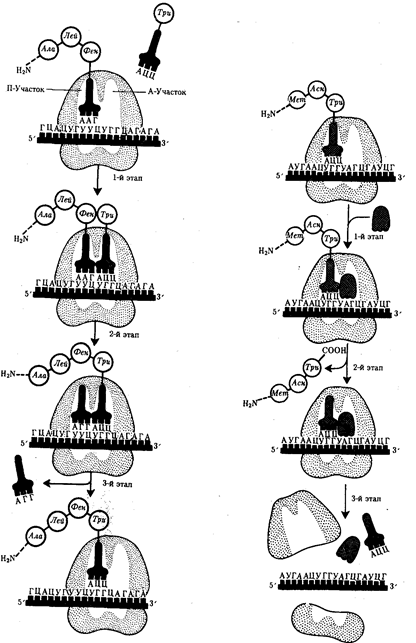
Сборка пептидной цепи осуществляется с достаточно большой скоростью, зависящей от температуры. У бактерий при 37 °С она выражается в добавлении к подипептиду от 12 до 17 аминокислот в 1 с. В эукариотических клетках эта скорость ниже и выражается в добавлении двух аминокислот в 1 с. Фаза терминации, или завершения синтеза полипептида, связана с узнаванием специфическим рибосомным белком одного из терминирующих кодонов (УАА, УАГ или У ГА), когда тот входит в зону А-участка рибосомы. При этом к последней аминокислоте в пептидной цепи присоединяется вода, и ее карбоксильный конец отделяется от тРНК. В результате завершенная пептидная цепь теряет связь с рибосомой, которая распадается на две субчастицы (рис. 3.34). 7. Тонкая структура генов у про- и эукариот. Особенности экспрессии генетической информации у про- и эукариот. Взаимосвязь между геном и признаком. 05 8. Мутации, их классификация, механизмы возникновения. Ген как единица изменчивости. Генные мутации и их классификация. Причины и механизмы возникновения генных мутаций. |