Биология экзамен. 1 Раздел I. Общая характеристика жизни
 Скачать 0.75 Mb. Скачать 0.75 Mb.
|

|
Мутация – внезапное наследственное изменение какого-либо фенотипического признака, вызванное резким структурным или функциональным изменением генетического материала. Генные мутации связаны с изменением внутренней структуры генов, что превращает одни аллели в другие. Можно выделить несколько типов генных мутаций на молекулярном уровне: - замена пар нуклеотидов - делеция (выпадение) - вставка нуклеотида - перестановка (инверсия) участка гена. Замена пар нуклеотидов. Замена пуринового основания на другое пуриновое, или одного пиримидинового на другое пиримидиновое – транзиция.Замена пуринового основания на пиримидиновое и наоборот – трансверсия. При замене нуклеотидов в структурных генах происходит изменение смысла гена – возникают миссенс-мутации. При этом одна аминокислота в полипептиде замещается другой. Фенотипическое проявление мутации зависит от положения аминокислоты в полипептиде. При замене последовательности ЦТЦ на ЦАЦ возникает серповидно-клеточная анемия. Образуется новый полипептид и гемоглобин имеет совсем другие свойства. Некоторые миссенс-мутации приводят к возникновению фермента, обладающего высокой активностью в одних условиях и средней в других условиях. Т.к. генетический код вырожден, то при замене триплетов, кодирующий одну и ту же аминокислоту, мутации не проявляются. Другой вид мутаций – нонсенс - мутации. При этих мутациях при замене одного нуклеотида другим образуются бессмысленные триплеты. Синтез полипептида прекращается и белок имеет совсем иные свойства. УАГ. УАА. УГА. бессмысленные триплеты. Делеция или вставка одного или нескольких нуклеотидов ведут за собой утрату или вставку одной или нескольких аминокислот в полипептиде. эффекта может не быть. Если происходят делеция или вставка 1 нуклеотида (или другого числа нуклеотидов не кратного 3), наблюдается сдвиг рамки считывания, при этом нарушается структура полипептида. Большинство изменений молекулярной структуры генов приводит к новым формам считывания генетической информации, которая реализуется в метаболических путях и биохимических реакциях, появляются новые свойства клеток и всего организма. В организме происходит большое количество мутаций. Они затрагивают интеллект, поведение, метаболические признаки и т.д. мутации, изменяющие видимые морфологические признаки – видимые (мутация альбинизма). Нормальный доминантный ген превращается в рецессивный, выработка меланина прекращается, фенотипически проявляется белой окраской волос и глаз. Есть группа биохимических мутаций, которые выявляются с помощью сложных биохимических способов. Например, у человека синтезируется ряд ферментов, осуществляющих превращение лактозы в галактозу. При отсутствии фермента-лактазы происходит брожение в толстом кишечнике, газообразование и др. могут быть детская и взрослая формы. При накоплении галактозы – галактоземия, которая может привести к умственной отсталости. Мутации, нарушающие жизнь – летальные, полулетальные и сублетальные. Летальные – гибель зиготы или развившегося организма на определенной стадии эмбриогенеза – выкидыши. Полулетальные и сублетальные ослабляют жизнеспособность организма или отдельных клеток (например, брахидактилия – гомозиготы погибают). Хромосомные мутации (хромосомные абберации) – структурные перестройки, затрагивающие одну или несколько хромосом. При всем многообразии структурных перестроек все они связаны с потерей либо с добавлением участка хромосомы. Частичные моносомии и трисомии (смотри 8 лекцию). На долю хромосомных мутаций приходится 7% хромосомных болезней. Клинически они сопровождаются множественными пороками развития и аномалиями. Геномные мутации. Полиплоидия – увеличение числа хромосом, кратное диплоидному набору (клетки печени в норме). Анеуплоидия (гетероплоидия)- уменьшение или увеличение количества хромосом не кратное диплоидному. Гаплоидия – наличие гаплоидного набора хромосом в некоторых клетках (как правило, происходит гибель клеток). Мутации могут быть полезными, вредными или не оказывать явного влияния – т.е. быть нейтральными. Обычные гены в популяции адаптивны, обладатели лучше приспосабливаются, а вновь возникающие мутации чаще всего уже встречались ранее и были утрачены, потому что не способствовали лучшему приспособлению к определенным условиям жизни. Мутантный ген может накапливаться, может быть полезным. И все же большинство мутаций – вредны. Генные мутации. Механизмы генных мутаций. По характеру изменений в составе гена различают следующие типы мутаций: Делеции — утрата сегмента ДНК размером от одного нуклеотида до гена. Дупликации — удвоение или повторное дублирование сегмента ДНК от одного нуклеотида до целых генов. Инверсии — поворот на 180° сегмента ДНК размером от двух нуклеотидов до фрагмента, включающего несколько генов. Инсерции — вставка фрагментов ДНК размером от одного нуклеотида до целого гена. Трансверсии — замена пуринового основания на пиримидиновое или наоборот в одном из кодонов. Транзиции — замена одного пуринового основания на другое пуриновое или одного пиримидинового на другое в структуре кодона. По последствиям генных мутаций их классифицируют на нейтральные, регу-ляторные и динамические, а также на миссенс- и нонсенс-мутации. Нейтральная мутации (молчащая мутация) — мутация не имеет фенотипи-ческого выражения (например, в результате вырожденности генетического кода). Миссенс-мутация — замена нуклеотида в кодирующей части гена — приводит к замене аминокислоты в полипептиде. Нонсенс-мутация — замена нуклеотида в кодирующей части гена — приводит к образованию кодона-терминатора (стоп-кодона) и прекращению трансляции. Регуляторная мутация — мутация в 5'- или З'-нетранслируемых областях гена, такая мутация нарушает экспрессию гена. Динамические мутации — мутации, обусловленные увеличением числа три-нуклеотидных повторов в функционально значимых частях гена. Такие мутации могут привести к торможению или блокаде транскрипции, приобретению белковыми молекулами свойств, нарушающих их нормальный метаболизм. 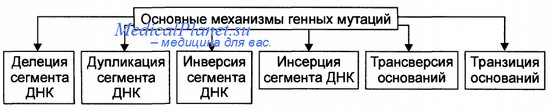 Генные мутации. затрагивают структуру самого гена. Мутации могут изменять участки молекулы ДНК различной длины. Наименьший участок, изменение которого приводит к появлению мутации, назван мутоном. Его может составить только одна пара нуклеотидов. Изменение последовательности нуклеотидов в ДНК обусловливает изменение в последовательности триплетов и е конечном итоге изменяет программу синтеза белка. Следует помнить, что нарушения в структуре ДНК приводят к мутациям только тогда, когда не осуществляется репарация. Мутации, возникающие под влиянием специальных воздействий — ионизирующей радиации, химических веществ, температурных факторов и т. п. — называются индуцированными, В свою очередь спонтанными называют мутации» возникшие без преднамеренного воздействия, под влиянием факторов внешней среды или вследствие биохимических и физиологических изменений в организме. Виды мутаций. Классификация мутаций. Многочисленные виды мутаций перечислены на рисунке. Спонтанные мутации возникают под влиянием естественных мутагенов экзо-или эндогенного происхождения, без специального (целенаправленного) вмешательства человека. Спонтанные мутации возникают, например, в результате действия химических веществ, образующихся в процессе метаболизма; воздействия естественного фона радиации или УФ-излучения; ошибок репликации и т.д. Индуцированные мутации вызваны направленным воздействием факторов внешней или внутренней среды. Индуцированный мутационный процесс может быть контролируемым и неконтролируемым. - Контролируемые мутации возникают, например, в эксперименте с целью изучения механизмов мутагенеза и/или его последствий. - Неконтролируемые мутации, например при случайном выбросе радиоактивных элементов в среду обитания. Виды мутаций. Классификация мутаций. Гаметические мутации — мутации в половых клетках. Они наследуются потомками и, как правило, обнаруживаются во всех клетках организма. Соматические мутации — мутации в неполовых (соматических) клетках — проявляются у того индивида, у которого они возникают. Они передаются при делении только дочерним клеткам и не наследуются следующим поколением индивида. Мозаики. Если соматическая мутация возникает на ранних стадиях дробления зиготы (но не первого деления) возникают клеточные линии с различными генотипами. Чем раньше в онтогенезе происходит соматическая мутация, тем больше клеток содержит данную мутацию. Подобные организмы получили название мозаичных. Патогенные мутации приводят к гибели эмбриона (или плода) и к развитию наследственных и врождённых заболеваний. Нейтральные мутации обычно не влияют на жизнедеятельность организма (например, мутации, вызывающие веснушки, изменение цвета волос, радужной оболочки глаза). Благоприятные мутации повышают жизнеспособность организма или вида (например, тёмная окраска кожных покровов у жителей африканского континента). Генные (точковые) мутации — любые изменения молекулярной структуры ДНК. Некоторые мутации не оказывают влияния на функцию соответствующего полипептида (например, замена нуклеотидов, не приводящая к замене аминокислоты в силу вырожденности генетического кода). Значительная часть точковых мутаций нарушает функционирование гена и приводит к развитию генных (моногенных) болезней. Фенотипически генные болезни проявляются признаками нарушений метаболизма (например, фенилкетонурия, гемофилии, нейрофиброматоз, муковисцидоз, мышечная дистрофия Дюшенна—Беккера, гемоглобинопатия S). Источник: http://medicalplanet.su/Patfiz/45.html MedicalPlanet 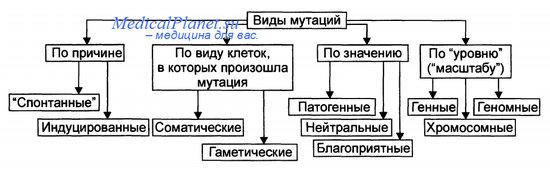 9. Хромосомы – структурные компоненты ядра. Структурная организация хроматина. Морфология хромосом. Нуклеосомная модель строения хромосом. Этапы упаковки хромосом. Понятие о кариотипе. Правила хромосомных наборов. В. Вальдейер, 1888 г. – предложил термин хромосома Т. Бовери и У.Сэттон – 1902 – 1907 гг. – сделали вывод, что хромосомы являются материальными носителями наследственной информации 80е гг. XIX века – описан митоз – деление соматических клеток и мейоз – деление половых клеток Вильсон, 1905- 1906 гг. - открыл половые хромосомы Т. Морган и его сотрудники А Стертевант ,. Г. Мёллер , К. Бриджес в 1910 г. открыли сцепленное наследование и доказали , что хромосома состоит из генов, расположенных в линейном порядке. Они сформулировали хромосомную теорию наследственности Морфология хромосом: В ходе клеточного цикла облик хромосомы меняется. В интерфазе это очень нежные структуры, занимающие в ядре отдельные хромосомные территории, но не заметные как обособленные образования при визуальном наблюдении. В митозе хромосомы преобразуются в плотно упакованные элементы, способные сопротивляться внешним воздействиям, сохранять свою целостность и форму[9][10]. Именно хромосомы на стадии профазы, метафазы или анафазы митоза доступны для наблюдения с помощью светового микроскопа. Митотические хромосомы можно увидеть у любого организма, клетки которого способны делиться митозом, исключение составляют дрожжи S.cerevisiae, чьи хромосомы слишком малы[11]. Обычно митотические хромосомы имеют размеры в несколько микрон. Например, самая большая хромосома человека хромосома 1 имеет длину около 7 — 8 мкм в метафазе и 10 мкм в профазе митоза. На стадии метафазы митоза хромосомы состоят из двух продольных копий, которые называются сестринскими хроматидами и которые образуются при репликации. У метафазных хромосом сестринские хроматиды соединены в районе первичной перетяжки, называемойцентромерой. Центромера отвечает за расхождение сестринских хроматид в дочерние клетки при делении. На центромере происходит сборкакинетохора — сложной белковой структуры, определяющей прикрепление хромосомы к микротрубочкам веретена деления — движителям хромосомы в митозе[13]. Центромера делит хромосомы на две части, называемые плечами. У большинства видов короткое плечо хромосомы обозначают буквой p, длинное плечо — буквой q. Длина хромосомы и положение центромеры являются основными морфологическими признаками метафазных хромосом. В зависимости от расположения центромеры различают три типа строения хромосом: акроцентрические хромосомы, у которых центромера находится практически на конце, и второе плечо настолько мало, что его может быть не видно на цитологических препаратах; субметацентрические хромосомы с плечами неравной длины; метацентрические хромосомы, у которых центромера расположена посередине или почти посередине В основе Парижской классификации хромосом человека (1971 г.) лежат методы специальной дифференциальной их окраски, при которой в каждой хромосоме выявляется характерный только для нее порядок чередования поперечных светлых и темных сегментов. Различные типы сегментов обозначают по методам, с помощью которых они выявляются наиболее четко. Например. Q-сегменты - это участки хромосом, флюоресцирующие после окрашивания акрихин-ипритом; G-сегменты выявляются при окрашивании красителем Гимза (Q- и G-сегменты идентичны); R-сегменты окрашиваются после контролируемой тепловой денатурации и т.д. Данные методы позволяют четко дифференцировать хромосомы человека внутри групп. Короткое плечо хромосом обозначают латинской буквой р. а длинное -q. Каждое плечо хромосомы разделяют на районы, нумеруемые по порядку от центромеры к теломере. В некоторых коротких плечах выделяют один такой район, а в других (длинных) - до четырех. Полосы внутри районов нумеруются по порядку от центромеры. Если локализация гена точно известна, для ее обозначения используют индекс полосы. Например, локализация гена, кодирующего эстеразу D. обозначается 1 Зр 14 - четвертая полоса первого района короткого плеча тринадцатой хромосомы. Локализация генов не всегда известна до полосы. Так, расположение гена ретинобластомы обозначают 13q, что означает локализацию его в длинном плече тринадцатой хромосомы. 10. Хромосомные мутации, их классификация. Причины и механизмы возникновения хромосомных мутаций. Роль хромосомных мутаций в развитии патологии человека и эволюционном процессе. Хромосомные мутации (аберрации) характеризуются изменением структуры отдельных хромосом. При них последовательность нуклеотидов в генах обычно не меняется, но изменение числа или положения генов при аберрациях может привести к генетическому дисбалансу, что пагубно сказывается на нормальном развитии организма. Виды аберраций и их механизмы представлены на рисунке. Различают внутрихромосомные, межхромосомные и изохромосомные аберрации. Внутрихромосомные аберрации — аберрации в пределах одной хромосомы. К ним относятся делеции, инверсии и дупликации. Делеция — утрата одного из участков хромосомы (внутреннего или терминального), что может стать причиной нарушения эмбриогенеза и формирования множественных аномалий развития (например, делеция в регионе короткого плеча хромосомы 5, обозначаемая как 5р-, приводит к недоразвитию гортани, ВПР сердца, отставанию умственного развития). Этот симптомокомплекс обозначен как синдром кошачьего крика, поскольку у больных детей из-за аномалии гортани плач напоминает кошачье мяуканье. Инверсия — встраивание фрагмента хромосомы на прежнее место после поворота на 180°. В результате нарушается порядок расположения генов. Дупликация — удвоение (или умножение) какого-либо участка хромосомы (например, трисомия по короткому плечу хромосомы 9 приводит к появлению множественных ВПР, включая микроцефалию, задержку физического, психического и интеллектуального развития). Межхромосомные аберрации — обмен фрагментами между негомологичными хромосомами. Они получили название транслокаций. Различают три варианта транслокаций: реципрокные (обмен фрагментами двух хромосом), нереципрокные (перенос фрагмента одной хромосомы на другую), робертсоновские (соединение двух акроцентрических хромосом в районе их центромер с потерей коротких плеч, в результате образуется одна метацентри-ческая хромосома вместо двух акроцентрических). Изохромосомные аберрации — образование одинаковых, но зеркальных фрагментов двух разных хромосом, содержащих одни и те же наборы генов. Это происходит в результате поперечного разрыва хроматид через центромеры (отсюда другое название — центрическое соединение). 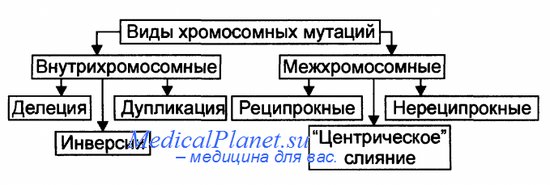 Хромосомные мутации. Мутации этого типа, называемые также хромосомными перестройками, или аберрациями, возникают в результате значительных изменений в структуре хромосом. Механизмом возникновения хромосомных перестроек являются образовавшиеся при мутагенном воздействии разрывы хромосом, последующая утрата некоторых фрагментов и воссоединение оставшихся частей хромосомы в ином порядке по сравнению с нормальной хромосомой. Хромосомные перестройки могут быть обнаружены с помощью светового микроскопа. Главные из них: нехватки, делении, дупликации, инверсии, транслокации и транспозиции. Внутрихромосомные: 1) Делеция – утрата участка хромосомы В гомозиготном состоянии =» летальные мутации В гетерозиготном состоянии =» тяжелые нарушения развития Синдром кошачьего крика – делеция ½ короткого плеча 5-ой хромосомы (маленький головной мозг, мяукающий плач, продолжительность жизни 5 лет) Синдром Орбели – делеция длинного плеча 13ой хромосомы (тяжелые соматические и умственные расстройства) 2) Дупликация – удвоение участка хромосомы У человека описаны по всем аутосомам Не происходит утраты генетического материала Синдром 9р+ (задержка роста, короткая шея, птоз, умственная отсталость) 3) Инверсия – поворот участка хромосомы на 180о количество генетического материала не изменяется, но изменяется порядок сцепления генов Инверсия длинного плеча 21-ой хромосомы – признаки синдрома Дауна. Инверсия длинного плеча Х-хромосомы – признаки синдрома Шерешевского – Тернера. 4) Транспозиция– перемещение участка хромосомы либо внутри хромосомы, либо в другую хромосомы. Играют регуляторную роль: включают или выключают ген в работу Межхромосомные мутации – происходит обмен участками негомологичных хромосом. Реципрокные транслокации – взаимный обмен участками двух негомологичных хромосом Умственно отсталые дети Транслокация в соматических клетках 9 и 22 хромосомы =» развитие миелоидного лейкоза (филадельфийская ph-хромосома укорочена =» синтез окнобелков) Нереципрокные транслокации – взаимного обмена не происходит. Потеря части коротких плеч двух негомологичных хромосом и их соединения. Транслокация целой хромосомы в другую негомологичную хромосому (рождение детей с транслокацией синдрома Дауна) Робертсоновские транслокации – слияние негомологичных акроцентрических хромосом в области центромеры Фенотипически здоровый человек, так как имеют сбалансированный генотип, но велика вероятность рождения детей-Даунов. Хромосомные перестройки играют определенную роль в эволюционном процессе и видообразовании, в нарушении фертильности, в онкологических и врождённых наследственных заболеваниях человека. 11. Митотический (пролиферативный) цикл клетки. Фазы митотического цикла, их характеристика и значение. Главные механизмы пролиферативного цикла. Регуляция митоза. Амитоз. Эндомитоз, политения, их значение. Жизненный цикл клетки – это периоды существования клетки от момента ее образования путем деления материнской клетки до собственного деления или смерти. Возможны три варианта жизненного цикла: 1. Клетки готовятся к делению и заканчивают свою жизнь митозом (митотический цикл) 2. Клетки дифференцируются, функционируют и погибают. 3. Клетки переходят в период G0. При определенных условиях могут выйти либо в митотический цикл, либо терминальную дифференцировку. Обязательный компонент клеточного цикла, митотический цикл – комплекс взаимодействующих и согласованных во времени событий, происходящих в процессе подготовки клетки к делению, и на протяжении всего деления. Продолжительность от 10-50 часов. Биологическое значение: обеспечивает преемственность хромосом в ряду клеточных поколений, образования клеток, равноценных по объему и содержанию наследственной информации –механизм воспроизведения клеточной организации эукариотического типа в индивидуальном развитии. Периоды: 1. Пресинтетический период (G) 2n2с: самый длинный период. Начинается рост клеток, синтез белков и РНК, которые начинаются после митоза. Происходит подготовка клеток к синтезу ДНК (S-периоду). Завершается формирование ядрышка. Из цитоплазмы в ядро поступает до 90% белка. Рост массы клетки. 2. Синтетический период (S) 2n4с: удваивается количество наследственного материала клетки – молекул ДНК. Каждая хромосома становится двух – хроматидной. Происходит репликация молекул ДНК, синтез белков – гистонов, с которыми связана каждая нить ДНК. Синтез РНК увеличивается соответственно количеству ДНК. При репликации второй молекулы ДНК спирали раскручиваются, рвутся водородные связи, каждая становится матрицей для воспроизводства новой цепей ДНК. Синтез новых молекул ДНК осуществляется при участии ферментов. Каждая из двух дочерних молекул обязательно включает одну старую и одну новую спираль – этополуконсервативный способ репликации. Начинается удвоение центриолей. Продолжительность 7-12 часов. Удвоение ДНК и хромосом: репликация ДНК начинается одновременно во многих местах точках инициации. Происходит прикрепление комплекса ферментов (репликационная машина), ДНК освобождается от гистонов и расплетается – репликационный глазок.Разделение исходных матричных и синтез новых дочерних цепей ДНК в глазке происходит одновременно в обе стороны в репликационных вилках. После удвоения ДНК с ними соединяются гистоны, и хромосома становится двойной, состоящей из двух хроматид, которые соединены в области центромеры. 3. Постсинтетический период (G2) 2n4c: отрезок времени от окончания синтетического периода до начала митоза. Интенсивный синтез РНК и белка, накопление АТФ (для деления). Завершается удвоение центриолей, митохондрий, пластид, синтезируются белки из которых строится веретено деления. 4. Митоз – это непрямое деление клетки. Состоит из четырех фаз: 1. Профаза (2n4c): увеличение объема ядра. Хромосомы спирализуются и приобретают вид нитей. Ядрышко разрушается. Распад ядерной оболочки. В цитоплазме уменьшается количество структур шероховатой сети. Резко сокращается число полисом. Центриоли клеточного центра расходятся к полюсам клетки, между ними микротрубочки образуют веретено деления. Самая длинная фаза.  2. Метафаза (2n4c): заканчивается образование веретена деления. Хромосомы выстраиваются в экваторной плоскости клетки (метафазная пластинка). Микротрубочки веретена деления связаны с кинетахорами хромосом. Каждая хромосома расщепляется на две хроматиды (дочерние хромосомы), соединенные в области кинетохора. Самая короткая. 3. Анафаза (4n4c): связь между хроматидами нарушаются, и они в качестве самостоятельных хромосом перемещаются к полюсам клетки. По завершению движения на полюсах собираются два полных набора хромосом (диплоидных). Короткая фаза. 4. Телофаза (2n2c): реконструируются интерфазные ядра дочерних клеток. Хромосома деспирализуется, образуются ядрышки. Разрушено веретено деления. Материнская клетка делится на две дочерние. Значение митоза: точное распределение генетического материала между двумя дочерними клетками. Обе дочерние клетки получают диплоидный набор хромосом. Обеспечивает постоянство числа хромосом, клеточный механизм роста, развитие организма, регенерации, бесполого размножения. Механизмы пролиферативного цикла, обеспечивающие равномерное распределение генетического материала. Главные события митотического цикла заключаются в редупликации (самоудвоении) наследственного материала материнской клетки и в равномерном распределении этого материала между дочерними клетками.Указанным событиям сопутствуют закономерные изменения химической и морфологической организации хромосом — ядерных структур, в которых сосредоточено более 90% генетического материала эукариотической клетки (основная часть внеядерной ДНК животной клетки находится в митохондриях) Регуляция митоза. В организме М. контролируются системой нейрогуморальной регуляции, котораяосуществляется нервной системой, гормонами надпочечников, гипофиза, щитовидной и половых желёз, атакже местными факторами (продукты тканевого распада, функциональная активность клеток).Взаимодействие различных регуляторных механизмов обеспечивает как общие, так и местные изменениямитотической активности. М. опухолевых клеток выходят из-под контроля нейрогуморальной регуляции. Выражением регуляции М. в связи с взаимодействием организма и среды служит суточный ритмделения клеток. В большинстве органов ночных животных максимум М. отмечается утром, а минимум — вночное время. У дневных животных и человека отмечается обратная динамика суточного ритма. Суточныйритм М. — следствие цепной реакции, в которую вовлекаются ритмические изменения внешней среды(освещённость, температура, режим питания и др.), ритм функциональной активности клеток и измененияпроцессов обмена веществ (см. Биологические ритмы). Нетипичные формы митоза К нетипичным формам митоза относятся амитоз, эндомитоз, политения. 1. Амитоз – это прямое деление ядра. При этом сохраняется морфология ядра, видны ядрышко и ядерная мембрана. Хромосомы не видны, и их равномерного распределения не происходит. Ядро делится на две относительно равные части без образования митотического аппарата (системы микротрубочек, центриолей, структурированных хромосом). Если при этом деление заканчивается, возникает двухъядерная клетка. Но иногда перешнуровывается и цитоплазма. 2. Эндомитоз. При этом типе деления после репликации ДНК не происходит разделения хромосом на две дочерние хроматиды. Это приводит к увеличению числа хромосом в клетке иногда в десятки раз по сравнению с диплоидным набором. Так возникают полиплоидные клетки. В норме этот процесс имеет место в интенсивно функционирующих тканях, например, в печени, где полиплоидные клетки встречаются очень часто. Однако с генетической точки зрения эндомитоз представляет собой геномную соматическую мутацию. 3. Политения. Происходит кратное увеличение содержания ДНК (хромонем) в хромосомах без увеличения содержания самих хромосом. При этом количество хромонем может достигать 1000 и более, хромосомы при этом приобретают гигантские размеры. При политении выпадают все фазы митотического цикла, кроме репродукции первичных нитей ДНК. Такой тип деления наблюдается в некоторых высокоспециализированных тканях (печеночных клетках, клетках слюнных желез двукрылых насекомых). Политенные хромосомы дрозофил используются для построения цитологических карт генов в хромосомах. 12. Мейоз как процесс формирования гаплоидных клеток. Фазы мейоза, их характеристика и значение. Рекомбинация наследственного материала, ее медицинское и эволюционное значение. При половом размножении дочерний организм возникает в результате слияния двух половых клеток (гамет) и последующего развития из оплодотворенной яйцеклетки —зиготы. Половые клетки родителей обладают гаплоидным набором (n) хромосом, а в зиготе при объединении двух таких наборов число хромосом становится диплоидным (2n): каждая пара гомологичных хромосом содержит одну отцовскую и одну материнскую хромосому. Гаплоидные клетки образуются из диплоидных в результате особого клеточного деления — мейоза. Мейоз — разновидность митоза, в результате которого из диплоидных (2п) соматических клеток половых желез образуются гаплоидные гаметы (1n). При оплодотворении ядра гаметы сливаются, и восстанавливается диплоидный набор хромосом. Таким образом, мейоз обеспечивает сохранение постоянного для каждого вида набора хромосом и количества ДНК. Мейоз представляет собой непрерывный процесс, состоящий из двух последовательных делений, называемых мейозом I и мейозом II. В каждом делении различают профазу, метафазу, анафазу и телофазу. В результате мейоза I число хромосом уменьшается вдвое (редукционное деление): при мейозе II гаплоидность клеток сохраняется (эквационное деление). Клетки, вступающие в мейоз, содержат генетическую информацию 2n2хр (рис. 1). В профазе мейоза I происходит постепенная спирализация хроматина с образованием хромосом. Гомологичные хромосомы сближаются, образуя общую структуру, состоящую из двух хромосом (бивалент) и четырех хроматид (тетрада). Соприкосновение двух гомологичных хромосом по всей длине называется конъюгацией. Затем между гомологичными хромосомами появляются силы отталкивания, и хромосомы сначала разделяются в области центромер, оставаясь соединенными в области плеч, и образуют перекресты (хиазмы). Расхождение хроматид постепенно увеличивается, и перекресты смещаются к их концам. В процессе конъюгации между некоторыми хроматидами гомологичных хромосом может происходить обмен участками — кроссинговер, приводящий к перекомбинации генетического материала. К концу профазы растворяются ядерная оболочка и ядрышки, формируется ахроматиновое веретено деления. Содержание генетического материала остается прежним (2n2хр). В метафазе мейоза I биваленты хромосом располагаются в экваториальной плоскости клетки. В этот момент спирализация их достигает максимума. Содержание генетического материала не изменяется (2п2хр). В анафазе мейоза I гомологичные хромосомы, состоящие из двух хроматид, окончательно отходят друг от друга и расходятся к полюсам клетки. Следовательно, из каждой пары гомологичных хромосом в дочернюю клетку попадает только одна — число хромосом уменьшается вдвое (происходит редукция). Содержание генетического материала становится 1n2хр у каждого полюса. В телофазе происходит формирование ядер и разделение цитоплазмы — образуются две дочерние клетки. Дочерние клетки содержат гаплоидный набор хромосом, каждая хромосома — две хроматиды (1n2хр). Интеркинез — короткий промежуток между первым и вторым мейотическими делениями. В это время не происходит репликации ДНК, и две дочерние клетки быстро вступают в мейоз II, протекающий по типу митоза. 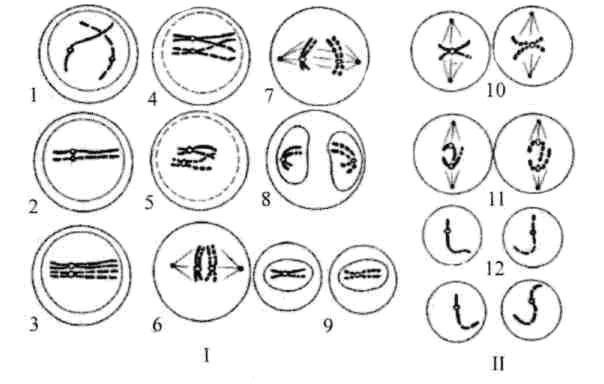 Рис. 1. Схема мейоза (показана одна пара гомологичных хромосом). Мейоз I: 1, 2, 3. 4. 5 — профаза; 6 —метафаза; 7 — анафаза; 8 — телофаза; 9 — интеркинез. Мейоз II; 10 —метафаза; II —анафаза; 12 — дочерние клетки. В профазе мейоза II происходят тс же процессы, что и в профазе митоза. В метафазе хромосомы располагаются в экваториальной плоскости. Изменений содержания генетического материала не происходит (1n2хр). В анафазе мейоза II хроматиды каждой хромосомы отходят к противоположным полюсам клетки, и содержание генетического метериала у каждого полюса становится lnlxp. В телофазе образуются 4 гаплоидные клетки (lnlxp). Таким образом, в результате мейоза из одной диплоидной материнской клетки образуются 4 клетки с гаплоидным набором хромосом. Кроме того, в профазе мейоза I происходит перекомбинация генетического материала (кроссинговер), а в анафазе I и II — случайное отхождение хромосом и хроматид к одному или другому полюсу. Эти процессы являются причиной комбинативной изменчивости. Биологическое значение мейоза: 1) является основным этапом гаметогенеза; 2) обеспечивает передачу генетической информации от организма к организму при половом размножении; 3) дочерние клетки генетически не идентичны материнской и между собой. Атак же, биологическое значение мейоза заключается в том, что уменьшение числа хромосом необходимо при образовании половых клеток, поскольку при оплодотворении ядра гамет сливаются. Если бы указанной редукции не происходило, то в зиготе (следовательно, и во всех клетках дочернего организма) хромосом становилось бы вдвое больше. Однако это противоречит правилу постоянства числа хромосом. Благодаря мейозу половые клетки гаплоидны, а при оплодотворении в зиготе восстанавливается диплоидный набор хромосом (рис. 2 и 3). 13. Геномные мутации, причины и механизмы их возникновения. Мутагены и их классификация. Классификация геномных мутаций. Значение геномных мутаций. Антимутационные механизмы. Геномные мутации. Виды геномных мутаций. Геномные мутации характеризуются изменением числа хромосом. У человека известны полиплоидия (в том числе тетраплоидия и триплоидия) и анеуплоидия. Полиплоидия — увеличение числа наборов хромосом, кратное гаплоидному (Зn, 4n, 5n и т.д.). Причины: двойное оплодотворение и отсутствие первого мейотического деления. У человека полиплоидия, а также большинство анеуплоидий приводят к формированию леталей. Анеуплоидия — изменение (уменьшение — моносомия, увеличение — трисомия) числа хромосом в диплоидном наборе, т.е. не кратное гаплоидному (2n+1, 2n-1 и т.д.). Механизмы возникновения: нерасхождение хромосом (хромосомы в анафазе отходят к одному полюсу, при этом на каждую гамету с одной лишней хромосомой приходится другая — без одной хромосомы) и «анафазное отставание» (в анафазе одна из передвигаемых хромосом отстаёт от всех других). Трисомия — наличие трёх гомологичных хромосом в кариотипе (например, по 21-й паре, что приводит к развитию синдрома Дауна; по 18-й паре — синдрома Эдвардса; по 13-й паре — синдрома Патау). Моносомия — наличие только одной из двух гомологичных хромосом. При моносомии по любой из аутосом нормальное развитие эмбриона невозможно. Единственная совместимая с жизнью моносомия у человека — по хромосоме X — приводит к развитию синдрома Шерешевского—Тернера (45,Х0).  1.3. Раздел III. Организменный (онтогенетический) уровень организации биологических систем 1. Размножение организмов. Бесполое и половое размножение. Формы бесполого размножения, его сущность, биологическое значение. Половое размножение, его эволюционное значение. Размножение — свойство живых организмов воспроизводить себе подобных. Существуют два основных способа размножения — бесполое и половое. |