Биология экзамен. 1 Раздел I. Общая характеристика жизни
 Скачать 0.75 Mb. Скачать 0.75 Mb.
|

КомплиментарностьКомплементарное (дополнительное) действие генов — это вид взаимодействия неаллельных генов, доминантные аллели которых при совместном сочетании в генотипе обусловливают новое фенотипическое проявление признаков. При этом расщепление гибридов F2 по фенотипу может происходить в соотношениях 9:6:1, 9:3:4, 9:7, иногда 9:3:3:1. Примером комплементарности является наследование формы плода тыквы. Наличие в генотипе доминантных генов А или В обусловливает сферическую форму плодов, а рецессивных — удлинённую. При наличии в генотипе одновременно доминантных генов А и В форма плода будет дисковидной. При скрещивании чистых линий с сортами, имеющими сферическую форму плодов, в первом гибридном поколении F1 все плоды будут иметь дисковидную форму, а в поколении F2 произойдёт расщепление по фенотипу: из каждых 16 растений 9 будут иметь дисковидные плоды, 6 — сферические и 1 — удлинённые. ЭпистазПодавление (ингибирование) действия одной аллельной пары генов геном другой, не аллельной им пары, называется эпистазом. Различают доминантный и рецессивный эпистаз. Если обычное аллельное доминирование можно представить в виде формулы А>а, То явление эпистаза выразится формулой А>В (доминантный эпистаз) или А>В (рецессивный эпистаз), когда доминантный или рецессивный ген одной аллельной пары не допускает проявления генов другой аллельной пары. Гены, подавляющие действие других, не аллельных им генов, называются Эпистатичными, А подавляемые — Гипостатичными. Эпистатическое взаимодействие генов по своему характеру противоположно комплементарному взаимодействию. При эпистазе фермент, образующийся под контролем одного гена, полностью подавляет или нейтрализует действие фермента, контролируемого другим геном. Разберем эпистатическое действие генов на примере наследования окраски зерна у овса (рис. 28). У этой культуры были установлены доминантные гены, определяющие черную и серую окраску зерна. Обозначим один из них буквой А, А второй — В. При этом можно представить себе скрещивание, в котором родительские формы имели генотипы A Abb (черносемянный) и АаВВ (серосемянный). В генотипе растения первого поколения (АаВB) Содержатся доминантные гены и черной окраски А, И серой окраски В. Так как ген А Эпистатичен по отношению к гену В, Он не дает ему проявиться, и все гибриды F1 Будут черносемянными. В F1 Произойдет расщепление в отношении 12 черных : 3 серых: 1 белый. Такой результат расщепления легко понять, если представить себе отношение 12:3:1 как видоизменение типичного для дигибридных скрещиваний отношения 9:3:3:1. В девяти сочетаниях присутствуют оба доминантных гена А И В, Но ген серой окраски В Не может проявляться, и они дают черносемянные растения. В трех сочетаниях (AAbbAabb, Aabb) Ген черной окраски семян А Также обусловит развитие черносемянных растений. Эта группа по фенотипу будет совершенно сходна с первой, и, следовательно, из каждых 16 растений 12 будут черносемянными. В трех сочетаниях (ааВВ, ааВB, ааВB) Доминантный ген В При отсутствии эпистатичного гена А Может проявить доминантное действие по отношению к своему рецессивному аллелю b, И разовьются растения с серыми семенами. Один генотип (Aabb) Представляет собой новую комбинацию, в которой проявится белая окраска зерна, так как отсутствуют оба доминантных гена. ПолимерияПолимерия — взаимодействие неаллельных множественных генов, однонаправленно влияющих на развитие одного и того же признака; степень проявления признака зависит от количества генов. Полимерные гены обозначаются одинаковыми буквами, а аллели одного локуса имеют одинаковый нижний индекс. Полимерное взаимодействие неаллельных генов может быть кумулятивным и некумулятивным. При кумулятивной (накопительной) полимерии степень проявления признака зависит от суммарного действия нескольких генов. Чем больше доминантных аллелей генов, тем сильнее выражен тот или иной признак. Расщепление в F2 по фенотипу при дигибридном скрещивании происходит в соотношении 1:4:6:4:1, а в целом соответствует третьей, пятой (при дигибридном скрещивании), седьмой (при тригибридном скрещивании) и т.п. строчкам в треугольнике Паскаля. При некумулятивной полимерии признак проявляется при наличии хотя бы одного из доминантных аллелей полимерных генов. Количество доминантных аллелей не влияет на степень выраженности признака. Расщепление в F2 по фенотипу при дигибридном скрещивании — 15:1. Пример полимерии — наследование цвета кожи у людей, который зависит (в первом приближении) от четырёх генов с кумулятивным эффектом. 6. Сцепленное наследование. Группы сцепления. Хромосомная теория наследственности. Результатом исследований Т. Моргана стало создание им хромосомной теории наследственности: гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов; набор генов каждой из негомологичных хромосом уникален; каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены; гены расположены в хромосомах в определенной линейной последовательности; гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов; сцепление генов может нарушаться в процессе кроссинговера, что приводит к образованию рекомбинантных хромосом; частота кроссинговера зависит от расстояния между генами: чем больше расстояние, тем больше величина кроссинговера; каждый вид имеет характерный только для него набор хромосом — кариотип. В 1906 году У. Бэтсон и Р. Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве, гибриды всегда повторяли признаки родительских форм. Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование. Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков. Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Т. Морган. Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила. Дрозофила каждые две недели при температуре 25 °С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее. Они имеют всего 8 хромосом в диплоидном наборе, достаточно легко размножаются в пробирках на недорогой питательной среде. 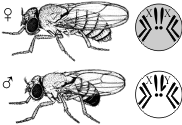 Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибриды, имеющие серое тело и нормальные крылья (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев, — над геном недоразвитых). При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% — серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% — черные длиннокрылые и 8,5% — серые с зачаточными крыльями). Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме. 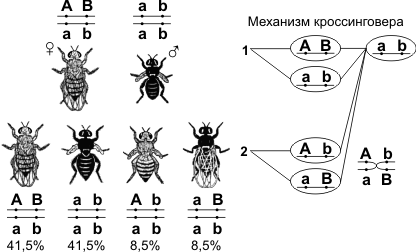 1 — некроссоверные гаметы; 2 — кроссоверные гаметы. Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и аb, а отцовский — один тип — аb. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и ааbb. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Ааbb и ааВb. Для того, чтобы объяснить это, необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генамиА и В, появляются гаметы Аb и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но, поскольку кроссинговер происходит при образовании небольшой части гамет, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1. Группа сцепления — гены, локализованные в одной хромосоме и наследующиеся совместно. Количество групп сцепления соответствует гаплоидному набору хромосом. Сцепленное наследование — наследование признаков, гены которых локализованы в одной хромосоме. Сила сцепления между генами зависит от расстояния между ними: чем дальше гены располагаются друг от друга, тем выше частота кроссинговера и наоборот. Полное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление — разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними. Независимое наследование — наследование признаков, гены которых локализованы в разных парах гомологичных хромосом. Некроссоверные гаметы — гаметы, в процессе образования которых кроссинговер не произошел.
Кроссоверные гаметы — гаметы, в процессе образования которых произошел кроссинговер. Как правило кроссоверные гаметы составляют небольшую часть от всего количества гамет.
Нерекомбинанты — гибридные особи, у которых такое же сочетание признаков, как и у родителей. Рекомбинанты — гибридные особи, имеющие иное сочетание признаков, чем у родителей. Расстояние между генами измеряется в морганидах — условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам. У дигетерозигот доминантные гены могут располагаться или в одной хромосоме (цис-фаза), или в разных (транс-фаза). 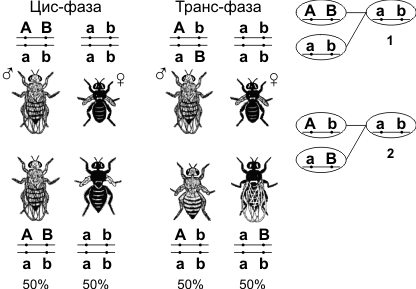 1 — Механизм цис-фазы (некроссоверные гаметы); 2 — механизм транс-фазы (некроссоверные гаметы). 7. Наследование пола и признаков, сцепленных с полом. Половые хромосомы и их роль в детерминации пола. 8. Формы изменчивости. Их значение в онтогенезе и в эволюции. Изменчивость — это возникновение индивидуальных различий. На основе изменчивости организмов появляется генетическое разнообразие форм, которые в результате действия естественного отбора преобразуются в новые подвиды и виды. Различают изменчивость модификационную, или фенотипическую, и мутационную, или генотипическую. ТАБЛИЦА Сравнительная характеристика форм изменчивости (Т.Л. Богданова. Биология. Задания и упражнения. Пособие для поступающих в ВУЗы. М.,1991)
Модификационная изменчивость Модификационная изменчивость не вызывает изменений генотипа, она связана с реакцией данного, одного и того же генотипа на изменение внешней среды: в оптимальных условиях выявляется максимум возможностей, присущих данному генотипу. Так, продуктивность беспородных животных в условиях улучшенного содержания и ухода повышается (надои молока, нагул мяса). В этом случае все особи с одинаковым генотипом отвечают на внешние условия одинаково (Ч. Дарвин этот тип изменчивости назвал определенной изменчивостью). Однако другой признак — жирность молока — слабо подвержен изменениям условий среды, а масть животного — еще более устойчивый признак. Модификационная изменчивость обычно колеблется в определенных пределах. Степень варьирования признака у организма, т. е. пределы модификационной изменчивости, называется нормой реакции. Широкая норма реакции свойственна таким признакам, как удои молока, размеры листьев, окраска у некоторых бабочек; узкая норма реакции — жирности молока, яйценоскости у кур, интенсивности окраски венчиков у цветков и др. Фенотип формируется в результате взаимодействий генотипа и факторов среды. Фенотипические признаки не передаются от родителей потомкам, наследуется лишь норма реакции, т. е. характер реагирования на изменение окружающих условий. У гетерозиготных организмов при изменении условий среды можно вызвать различные проявления данного признака. Свойства модификаций: 1) ненаследуемость; 2) групповой характер изменений; 3) соотнесение изменений действию определенного фактора среды; 4) обусловленность пределов изменччивоти генотипом. Генотипическая изменчивость Генотипическая изменчивость подразделяется на мутационную и комбинативную. Мутациями называются скачкообразные и устойчивые изменения единиц наследственности — генов, влекущие за собой изменения наследственных признаков. Термин “мутация” был впервые введен де Фризом. Мутации обязательно вызывают изменения генотипа, которые наследуются потомством и не связаны со скрещиванием и рекомбинацией генов. Классификация мутаций. Мутации можно объединять, в группы—классифицировать по характеру проявления, по месту или, по уровню их возникновения. Мутации по характеру проявления бывают доминантными и рецессивными. Мутации нередко понижают жизнеспособность или плодовитость. Мутации, резко снижающие жизнеспособность, частично или полностью останавливающие развитие, называют полулетальными а несовместимые с жизнью — летальными. Мутации подразделяют по месту их возникновения. Мутация, возникшая в половых клетках, не влияет на признаки данного организма, а проявляется только в следующем поколении. Такие мутации называют генеративными. Если изменяются гены в соматических клетках, такие мутации проявляются у данного организма и не передаются потомству при половом размножении. Но при бесполом размножении, если организм развивается из клетки или группы клеток, имеющих изменившийся — мутировавший — ген, мутации могут передаваться потомству. Такие мутации называют соматическими. Мутации классифицируют по уровню их возникновения. Существуют хромосомные и генные мутации. К мутациям относится также изменение кариотипа (изменение числа хромосом).. Полиплоидия — увеличение числа хромосом, кратное гаплоидному набору. В соответствии с этим у растений различают триплоиды (Зп), тетраплоиды (4п) и т. д. В растениеводстве известно более 500 полиплоидов (сахарная свекла, виноград, гречиха, мята, редис, лук и др.). Все они выделяются большой вегетативной массой и имеют большую хозяйственную ценность. Большое многообразие полиплоидов наблюдается в цветоводстве: если одна исходная форма в гаплоидном наборе имела 9 хромосом, то культивируемые растения этого вида могут иметь 18, 36, 54 и до 198 хромосом. Полиплоиды пблучают в результате воздействия на растения температуры, ионизирующей радиации, химических веществ (колхицин), которые разрушают веретено деления клетки. У таких растений гаметы диплоидны, а при слиянии с гаплоидными половыми клетками партнера в зиготе возникает триплоидный набор хромосом (2п + п = Зп). Такие триплоиды не образуют семян, они бесплодны, но высокоурожайны. Четные полиплоиды образуют семена. Гетероплоидия —изменение числа Хромосом, не кратное гаплоидному набору. При этом набор хромосом в клетке может быть увеличен на одну, две, три хромосомы (2п + 1; 2п + 2; 2п + 3) или уменьшен на одну хромосому (2л-1). Например, у человека с снндромом Дауна оказывается одна лишняя хромосома по 21-й паре и кариотип такого человека составляет 47 хромосом У людей с синдромом Шерешевского —Тернера (2п-1) отсутствует одна Х-хромосома и в кариотипе остается 45 хромосом. Эти и другие подобные отклонения числовых отношений в кариотипе человека сопровождаются расстройством здоровья, нарушением психики и телосложения, снижением жизнеспособности и др. Хромосомные мутации связаны с изменением структуры хромосом.Существуют следующие виды перестроек хромосом: отрыв различных участков хромосомы, удвоение отдельных фрагментов, поворот участка хромосомы на 180° или присоединение отдельного участка хромосомы к другой хромосоме. Подобное изменение влечет за собой нарушение функции генов в хромосоме и наследственных свойств организма, а иногда и его гибель. Генные мутации затрагивают структуру самого гена и влекут за собой изменение свойств организма (гемофилия, дальтонизм, альбинизм, окраска венчиков цветков и т. д.). Генные мутации возникают как в соматических, так и в половых клетках. Они могут быть доминантными и рецессивными. Первые проявляются как у гомозигот, так и. у гетерозигот, вторые — только у гомозигот. У растений возникшие соматические генные мутации сохраняются при вегетативном размножении. Мутации в половых клетках наследуются при семенном размножении растений и при половом размножении животных. Одни мутации оказывают на организм положительное действие, другие безразличны, а третьи вредны, вызывая либо гибель организма, либо ослабление его жизнеспособности (например, серповидноклеточная анемия, гемофилия у человека). При выведении новых сортов растений и штаммов микроорганизмов используют индуцированные мутации, искусственно вызываемые теми или иными мутагенными факторами (рентгеновские или ультрафиолетовые лучи, химические вещества). Затем проводят отбор полученных мутантов, сохраняя наиболее продуктивные. В нашей стране этими методами получено много хозяйственно перспективных сортов растений: неполегающие пшеницы с крупным колосом, устойчивые к заболеваниям; высокоурожайные томаты; хлопчатник с крупными коробочками и др. Свойства мутаций: 1. Мутации возникают внезапно, скачкообразно. 2. Мутации наследственны, т. е. стойко передаются из поколения в поколение. 3. Мутации ненаправденны - мутировать может любой локус, вызывая изменения как незначительных, так и жизненно важных признаков. 4. Одни и те же мутации могут возникать повторно. 5. По своему проявлению мутации могут быть полезными и вредными, доминантными и рецессивными. Способность к мутированию — одно из свойств гена. Каждая отдельная мутация вызывается какой-то причиной, но в большинстве случаев эти причины неизвестны. Мутации связаны с изменениями во внешней среде. Это убедительно доказывается тем, что путем воздействия внешними факторами удается резко повысить их число. |