Анатомия письменная. 1. Строение нервной ткани
 Скачать 0.71 Mb. Скачать 0.71 Mb.
|

ВЫПОЛНЕНИЕ ПРАКТИЧЕСКИХ ЗАДАНИЙ ПО ДИСЦИПЛИНЕ _______________Анатомия центральной нервной системы________ Группа Бу20П161 Студент М.Х. Мухаммадиева МОСКВА 2022 Практическое задание № 1 Тема 1. Строение нервной ткани. Нервная система. Задание № 1. Напишите небольшое эссе на одну из тем по выбору 3. Классификация, свойства и функции нейронов. Нейрон - это структурно-функциональная единица нервной ткани. Это специализированная клетка, которая, наряду с общими физиологическими свойствами (возбудимость, проводимость), обладает и рядом специфических свойств: - Воспринимать информацию - переводить информацию раздражителя на биологический язык клетки. - Обрабатывать информацию - т.е. проводить анализ информации, синтез - соединение различных частей информации после анализа с получением нового качества. - Кодировать информацию - превращать информацию в форму удобную для хранения в мозге. - Формировать командный управляющий сигнал, который распространяется на другие клетки нейроны, мышечные клетки. - Передача информации нейрона на другие структуры. Нейроны способны контактировать с другими клетками и оказывать на них информационное воздействие (место контактов - синапс). Все свои виды деятельности нейрон осуществляет за счѐт 3-х физиологических свойств (помимо возбудимости и проводимости): - Рецепция; - Электрогенез; - Нейросекреция. В общем плане, все нейроны имеют тело – сому и отростки – дендриты и аксоны. Их условно разделяют по структуре и функциям на следующие группы: - По форме тела: многоугольные, пирамидные, круглые, овальные. - По количеству и характеру отростков: Униполярные – имеющие один отросток Псевдоуниполярные – от тела отходит один отросток, который затем делится на 2 ветви. Биполярные – 2 отростка, один дендритоподобный, другой аксон. Мультиполярные – имеют 1 аксон и много дендритов. - По медиатору, выделяемому нейроном в синапсе: холинергические, адренергические, серотонинергические, пептидергические и т.д. - По функциям: Афферентные, или чувствительные - служат для восприятия сигналов из внешней и внутренней среды и передачи их в ЦНС. Вставочные, или интернейроны, промежуточные - беспечивают переработку, хранение и передачу информации к эфферентным нейронам. Их в ЦНС большинство. Эфферентные или двигательные - формируют управляющие сигналы, и передают их к периферическим нейронам и исполнительным органам. - По физиологической роли: возбуждающие и тормозные. Общими функциями нейронов ЦНС являются прием, кодирование, хранение информации и выработка нейромедиатора. Нейроны, с помощью многочисленных синапсов, получают сигналы в виде постсинаптических потенциалов. Затем перерабатывают эту информацию и формируют определенную ответную реакцию. Следовательно, они выполняют и интегративную, т.е. объединительную функцию. Связь между нейронами, как видно, осуществляется через промежуток между концами аксона одного нейрона и дендритами другого. Если они лежат в достаточной близости, т. е. промежуток мал, то в этом месте может образоваться синаптический узел, или синапс, связывающий эти два нейрона. Синапс подобен сопротивлению в электрической цепи. Если это сопротивление велико, то связь между нейронами слабая и возбуждение одного нейрона не вызывает возбуждения другого. Если же «сопротивление» синапса мало, то имеется сильная связь и нейрон без труда возбуждается от аксона другого нейрона, с ним связанного. Возбуждение нейрона происходит по принципу «все или ничего». Это значит, что нейрон может быть либо возбужден, и от клетки вдоль аксона к синаптическим узлам и далее к другим нейронам идет нервный импульс, либо не возбужден. Практическое задание № 2 Тема 6. Общее строение мозжечка Задание № 1. Напишите эссе по одной из следующих тем. 4. Ромбовидная ямка. Ромбовидная ямка, fossa rhomboidea, имеет соответственно ромбовидной форме четыре стороны — две верхние и две нижние. Верхние стороны ромба ограничены двумя верхними мозжечковыми ножками, а нижние стороны — двумя нижними ножками. Вдоль ромба, по средней линии, от верхнего угла к нижнему тянется срединная борозда, sulcus medianus, которая делит ромбовидную ямку на правую и левую половины. По сторонам борозды расположено парное возвышение, eminentia medialis, обусловленное скоплением серого вещества. Книзу eminentia medialis постепенно суживается, переходя в треугольник, на который проецируется ядро подъязычного нерва, trigonum nervi hypoglossi. Латеральнее нижней части этого треугольника лежит меньший треугольник, заметный по своей серой окраске, trigonum nervi vagi, в котором заложено вегетативное ядро блуждающего нерва, nucleus dorsalis nervi vagi. Вверху eminentia medialis имеет возвышение — лицевой бугорок, colliculus acialis, обусловленный прохождением корешка лицевого и проекцией ядра отводящего нервов. В области латеральных угтов располагается с обеих сторон вестибулярное поле, area vestibularis, здесь помещаются ядра VIII пары. Часть выходящих из них волокон идет поперек ромбовидной ямки от латеральных углов к срединной борозде в виде горизонтальных полосок, striae medullares ventriculi quarti. Эти полоски делят ромбовидную ямку на верхнюю и нижнюю половины и соответствуют границе между продолговатым мозгом и мостом.  Серое вещество спинного мозга непосредственно переходит в серое вещество мозгового ствола и частью расстилается по ромбовидной ямке и стенкам водопровода, а частью разбивается на отдельные ядра черепных нервов или ядра пучков проводящих путей. Чтобы понять расположение этих ядер, нужно учитывать, как отмечалось, что замкнутая нервная трубка при переходе от спинного в продолговатый мозг раскрылась на своей задней стороне и развернулась в ромбовидную ямку. Вследствие этого задние рога серого вещества спинного мозга как бы разошлись в стороны. Заложенные в задних рогах соматически-чувствительные ядра расположились в ромбовидной ямке латерально, а соответствующие передним рогам соматически-двигательные ядра остались лежать медиально. Что касается вегетативных ядер, заложенных в боковых рогах спинного мозга, то соответственно положению боковых рогов между задними и передними эти ядра при развертывании нервной трубки оказались лежащими в ромбовидной ямке между соматически-чувствительными и соматически-двигательными ядрами. В результате в области ромбовидной ямки в отличие от спинного мозга ядра серого вещества расположены не в перед-незаднем направлении, а лежат рядами — медиально и латерально. Так, например, соматически-двигательные ядра XII и VI пар лежат в медиальном ряду, вегетативные ядра X, IХ, VII пар — в среднем ряду и соматически-чувствительные ядра VIII пары — латерально. Практическое задание № 3 Тема 8. Строение и функции конечного мозга (большие полушария) Задание № 1.Напишите небольшое эссе на одну из тем по выбору Основные борозды и извилины долей коры полушарий. Верхнебоковая (конвекситальная) поверхность полушарий. Самая большая и глубокая – латеральная борозда (sulcus lateralis), или сильвиева борозда, - отделяет лобную и передние отделы теменной доли от расположенной ниже височной доли. Лобная и теменная доли разграничиваются центральной, или роландовой, бороздой(sulcus centralis), которая прорезает верхний край полушария и направляется по конвекситальной его поверхности вниз и вперед, немного не доходя до латеральной борозды. Теменную долю отделяют от расположенной позади нее затылочной доли проходящие по медиальной поверхности полушария теменно-затылочная и поперечная затылочная борозды. В лобной доле впереди центральной извилины и параллельно ей расположена предцентральная (gyrus precentralis), или передняя центральная, извилина, которую спереди ограничивает предцентральная борозда (sulcus precentralis). От предцентральной борозды кпереди отходят верхняя и нижняя лобные борозды, разделяющие конвекситальную поверхность передних отделов лобной доли на три лобные извилины - верхнюю, среднюю и нижнюю (gyri frontales superior, media et inferior). Передний отдел конвекситальной поверхности теменной доли составляет находящаяся позади центральной борозды постцентральная (gyrus postcentralis), или задняя центральная, извилина. Сзади ее окаймляет постцентральная борозда, от которой назад тянется внутритеменная борозда (sulcus intraparietalis), разграничивающая верхнюю и нижнюю теменные дольки (lobuli parietales superior et inferior). В нижней теменной дольке в свою очередь выделяются надкраевая извилина(gyrus supramarginalis), окружающая задний отдел латеральной (сильвиевой) борозды, и угловая извилина (girus angularis),окаймляющая заднюю часть верхней височной извилины. На конвекситальной поверхности затылочной доли мозга борозды неглубокие и могут существенно варьировать, вследствие этого вариабелен и характер расположенных между ними извилин. Конвекситальную поверхность височной доли разделяют имеющие почти параллельное с латеральной (сильвиевой) бороздой направление верхняя и нижняя височные борозды, разделяющие конвекситальную поверхность височной доли на верхнюю, среднюю и нижнюю височные извилины (gyri temporales superior, media et inferior). Верхняя височная извилина составляет нижнюю губу латеральной (сильвиевой) борозды. На ее поверхности, обращенной в сторону боковой борозды, имеется несколько поперечных мелких борозд, выделяющих на ней небольшие поперечные извилины (извилины Гешля), которые можно рассмотреть, лишь раздвинув края латеральной борозды. Передняя часть латеральной (сильвиевой) борозды представляет собой углубление с широким дном, образующим так называемый островок (insula), или островковую долю (lubus insularis). Прикрьгающий этот островок верхний край боковой борозды называется покрышкой (operculum). Внутренняя (медиальная) поверхность полушария. Центральная часть внутренней поверхности полушария тесно связана со структурами про- межуточного мозга, от которых ее отграничивают относящиеся к большому мозгу свод (fornix) и мозолистое тело (corpus callosum). Последнее снаружи окаймлено бороздой мозолистого тела(sulcus corporis callosi), начинающейся у передней его части - клюва (rostrum) и заканчивающейся у его утолщенного заднего конца (splenium). Здесь борозда мозолистого тела переходит в глубокую гиппокампову борозду (sulcus hippocampi), которая проникает глубоко в вещество полушария, вдавливая его в полость нижнего рога бокового желудочка, в результате чего и образуется так называемый аммониев рог. Несколько отступив от борозды мозолистого тела и гиппокамповой борозды, располагаются мозолисто-краевая, подтеменная и носовая борозды, являющиеся продолжением друг друга. Эти борозды отграничивают снаружи дугообразную часть медиальной поверхности полушария мозга, известную как лимбическая доля(lobus limbicus). В лимбической доле различают две извилины. Верхнюю часть лимбической доли составляет верхняя лимбическая (верхняя краевая), или опоясывающая, извилина (girus cinguli), нижняя часть образована нижней лимбической извилиной, или извилиной морского конька (girus hippocampi), или парагиппокампальной извилиной (girus parahyppocampalis), в передней части которой имеется крючок (uncus). Вокруг лимбической доли мозга расположены образования внутренней поверхности лобной, теменной, затылочной и височной долей. Большую часть внутренней поверхности лобной доли занимает медиальная сторона верхней лобной извилины. На границе между лобной и теменной долями большого полушария расположена парацентральная долька(lobulis paracentralis), которая является как бы продолжением передней и задней центральных извилин на медиальной поверхности полушария. На границе между теменной и затылочной долями хорошо видна теменно-затылочная борозда(sulcus parietooccipitalis). От нижней части ее назад отходит шпорная борозда (sulcus calcarinus). Между этими глубокими бороздами расположена извилина треугольной формы, известная под названием клин (cuneus). Впереди клина находится четырехугольная извилина, относящаяся к теменной доле мозга, - предклинье (precuneus). Нижняя поверхность полушария. Нижняя поверхность большого полушария состоит из образований лобной, височной и затылочной долей. Прилежащий к средней линии отдел лобной доли составляет прямая извилина (girus rectus).Снаружи ее отграничивает обонятельная борозда (sulcus olphactorius), к которой снизу прилежат образования обонятельного анализатора: обонятельная луковица и обонятельный тракт. Латеральнее его, вплоть до латеральной (сильвиевой) борозды, выходящей на нижнюю поверхность лобной доли, расположены мелкие глазничные извилины (gyri orbitalis). Латеральные отделы нижней поверхности полушария позади латеральной борозды занимает нижняя височная извилина. Медиальнее от нее находится латеральная височно-затылочная извилина (gyrus occipitotemporalis lateralis), или веретенообразная борозда. Передние отделы ее с внутренней стороны граничат с извилиной гиппокампа, а задние - с язычной (gyrus lingualis) или медиальной височно-затылочной извилиной (gyrus occipitotemporalis medialis). Последняя своим задним концом прилежит к шпорной борозде. Передние отделы веретенообразной и язычной извилин относятся к височной доле, а задние - к затылочной доле мозга. Практическое задание № 4. Тема 9. Автономная нервная система. Общее представление о сенсорных системах. 2.Подготовьте доклад на тему по выбору: Рефлекторная дуга вегетативной нервной системы, ее отличие от дуги соматического рефлекса. Клеточное тело воспринимающего нейрона как для анимальной, так и для вегетативной нервной системы помещается в спинномозговом узле, ganglion spinale, куда стекаются афферентные пути как от органов животной жизни, так и от органов растительной жизни и который, таким образом, является смешанным анимально-вегетативным узлом. Клеточное тело вставочного нейрона вегетативной нервной системы в отличие от анимальной нервной системы помещается в боковых рогах спинного мозга. При этом аксон вставочного анимального нейрона, исходящий из клеток заднего рога, заканчивается в пределах спинного мозга среди клеток его передних рогов. Что же касается вставочного нейрона вегетативной нервной системы, то он в спинном мозге не заканчивается, а выходит за его пределы, к нервным узлам, расположенным на периферии. 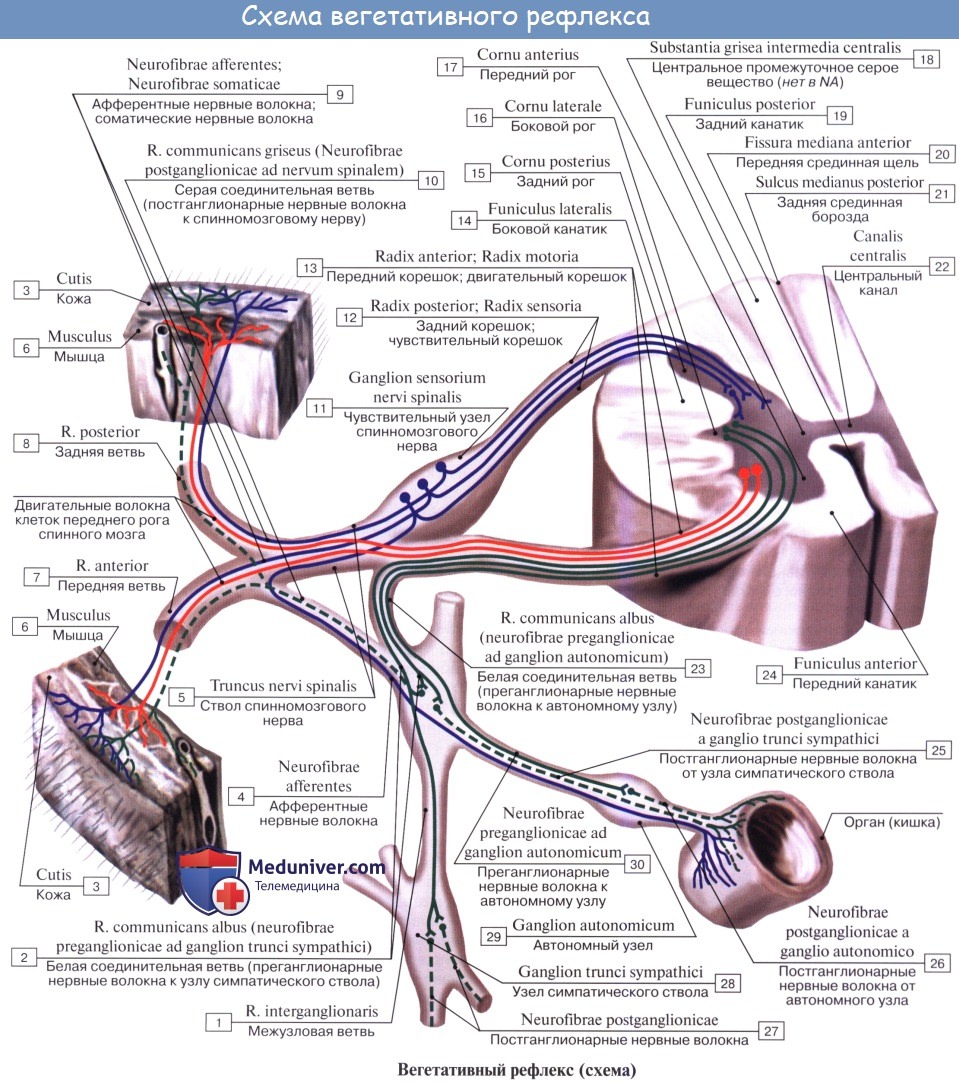 Выйдя из спинного мозга, аксон вставочного нейрона подходит или к узлам симпатического ствола, ganglia trunci sympathici, относящимся к симпатическому отделу вегетативной нервной системы (они образуют симпатический ствол), или волокна не заканчиваются в этих узлах, а направляются к предпозвоночным узлам, расположенным более периферично, между симпатическим стволом и органом (ganglia coeliaca, ganglia mesenterica). Эти узлы также относятся к симпатической системе. Наконец, волокна могут доходить, не прерываясь, до узлов, лежащих или около органа {околоорганные узлы, например ganglia ciliare, oticum и др.), или в толще органа (внутриорганные, интрамуральные узлы); и те и дугие называют конечными узлами (ganliga terminalia). Они относятся к парасимпатическому отделу вегетативной нервной системы. Кроме макроскопически видимых обособленных узлов, по ходу вегетативных нервов встречаются мигрировавшие сюда в ходе эмбрионального развития небольшие группы эффекторных нейронов — микроганглии. Все волокна, идущие до узлов первого, второго или третьего порядка и являющиеся аксонами промежуточного нейрона, называются предузловыми волокнами, rami preganglionares. Они покрыты миелином. Третий, эффекторный, нейрон анимальной рефлекторной дуги помещается в передних рогах спинного мозга, а эффекторный нейрон вегетативной рефлекторной дуги вынесен в процессе развития из центральной нервной системы в периферическую, ближе к рабочему органу, и располагается в вегетативных нервных узлах. Из такого расположения эффекторных нейронов на периферии вытекает главный признак вегетативной нервной системы — двухнейронность эфферентного периферического пути: первый нейрон — вставочный; тело его лежит в вегетативных ядрах черепных нервов или боковых рогах спинного мозга, а нейрит идет к узлу; второй — эфферентный, тело которого лежит в узле, а нейрит достигает рабочего органа. Эффекторные нейроны симпатических нервов начинаются в ganglia trunci sympathici (узлы первого порядка) или ganglia intermedia (узлы второго порядка), а для парасимпатических нервов — в около- или внутриорганных узлах, ganglia terminalia (третьего порядка); так как в названных узлах осуществляется связь вставочных и эфферентных нейронов, то отмеченная разница между симпатическим и парасимпатическим отделами вегетативной нервной системы связана именно с этими нейронами.  Аксоны эфферентных вегетативных нейронов почти лишены миелина — безмиелиновые (серые). Они составляют послеузловые волокна, rami postganglionics. Послеузловые волокна симпатической нервной системы, отходящие от узлов симпатического ствола, расходятся в двух направлениях. Одни волокна идут к внутренностям и составляют висцеральную часть симпатической системы. Другие волокна образуют rami communicantes grisei, соединяющие симпатический ствол с анимальными нервами. В составе последних волокна достигают соматических органов (аппарата движения и кожи), в которых иннервируют непроизвольную мускулатуру сосудов и кожи, а также железы. Совокупность описанных эфферентных вегетативных волокон, идущих от узлов симпатического ствола до органов сомы, составляет соматическую часть симпатического отдела. Такая структура обеспечивает функцию вегетативной нервной системы, которая регулирует обмен веществ всех частей организма применительно к непрерывно изменяющимся условиям среды и условиям функционирования (работы) тех или иных органов и тканей. Соответственно этой наиболее универсальной своей функции, связанной не с какими-либо отдельными органами и системами, а со всеми частями, со всеми органами и тканями организма, вегетативная нервная система и морфологически характеризуется универсальным, повсеместным распространением в организме. Следовательно, симпатический отдел иннервирует не только внутренности, но и сому, обеспечивая в ней обменные и трофические процессы. |