экзамен по биологии ПСПбГМУ. 1. Строение поверхностного аппарата клетки (пак)
 Скачать 0.66 Mb. Скачать 0.66 Mb.
|

|
14. Строение РНК. Транскрипция и процессинг и-РНК РНК – одноцепочечная полимерная молекула, образованная рибонуклеотидами, состоящими из рибозы, азотистого основания (пуриновые – аденин, гуанин, пиримидиновые – урацил, цитозин) и остатка фосфорной кислоты Различают и-РНК, несущие информацию о строении белка, р-РНК, входящие в состав рибосом, т-РНК, переносящие АК к месту синтеза и малые ядерные РНК, учавствующие в процессе сплайсинга и др. Транскрипция- синтез одноцепочечной РНК на матрице одноцепочечной ДНК Единица транскрипции, транскриптон, включает в себя: Инициатор Кодирующую область Терминатор Центральным ферментом, обеспечивающим процесс транскрипции, является ДНК-зависимая-РНКполимераза. У прокариот существует один универсальный фермент РНК-полимераза, которая работает на всех транскриптонах. У эукариот существует три варианта полимераз: полимераза I, полимераза II, полимераза III, которые взаимодействуют с тремя разными инициаторами и образовывать всевозможные виды РНК Полимераза I, реагируя с инициатором I отвечает за синтез большой рибосомальной РНК Полимераза II, реагируя с инициатором II отвечает за синтез и-РНК и малых ядерных Полимераза III, реагируя с инициатором III и IIIA отвечает за синтез т-РНК и малой рибосомальной РНК соответственно Для осуществления транскрипции полимераза связывается с белками-транскрибирующими факторами. Процесс энергозависим. Инициация начинается с района, который получил название инициатора. Он делится на два отдела: регулятор, промотор. Регулятор необходим для присоединения регуляторных молекул, которые могут активировать или тормозить процесс транскрипции. А к промотору в начале транскрипции присоединяется транскрибирующий фактор и только после этого комплекс промотор + Тф опознается РНК-полимеразой и связывается с ней. Промотор отвечает за начало транскрипции. Смысловая цепь – та, по которой идет синтез РНК, антисмысловая – ей комплементарная Образовавшийся белковый комплекс обладает геликазной активностью и способен к локальной деспирализации хромосом с разрывом водородных связей. Элонгация Идет формирование транскрипционного глазка. РНК-полимераза синтезирует 5’-3’ нить РНК по матрице ДНК, в транскрипционном глазке формируется двуцепочечная молекула из ДНК и РНК. Между дезоксирибонуклеотидами и рибонуклеатидами нет полного соответствия, они слабо реагируют друг с другом и не образуют прочных водородных связей, поэтому копия практически сразу же сходит с матрицы. По мере прохождения РНК-полимеразы, ДНК восстанавливает свою нормальную структуру в виде двойной спирали. Терминация Осуществляется с помощью двух видов терминаторов олигоадениловых и палиндромных, которые могут взаимодействовать с белковыми факторами терминации. ОТ содержат в своем составе несколько адениловых нуклеотидов, поэтому при образовании РНК на ней оказываются урациловые нуклеотиды. Водородные связи А=У слабые, поэтому происходит вытеснение 3’ конца синтезированной РНК из транскрипционного глазка. ПТ содержат в своем составе две взаимокомплементарные последовательности (ТТТААА), которые образуют шпильки за счет водородных связей между А и Т, что делает невозможным продвижение РНКполимеразы. ИНИ I имеют в своем составе две специфические последовательности нуклеотидов. С ними взаимодействует ряд регуляторных белков и транскрипционных факторов, которые способствуют присоединению РНК-полимеразы I к промотору и точному началу транскрипции с первого значащего кодона. ИНИ II содержат универсальный промоторный элемент – последовательность нуклеотидов, богатую Т и А – ТАТААА или ТАТА-бокс, находящийся на несмысловой цепи ДНК и определяющий точку начала ситнеза РНК. ИНИ III располагаются непосредственно в матрице транскрипции. Процессинг Процессинг рРНК - изменение первичных транскриптонов (происходит в ядре). В процессе транскрипции считывается 2 вида р-РНК: -большая р-РНК с коэффициентом седиментации 45S -малая р-РНК с коэффициентом седиментации 5S Малые р-РНК только подрезается по 3’ концу из-за неточности терминации. Большие р-РНК складываются с образованием двуцепочечных участков и так же подрезаются по 3’ концу, кроме того, она взаимодействует нековалентно с белковыми молекулами. Рибонуклеопротеид или РНП в таком виде разрезается на 3 части: 18S, 5,8S, 28S. Из этих фрагментов в ядрышковой сети, формируются большие субъединицы рибосом. Большая субъединица рибосом эукариот содержит 3 вида р-РНК: 5,8S, 5S, 28S и 49 белков. Малая субъединица рибосом эукариот содержит одну р-РНК 18S и 39 белков. В виде субъединиц р-РНК транспортируются из ядра в гиалоплазму. Процессинг тРНК В ходе транскрипции формируется первичная структура т-РНК в виде линейной нефункциональной молекулы. Процессинг начинается с того, что в транскриптоне образуется внутримолекулярные водородные связи. На вторичном уровне организации молекула т-РНК, подрезанная по 3’ концу принимает форму клеверного листа. В процессе созревания из плеча 2 вырезается кусок. Вторичный уровень организации нефункционален. Плечо 1 и плечо 3 необходимы для взаимодействия т-РНК с рибосомами. Плечо 2 необходимо для взаимодействия т-РНК с и-РНК, причем антикодон комплементарно способен взаимодействовать с кодонами и-РНК. У всех т-РНК на вторичном уровне организации унифицируется 3’ конец к нему нематрично присоединяется 3 нуклеотида ССА. После образования вторичной структуры за счет возникновения дополнительных водородных связей т-РНК переходит на третичный уровень организации и принимает вид так называемой L-формы. В таком виде т-РНК уходит в гиалоплазму. Процессинг иРНК Кэпирование – присоединение гуанозинтрифосфата к первому 5’-нуклеотиду. Это необходимо для: -правильного сплайсинга первого интрона в пре-и-РНК -для связывания и-РНК с рибосомой -обеспечения стабильности и-РНК -транспорта и-РНК в гиалоплазму Полиаденирование 3’-конца пре-и-РНК сопряжено с разрезанием специфической эндонуклеазной последовательности 5’-ААУААА-3’ и присоединяется от 100 до 300 остатков АМФ. Поли-А хвост необходим для: -эффективного удаления последнего интрона -транспорта и-РНК в гиалоплазму -унификации 3’-концов -для терминации и трансляции Сплайсинг иРНК Ему подвергаются не все виды РНК. В ходе процессинга иРНК связывается со специальными белками информатинами. Эти белки встречаются только в ядре, взаимодействуют друг с другом и образуют 15. Рибосомы. Трансляция. Строение белка Рибосомы - Небольшие частицы из двух субъединиц рибонуклеопротеидной природы. Трансляция: на матрице - и-РНК, синтезируется копия - полипептидная цепь (перевод нуклеотидной последовательности в последовательность аминокислот полипептидной цепи). Процесс трансляции возможен благодаря тому, что существует правила записи информации в и-РНК и правила ее реализации. Эти правила получили название генетический код. Свойства генетического кода: Триплетность (одной аминокислоте в полипептидной цепи соответствует 3 нуклеотида в иРНК) Непрерывность (между триплетами нет знаков препинания, те информация считывается непрерывно) Неперекрываемость (каждый нуклеотид входит в состав только одного кодона) Вырожденность или избыточность (1 АК может соответствовать нескольким кодонам и-РНК. Из 64 возможных кодонов 3 являются стоп-кодонами, эти кодоны не несут информации об аминокислотах и необходимы для окончания биосинтеза белка (УГА, УАГ, УАА кодоны детского плача). Некоторые аминокислоты могут кодироваться 6 кодонами, однако существуют аминокислоты, которые кодируются только 1 кодоном, например, аминокислота метионин и триптофан. Специфичность или однозначность (каждый кодон кодирует только одну аминокислоту) Универсальность (генетический код универсален для всех живых организмов) Коллинеарность (триплеты ДНК расположены в линейном порядке, и их последовательность соответствует последовательности АК в белке) Для биосинтеза белка требуется белоксинтезирующий аппарат клетки (и-РНК, т-РНК, рибосомы, белковые факторы инициации (ИФ1-ИФ6), элонгации, терминации), энергия молекул ГТФ. Можно выделить 3 функциональных района и-РНК: 5’-нетранслируемая область Лидер (необходима для взаимодействия и-РНК с рибосомой, возможно, регуляция работы генов на уровне инициации трансляции) Кодирующая область или матрица (начинается инициирующим кодоном метионином (АУГ), заканчивается стоп-кодоном, содержит информацию о первичной структуре белка – матрица трансляции) 3’-нетранслируемая область Трейлер (терминация трансляции, транспорт и-РНК через ядерную пору в гиалоплазму) Инициация начинается с того, что происходит диссоциация рибосомы на малую и большую субъединицу. К малой субъединице рибосомы присоединяется стартовая метионин-аминоацил-тРНК, имеющая антикодом УАЦ и несущая остаток метионина. К этому комплексу присоединяется иРНК и большая субъединица. Антикодон УАЦ аа-т-РНК взаимодействует с инициирующим кодоном АУГ и-РНК. Идет формирование активных центров рибосомы, стартовая метионин-аат-РНК связана с иРНК и рибосомой в Р-центре. А-центр или аминоацильный центр. Здесь происходит узнавание антикодонов новых аа-т-РНК Р-центр или пептидильный центр. Здесь происходит удержание пептидил-т-РНК Т-центр или трансферазный центр. Здесь сближаются карбокси- и амино-группы АК и образуются пептидные связи между ними Е-центр или эжекторный. Здесь происходит выталкивание освободившихся тРНК. Элонгация Под действием элонгирущего фактора в А-центре ко второму кодону и-РНК присоединяется аминоацил-т-РНК с АК-2, если ее антикодон будет комплементарен к кодону, находящемся в и-РНК, то в Т-центре АК-1 и АК-2 сближаются и образуют пептидную связь. Т-РНК с растущим пептидом становится пептидил-т-РНК. Стартовая аа-т-РНК освобождается, рибосома меняет свою конфирмацию и протаскивает и-РНК на один триплет вперед (транслокация). Стартовая аа-т-РНК выталкивается из рибосомы в Е-центре, а в Р-центр поступает пептидил-т-РНК с АК-1 и АК-2, образовавшими дипептид. В А-центре 3 триплет, кодирующий АК-3. Элонгация циклична до тех пор, пока в А-центр не попадет стоп-кодон (в т-РНК не существует антикодонов к терминальным кодонам). Терминация Пауза в синтезе белка идет с участием белкового фактора терминации – рилизингфактора. Он связывается со стоп-кодоном, активируя пептидилтрансферазу, катализирующую отщипление пептида от последней т-РНК. К последней АК присоединяется молекула Н2О для восстановления карбоксигруппы (-СООН). Белок покидает рибосому, диссоциирующую на большую и малую субъединицу. Ошибки терминации - замена кодона, следовательно, замена АК - сдвиг рамки считывания На процесс трансляции влияют различные антибиотики, но надо иметь в виду, что они блокируют синтез белков на митохондриальных рибосомах, имеющих константу седиментации 70S, как и рибосомы бактериальных клеток. Белки – высокомолекулярные полимерные органические вещества, определяющие структуру и жизнедеятельность клетки и организма в целом. Мономер – АК. F ферментативная (например, амилаза, расщепляет углеводы), структурная (например, входят в состав мембран клетки), рецепторная (например, родопсин, способствует лучшему зрению), транспортная (например, гемоглобин, переносит кислород или диоксид углерода), защитная (например, иммуноглобулины, участвуют в образовании иммунитета), двигательная (например, актин, миозин, участвуют в сокращении мышечных волокон), энергетическая (при расщеплении 1 г белка выделяется 4,2 ккал энергии), гормональная (например, инсулин, превращает глюкозу в гликоген) 16. Строение и функции нуклеиновых кислот. ДНК и РНК, понятие о матричных процессах ДНК – правозакрученная спираль, состоящая из двух антипараллельных комплеменарных полинуклеотидных цепей, связанных водородными связями Мономеры – дезоксирибонуклеотиды, состоящие из азотистого основания (пуриного – А, Г или пиримидинового – Ц, Т), дезоксирибозы и остатка фосфорной кислоты Нуклеотиды связываются между собой при помощи сложноэфирных связей, РНК – одноцепочечная полимерная молекула, образованная рибонуклеотидами, состоящими из рибозы, азотистого основания (пуриновые – аденин, гуанин, пиримидиновые – урацил, цитозин) и остатка фосфорной кислоты Различают и-РНК, несущие информацию о строении белка, р-РНК, входящие в состав рибосом, т-РНК, переносящие АК к месту синтеза и малые ядерные РНК, учавствующие в процессе сплайсинга и др. F хранение, передача и реализация генетической информации Матричный процесс – процесс копирования основы или матрицы, при этом первичная структура матрицы однозначно определяет первичную структуру копии. Для всех МП необходимы ферменты, белковые факторы и энергия АТФ (репликация, транскрипция, репарация, рекомбинация – в ядре, трансляция – в ЦП) Аденозинмонофосфорная кислота (АМФ) входит в состав всех РНК. При присоединении еще двух молекул фосфорной кислоты (Н3РО4) АМФ превращается в аденозинтрифосфорную кислоту (АТФ) и становится источником энергии, необходимой для биологических процессов, идущих в клетке. Реакции, в результате которых энергия освобождается - экзотермические реакции, реакции, потребляющие эту энергию – эндотермические. Последние реакции очень разнообразны: биосинтез, мышечные сокращения и т.д. Аденозинтрифосфорная кислота (АТФ) состоит из азотистого основания - аденина, сахара - рибозы и трех остатков фосфорной кислоты. Молекула АТФ очень неустойчива и способна отщеплять одну или две молекулы фосфата с выделением большого количества энергии, расходуемой на обеспечение всех жизненных функций клетки (биосинтез, трансмембранный перенос, движение, образование электрического импульса и др.). Связи в молекуле АТФ называют макроэргическими. Отщепление концевого фосфата от молекулы АТФ сопровождается выделением 40 кДж энергии. Синтез АТФ происходит в митохондриях. 17. Общая характеристика клеточного цикла Клеточный цикл – время жизни клетки от одного деления до другого. М-фаза 10% времени цикла, интерфаза – 90%. Интерфаза - пресинтетический период (до нескольких суток) - синтетический период (3-5 часов) - постсинтетический (6-8 часов) Постоянно в клеточной цикле находятся стволовые клетки крови, кишки или семенных канальцев. R период – состояние покое, временный уход из клеточного цикла, включающий в себя дифференцировку, но при этом способность к делению сохраняется – гепатоциты, эпителий кровеносных сосудов, клетки раковых опухолей. Покоящиеся клетки устойчивы к радиации, химическому воздействию и недостатку питательных веществ. При уходе клеток на терминальную дифференцировку способность к делению теряется, характерна фаза старения, а клеточный цикл завершается гибелью – нейроны, кардиомиоциты, эритроциты, палочки и колбочки сетчатки глаза. Интерфаза Необходима работа циклинзависимых протеинкиназ, активных только в комплексе с белкамициклинами (Е,Д,А,В), которые постоянно синтезируются и разрушаются в начале анафазы. G1-период Рост, синтез белка и РНК, восстановление наборов органелл Происходит синтез циклина Д, активирующего циклинзависимые протеинкиназы (2/4/6). Идет фосфорилирование белков, что необходимо для перехода в S-период. Фосфорилирование белка RB приводит к разрушению его комплекса с транскрипционным фактором E2F. Последний из которых, активировавшись, включает гены, отвечающие за репликацию ДНК. Проходя через рестрикционную точку (Rточку), клетка становится безразличной к сигналам, активирующих или ингибирующих клеточный цикл. При этом происходит проверка ДНК на наличие повреждений. Клетка находится в пресинтетическом периоде, пока не исправятся повреждения ДНК ферментами репарации. Иначе мутации => опухоли. Например, мутации гена RB белка приводят к стимуляции клеточных делений, идет образование ретинобластомы. Также к опухолям приводят мутации гена р53, что блокируют связывание белка р53 с ДНК, клетка продолжает делиться без остановки, тк репликация не блокируется. Циклин Д разрушается и начинается синтез циклина А и Е. S-период Репликация ДНК, синтез белков-гистонов, удвоение клеточного центра Репарация ошибок, произошедших при репликации (каждая хромосома из 2х хроматид). G2-период Синтез тубулинов и MPF (митозстимулирующий фактор), дезорганизация центриолярных сателлитов, формирование гало в клеточном центре, дополнительная репарация ДНК МПФ включает в себя циклин В и протеинкиназу р34, обладающих киназной и регуляторной активностями. При появлении этого фактора происходит конденсация хромосом и фосфорилирование 3х субстратов: 1) белков А, В, С ламины 2) гистонов Н1 3) белков СОСА (разрушение интерфазных микрофибрилл и микротрубочек и построение новых) Наиболее важная причина клеточного деления – клеточный рост. G1 (Я/ЦП1) G2 (Я/ЦП=1) 18. Митоз и его биологическое значение Непрямое деление клетки с образованием фигур деления, при котором из одной диплоидной материнской клетки образуются две дочерние, идентичные материнской. Диплоидная клетка имеет двойной набор хромосом, в котором каждая хромосома имеет себе пару или гомолога. Гомологичные хромосомы имеют одинаковую форму и размер и несут гены, отвечающие за одни и те же признаки. ПРОФАЗА Происходит разборка поровых комплексов ядра, конденсация хроматина за счет фосфорилирования гистонов Н1, фосфорилирование ламины А и С, разборка ламины до мембранных пузырьков, в составе которых остается белок В. Формирование вокруг центромеры хромосом особых белковых комплексовкинетохоров, к которым далее будут приклепляться нити веретена деления. Каждая хромосома состоит из двух сестринских хроматид, имеющих хромомерный уровень организации и объединены когериновым блековым комплексом. Кинетохор – 2 3х слойных полудиска, состоящих из фибрилл ДНК, обеспечивающих связь с центромерой, белков механохимической системы – кинезина, динеина и транслокаторных белков, имеющих сродство к тубулину. Динеин и кинезин обладают АТФазной активностью. Кинетохора движется по поверхности микротрубочки за счет энергии гидролиза АТФ (динеин от + к – концу, кинезин – наоборот). Оба эти белка перемещают хромосомы к полюсам в анафазе. Происходит расхождение удвоенного клеточного центра и обозначение полюсов клетки, от центриолей КЦ начинают формироваться МТ, образующие астральные, кинетохорные и полюсные нити веретена деления. Рост МТ – достраивание тубулина к + концу, а – конец фиксируется в цитоплазме. Функционирование веретена деления поддерживается сборкой и разборкой МТ. МЕТАФАЗА: Кинетохорные МТ приводят хромосомы к экватору, где они образуют метафазную пластинку. Хромосомы удерживаются натяжением нитей веретена деления. Это обеспечивается процессом непрерывного присоединения тубулинов к +концу МК и отсоединением их же от – конца. Матафазные хромосомы имеют петельное строение и максимально спирализованы. Также они имеют свой матрикс из рибонуклеопротеидов. Хромосома из 2х сестринских хроматид. Расположение хромосом определяет форму хромосом: метацентрические – равноплечие, субметацентрические – неравноплечие, акроцентрические – одноплечие. АНАФАЗА Происходит синхронное расщипление центромеры и хромосом на сестринские хроматиды. Активатор процесса – увелич конц Са в цитоплазме, выброс которых происходит из мембранных пузырьков. Сестринские хроматиды расходятся к полюсам. Происходит перемещение хроматид и укорочение кинетохорных микротрубочек, связанное с отсоед тубулина, движение кинетохора как скользящего кольца, поддерживающего боковую связь с полимеризованными тубулинами, раздвижение полюсов клетки. ТЕЛОФАЗА И ЦИТОКИНЕЗ Переход к телофазе идет за счет дефосфорилирования белков: гистона Н1 и белков ламины А и С. Пузырьки с ламиной В связываются с теломерами отдельных хромосом, которые начинают деспирализироваться. Далее пузырьки сливаются между собой и восстанавливают ядерную оболочку. Возобновляется синтез ДНК, восстанавливается ядрышко, начинается цитокинез. В плоскости метафазной пластинки образуется борозда деления. Цитокинез у эукариот – перетяжка благодаря актиновым и миозиновым филаментам, у прокариот – перегородка за счет пузырьков КГ. Значение – правильное распределение цитоплазматических органелл. Овоплазматическая сегрегация – попадание в бластомеры разных участков цитоплазмы в процессе дробления. Значение: равномерное распределение генетической информации между дочерними клетками. Рост и развитие, бесполое размножение, регенерация тканей. Патологии В первую очередь связаны с патологиями веретена деления, либо с патологией кинетохорного комплекса. Они могут быть наследственными, но их можно индуцировать, например, алкоголем и растительным алкалоидом колхицином. Если веретено деления разрушено до анафазы, то в клетке будут находиться хромосомы с двойным набором генетической информации, так называемые политентные хромосомы. В норме такие хромосомы характерны для некоторых секреторных клеток (слюнные железы). Если веретено деления разрушается на стадии анафазы, то образуются клетки, содержащие двойной набор хромосом или так называемые полиплоидные клетки. У животных полиплоидия практически несовместима с жизнью. У растений полиплоидия встречается часто. 19. Апоптоз и его механизмы Типы клеточной гибели Апоптоз – каскад метаболических событий, приводящий к самоуничтожению клетки. Происходит конденсация хроматина, разрушение ядра и цитоплазмы на мембранные тельца, выпячивание клеточной мембраны, фрагментация клетки с образованием дикретных апоптозных тел, которые в дальнейшем фагоцитируются. Запрограммированная клеточная гибель. Имеет важную роль в: Эмбриональном развитии (например, регресс личиночных органов), Уравновешивании результатов митоза и физиологическом обновлении клеток, Уничтожении мутантных клеток. Ингибиторы апоптоза – продукт гена bcl-2, белки FLIP, FAP, SENT, блокирующие активность каспаз. Неспособность претерпевать апоптоз вызывает рак (фолликулярные лимфомы, опухоли молочной железы, яичников и простаты), вирусные инфекции (герпес, аденовирус) и аутоиммунные заболевания (системная красная волчанка, гломерулонефрит). Ускоренная гибель клеток – СПИД и нейродегенеративные заболевания (синдром Альцгеймера и хорея Гентингтона). Некроз – катастрофическое разрушение клеточной целостности в результате повреждающего воздействия. Ранее разрушение клеточной мембраны, сморщивание МХ, отсутствие дальнейшего фагоцитирования. 20. Мейоз и его биологическое значение – деление клеток, при котором происходит уменьшение числа хромосом, и из диплоидной материнской клетки образуются гаплоидные дочерние. Состоит из 2х делений: - редукционное (уменьшение числа х-м в 2 раза) - эквационное (сохранение числа х-м) Первое деление Профаза I 5 фаз: - Лептонема - Зигонема - Пахинема - Диплонема - Диакинез Лептонема – стадия тонких нитей. Хромосомы из 2х сестринских хроматид тесно сближены и визуально не различимы. В световой микроскоп видны хромомеры. Хромосомы приклеплены к ламине прикрепительными дисками. Хромосома связана по всей длине с осевым белковым тяжем из белковгистонов. Зигонема – стадия сопряженных нитей. Происходит конъюгация (синаптонемальный процесс) благодаря белковому комплексу. Некоторые участки ДНК в предмейотический период не реплицируются, Хромосомы гибридизуются за счет комплементарных азотистых оснований (обычно эти участки в области теломер), Области удерживают гомологичные хромосомы у ядерной оболочки, инициация конъюгации, Конъюгация начинается с концов хромосом и идет к центру по принципу застежки-молнии, Осевые белковые тяжи гомологов сближаются и образуют два боковых элемента, Формирование центрального белкового элемента, сшивающегося с боковыми филаментами Бивалент или тетрада – конъюгирующие хромосомы (гомолог из 2х сестринских хроматид) Гомологи - хромосомы одинаковой формы и размера, несущие гены, отвечающие за одни и те же признаки. В каждой паре гомологов один получен от отца, а другой — от матери. Пахинема – стадия толстых нитей. Спирализация и компактизация, кроссинговер (появление в синаптонемальной щели рекомбинационных узелков, состоящих из ферментов рекомбинации, они подтягивают к себе локальные фрагменты ДНК, и в них происходит обмен участками). Рекомб узелки наблюдаются в тех участках хромосом, где есть гены. Происходит амплификация ДНК – процесс многократного копирования генов р-РНК И белков гистонов. Диплонема – стадия двойных нитей. В микроскоп видно двухром х-мы. Разрушение синаптонемального комплекса, отталкивание гомологичных х-м друг от друга в области центромеры. Теломеры остаются на месте и наблюдаются перекрестные фигуры – хиазмы, в которых происходит кроссинговер. У ооцитов лягушки Xenopus laevis хромосомы принимают форму «ламповых щеток». Происходит локальная деспирализация, транскрипция, образование большого количества РНК. У мутантов с малым кол-вом транскрипционных узелков – уменьшение частоты кроссинговера и хиазм, нарушение расхождения гомологов. Диакинез – стадия обособленных двойных нитей. Прекращение синтеза РНК, еще большее отталкивание гомологов, удерживающихся только концами плеч. ПАЯ разбирается, хромосомы не фиксированы на ламине, построение ахроматинового веретена деления. Метафаза I Биваленты сталкиваются нитями веретена деления в плоскость экватора, к каждой х-ме присоединяется одна кинетохорная нить веретена деления от ближайшего полюса. Анафаза I Нити ВД укорачиваются, к полюсам отходят двухроматидные х-мы, отделившиеся от гомолога. Телофаза I Деспирализация х-м. Восстановление ядерной оболочки и ядрышка, цитокинез. Образование 2х дочерних клеток с уменьшенным в 2 раза кол-вом х-м. Второе деление В интерфазу не происходит репликации, каждая х-ма из 2х сестринских х-д. Протекает как митоз. Дочерние клетки с гаплоидным набором х-м. Значение: поддержание постоянного числа х-м у особей одного вида, благодаря чему образовывается зигота, генотипическая изменчивость (кроссинговер, независимое расхождение). Генетический контроль мейоза Существуют гены, вкл и выкл мейоз, контролирующие конъюгация и кроссинговер, отвечающие за образование веретена деления. При мутациях у человека могут образовываться несбалансированные гаметы (бесплодие и невынашиваемость). Нарушение расхождения – трисомия по 21ой паре (с. Дауна), по 13ой (с. Патау), по по 18ой (с. Эдвардса). 21. Строение хромосом. Метафазные хромосомы. Кариотип человека Хромосомы состоят из ДНК и белков гистонов. Центромера из гетерохроматина соединяет хроматиды и делит х-му на плечи, на концах имеются теломерные участки. Организация хроматина: Нуклеосомный уровень - ДНК накручена на белковые коровые частицы, которые образованы 8 молекулами разных гистонов (Н2А, Н2В, Н3, Н4х2); Нуклеомерный уровень – сближение нуклеосом; Хромомерный уровень - укладка петлями нуклеомерного хроматина. Метафазные хромосомы имеют петельное строение и максимально спирализованы, для того чтобы предотвратить спутывание и разрыв ДНК при дальнейшем делении (каждая х-ма из 2х сестринских х-д). Метафазные х-мы имеют свой матрикс из рибонуклеопротеидов 21 Кариотип человека – диплоидный набор х-м, характеризующийся числом, размером и формой, которая определяется расположением центромер (метацентрические, субмета-, акро-). В норме кариотип: 46, ХХ – Ж, 46, ХУ – М. При нарушении нормального кариотипа возникают различные пороки развития. 22. Структура генов у прокариот и эукариот. Понятие о геноме Ген – участок молекулы ДНК (у некоторых вирусов РНК), несущий информацию об элементарной структуре или функции клетки. Геном – совокупность генов, локализованных в гаплоидном наборе хромосом клеток особей определенного вида и необходимых для нормального развития организма. Геном эукариот включает: Облигатные элементы структурные, несущие информацию о структуре и-РНК, и регуляторные гены, обеспечивающие работу структурных генов в процессе синтеза и-РНК. К регуляторным генам относят инициаторы, модуляторы, и терминаторы транскрипции. Инициатор находится перед структурным геном и содержит в себе промотор, узнающийся РНК-полимеразой. К модуляторам относят энхансеры и сайленсеры транскрипции, занимающие любое положение по отношению к структурным генам. Терминаторы, находящиеся после структурных генов, взаимодействуют с регуляторными белками терминации и заканчивают транскрипцию Структурные гены делятся на две группы: СГ-I - транскрибируемые, переводящие информацию с ДНК на разные РНК, СГ-II - транслируемые, переводящие информацию с ДНК на белок o Гены «домашнего хозяйства» - экспрессируются во всех клетках организма и необходимы для постоянного обеспечения жизнедеятельности (гены белков гистонов, тубулинов, гены т- и р-РНК) o Гены «роскоши» - экспрессируются только в дифференцированных клетках и необходимы для работы специализированных клеток и тканей на определенных этапах онтогенеза (гены белковых цепей гемоглобина (НВА, НВВ, НВD, HBE) экспрессируются в эритробластах, из которых формируются зрелые эритроциты. Также отвечают за адаптивный ответ клетки, например, при поступлении ионов тяжелых металлов активируется работа одного из двух генов, кодирующих белок металлотионеин, или при проникновении в организм различных ксенобиотиков – цитохром Р450) Факультативные элементы (некоторые повторяющиеся последовательности, вирусные ДНК, псевдогены, транспозоны, или мобильные генетические элементы) Оперон — способ организации генетического материала у прокариот, при котором цистроны (гены, единицы транскрипции), кодирующие совместно или последовательно работающие белки, объединяются под одним (или несколькими) промоторами. Такая функциональная организация позволяет эффективнее регулировать экспрессию (транскрипцию) этих генов. Отличия генома прокариот от эукариот: ДНК кольцевая, гены собраны в опероны, нет интронов, трансляция сопряжена с транскрипцией. 23. Характеристика генома человека 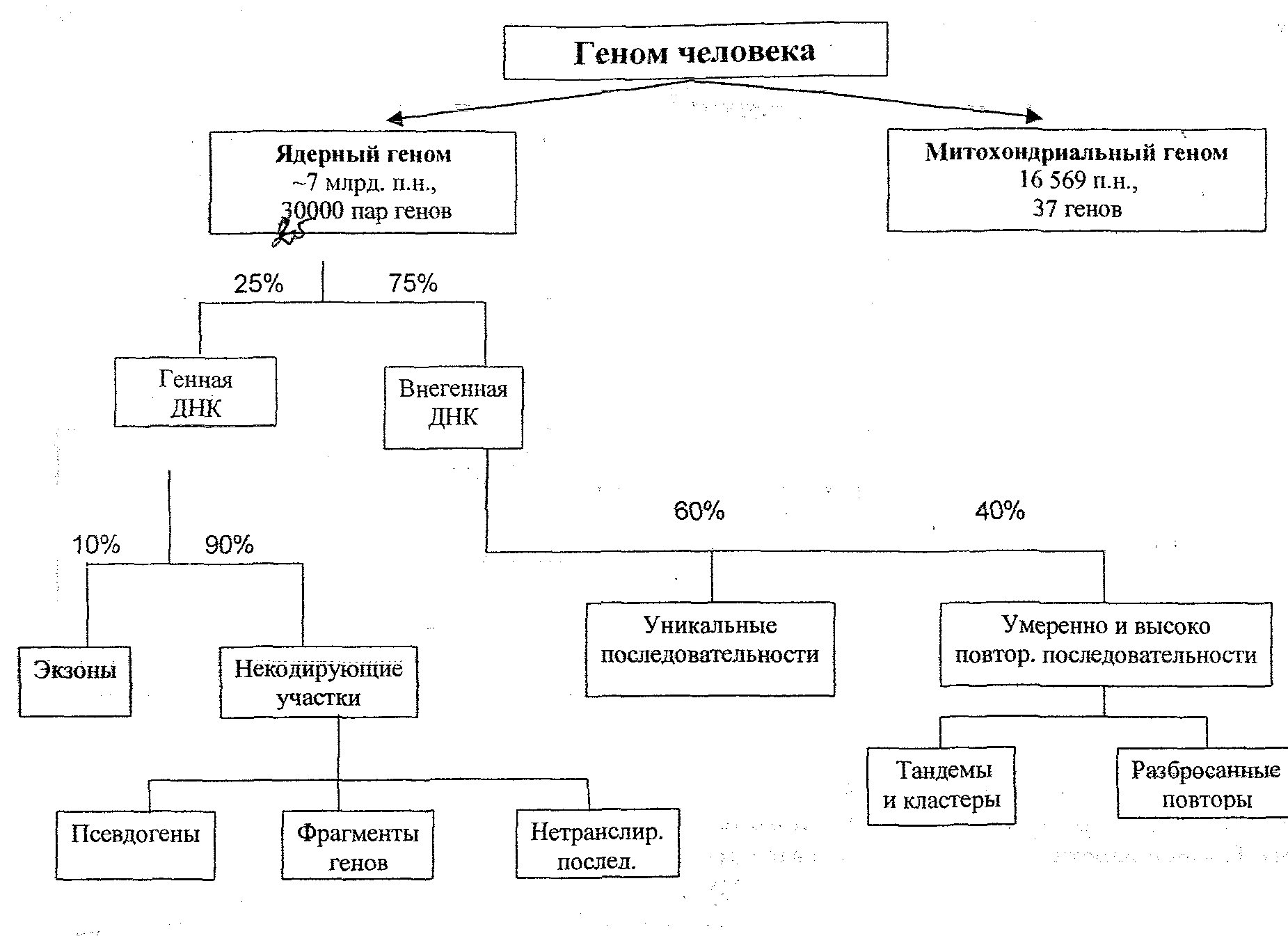 24. Строение и функция генов. Уровни реализации генетической информации Регуляцию действия генов можно осуществлять на разных уровнях: 1. Претранскрипционный – реализуется на уровне ДНК и хроматина 2. Транскрипционный – регуляция инициации транскрипции 3. Посттранскрипционный – осуществляется после синтеза РНК и базируется на различных вариантах процессинга РНК 4. Трансляционный – определяется возможностью изменять интенсивность синтеза полипептидов на стадии инициации трансляции 5. Пострансляционный – реализуется после синтеза полипептида и связан с процессингом молекул белка. Ген - функциональная единица генетического аппарата организма (ДНК или РНК); информационная структура, кодирующая полипептид, р-РНК или т-РНК. В состав гена входят азотистые основания (А, Г, Т, У); остатки фосфорной кислоты и сахар. Структура гена: мутоны – минимально одна пара нуклеотидов, способных мутировать (изменяться) реконы - минимально две пары нуклеотидов, способных рекомбинировать (меняться местами) цистроны – десятки и сотни нуклеотидов, контролирующих синтез полипептидов Свойства генов: 1) Свойство контролировать определенные ферментативные реакции. 2) Свойство к мутировать. 3) Свойство рекомбинировать. 4) Дискретность (делимость на более мелкие единицы). 5) Плейотропность (множественное действие). 6) Дозированность (зависимость результата от дозы гена). 7) Пенетрантность гена – количественный показатель: частота фенотипического проявления гена (в %) в популяции. 8) Экспрессивность – свойство характеризует степень выраженности признака. 9) Проявление гена зависит от факторов внешней среды. Строение оперона прокариот 1) Ген-промотор (место первичного прикрепления фермента) 2) Ген- оператор – включает и выключает работу структурных генов 3) Структурные гены расположены единым блоком – образуют центральное звено оперона 4) Ген – регулятор находится изолировано на расстоянии от основной части оперона. Он кодирует белок – репрессор, блокирующего при присоединении к гену – оператору транскрипцию – способность блокировать транскрипцию зависит от конформации белка – репрессора. Конформация меняется при 23 связывании его с веществами индукторами – субстратами ферментативных реакций, или супрессорами – продуктами ферментативных реакций, которые регулируют транскрипцию по типу прямой активации, индукции или обратной связи (ингибирование, репрессия). У эукариот оперон состоит из двух зон: информативной и неинформативной (акцепторной зоны). Информативная зона образована структурными генами. Особенности этой зоны у эукариот: а) один структурный ген может повторяться в опероне многократно б) структурные гены, ответственные за разные звенья одной цепи биохимических реакций могут находится не в одной, а в разных частях генома, т.е. рассеянными по всему геному. Неинформативная зона имеет 2 части: проксимальную (акцепторную) – представленную несколькими последовательно расположенными генами – промоторами, генами операторами; и дистальную (регуляционную) – представленную генами – регуляторами, рассеянными в различных местах генома. 25. Регуляция действия генов на претранскрипционном и транскрипционном уровнях Претранскрипционный (характерен для эукариот, тк кольцевая ДНК прокариот не связана с белками) Основан на изменении функциональной активности хроматина, зависящей от степени его конденсации. (см. уровни организации хроматина) - Химическая модификация белка-гистона Н1 (напр, фосфорилирование) → деспирализация хроматина и переход с нуклеомерного уровня на нуклеосомный → разборка нуклеосомы на две полунуклеосомы путем ацетилирования Н2А, Н2В, Н3, Н4 с помощью ацетилаз → ослабление связей между ДНК и гистонами - Помимо этого, возможна модификация самой ДНК Метилазы присоединяют метильные группировки к цитозину, расположенном на CG-островках в составе инициатора → снижение уровня синтеза РНК Метилирование ДНК поддерживается в ряду последовательных клеточных делений. (Репликация ДНК → две дочерние молекулы с одной метилированной цепью → неметилированные CG-сайты узнаются и метилируются поддерживающей метилазой). Инактивация генов (спираль левозакручена) сохраняется и в дальнейших делениях – «геномный импритинг». У млекопитающих CG последовательности подвергаются масштабному метилированию в процессе гаметогенеза. Часть генов метилируется в яйцеклетках, часть – в сперматозоидах. В зиготе – полный набор работающих генов. Это необходимо, тк далее, при делении, часть генов метилируется, потому что на определённых этапах развития требуются различные гены. В каждой клетке синтезируются разные белки, определяющие разные свойства, следовательно, и разные функции. Это метилирование приводит к дифференцировке клеток в многоклеточном организме. С мед точки зрения геномный импритинг реализуется в различном проявлении одного и того же дефекта, наследуемого от родителей разного пола. Пример – делеция небольшого участка длинного плеча 15 хромосомы: если она получена от отца – синдром Прадера-Вилли (низкий вес при рождении, мышечная гипотония, умственная отсталость, ожирение, гипогонадизм), от матери – синдрос Ангельмана (судороги, необычная походка, немотивированный смех, отсутствие речи). Транскрипционный Реализуется на этапе инициации транскрипции, обусловлен изменением эффективности присоединения РНК-полимеразы к промоторам и инициаторам генов специфических функций. С эволюционной точки зрения самый распространённый т.к. требует наименьшей затраты энергии, осуществляется за счет белковых факторов транскрипции, которые по своим функциям являются регуляторными белками, имеющими ДНК-связывающие домены, позволяющие им взаимодействовать с ДНКпоследовательностями инициаторов. 1. Белки активаторы. Запускают транскрипцию. 2. Белки репрессоры. Подавляют транскрипцию. Негативная регуляция – с помощью репрессоров Позитивная регуляция – с помощью активаторов Позитивная индукция - включение гена с помощью белка-активатора, который, присоединяясь к активатору, делает возможной транскрипцию. Нужный активатор синтезируется заново, или активируется путем изменения конформации (присоединение сигнальной молекулы) Позитивная репрессия – выключение гена за счет прекращения транскрипции, достигающееся путем инактивации белка-активатора с последующим отсоединением от инициатора. 26. Регуляция действия генов на посттранскрипционном уровне Альтернативная деградация РНК Стабильность зависит от кэпирования и поли-А хвоста, однако у и-РНК для белков-гистонов поли-А хвоста нет и стабильность обеспечивается наличием шпильки. У трансферрина на 3-конце нетранслируемый участок, образующий шпильки, к которым присоединяется белок-регулятор, увеличивающий стабильность трансферриновой и-РНК. При избытке железа белок отсоединяется и и-РНК разрушается РНКазами. Дефект – нестабильность РНК, недостаток железа, анемия. На 3-концах короткоживущих и-РНК (белки цитодифференцировки) есть многократно повторяющиеся последовательности нестабильности 5-АУУУА-3. Присоединение к ним белков-регуляторов повышает чувствительность и-РНК к эндонуклеазам. Дефект – делеция АУ-участка превращает протоонкогены в онкогены. Альтернативный сплайсинг 25 Из одной молекулы пре-и-РНК может образоваться несколько вариантов зрелой и-РНК за счет удаления не только интронов, но и разных комбинаций экзонов. Если сплайсируется лидерный участок, то разные клетки будут синтезировать один и тот же ген с разной скоростью. ген CYP19, катализирующий превращение андрогенов в эстрогены При альтернативном сплайсинге и-РНК клеток яичников остается лидерный (овариальный экзон), а в остальных тканях он удаляется. Поэтому в них синтез эстрогена выше, чем в других эстрогенпродуцирующих (плацента, печень плода, кожа и жировая ткань). Дефект – половой инфантилизм при удалении овариального экзона в яичниках и ускоренное половое созревание и ложный гермафродитизм у девочек и мальчиков соответственно при сохранении данного экзона в других тканях. За счет данного сплайсинга возможно образование двух форм амилазы – расщепляющей крахмал в ротовой полости и гликоген в печени. Также активация В-лимфоцитов и их дифференцировка. Сохранение трансмембранного домена – образование клеточного рецептора, плазматического – антител. Альтернативное редактирование Изменение последовательности нуклеотидов РНК после транскрипции за счет удаления, присоединения или замены 1-2 нуклеотидов. Замена кодона ЦАГ на ЦЦГ, кодирующего проводимость кальция в натрий-кальциевом канале постсинаптической мембраны нейронов в и-РНК гиппокамта. Дефект – наследственная амнезия. Замена кодона ЦАА на стоп-кодон УАА в и-РНК гена АРОВ, кодирующего белок аполипопротеин В, участвующий в транспорте холестерола, ЖК и триглицеридов. В гепатоцитах не происходит редактирования, но за это отвечают энтероциты, укорачивающие количество аминокислотных остатков белка в 2 раза. Это обусловлено цитодифференцировкой. 27. Регуляция действия генов на трансляционном и посттрансляционном уровнях Трансляционный: На стадии инициации синтеза белка. При этом необходим белоксинтезирующий аппарат (зрелая и-РНК, аминоацил-т-РНК, большая и малая рибосомальная субъединица, факторы трансляции). Многие и-РНК 5-лидерными районами взаимодействуют со специальными белками, образуя информосомы => не соединяются с рибосомами и не участвуют в трансляции (например, оогенез, характеризующийся активностью многих генов). Их продукты – различные виды и-РНК, накапливающиеся в виде информосом. После оплодотворения происходит их разборка, активация иРНК, идет синтез белков, необходимых для репликации, дробления зиготы и образования бластомеров. Адаптивная регуляция при отсутствии железа в клетке лидерный участок и-РНК ферритина (связывание ионов железа) формирует шпильки, на которой находится тормозящий трансляцию регуляторный белок. При избыточном поступлении железа белок отсоединяется и происходит синтез ферритина. Дефицит данного белка – сидероз (отложение солей железа). Также адаптивная регуляция возможна путем модификации факторов регуляции трансляции (в эритробластах протеинкиназа инактивирует ИФ2 путем фосфорилирования при недостатке гема, неактивный ген препятствует синтезу глобиновых цепей. При появлении гема синтез глобиновых цепей возобновляется). Посттрансляционный 26 Альтернативный процессинг и химическая модификация пептидов. АП происходит с предшественником гормонов гипофиза – ПОМК, синтезирующийся в передней и средней доле гипофиза. В передней доле из него образуется липотропный и адренокортиковый гормоны, в средней – меланостимулирующий, кортикотропинподобный пептид, эндорфины и энкефалины. При поступлении в клетку этанола скорость деградации белка цитохрома Р450 уменьшается, он не разрушается, поступает в пероксисомы и участвует в окислении этанола. ХМ – фосфорилирование, дефосфорилирование, аллостерические взаимодействия с различными ионами, циклическими нуклеотидами (цАМФ, цГМФ), стероидными гормонами или другими белками 28. Медицинские аспекты регуляции действия генов. Глобиновые гены. Талассемия Синтез гемоглобина в онтогенезе человека: Всего в ходе онтогенеза синтезируется 3 типа гемоглобина. Гемоглобин – это сложная молекула, состоящая из небелковой части или гема и глобина в состав, которого входит 4 цепи: 2 α-подобных и 2 β-подобных. В эмбриогенезе синтезируется эмбриональный гемоглобин или гемоглобин G, который состоит из 2 ζ (зита) и 2 ε (эпсилон) цепей. Плодный период – Hb или гемоглобин F (2 α и 2 γ), после рождения гемоглобин А (2 α и 2 β) и гемоглобин А2 (2 α и 2 δ). Для каждого вида цепей существуют свои собственные гены, причем гены для α и β-подобных цепей они тесно сцеплены и локализованы в одной хромосоме. Смена синтеза гемоглобина необходима, поскольку плодный и эмбриональный гемоглобин обладают большим сродством к кислороду и, следовательно, способен отнимать его у матери. Гены для ζ и ε репрессируются, гены для α, β, γ репрессируются, ген для γ сначала активируется, потом репрессируется. Синтез гемоглобина осуществляется в предшественниках эритроцитов - эритробластах. Регуляция действия гена зависит от места, в котором созревает эритробласт и, следовательно, от позиционной информации. В эмбриогенезе эритробласты развиваются в желчном мешке. У плода эритробласты локализованы в селезенке и печени. После рождения – красный костный мозг. Разное окружение – это разные сигнальные молекулы. Возможны нарушения регуляции действия гена, которые выражаются в снижении уровня синтеза тех или иных цепей. В общем виде это приводит к снижению уровня гемоглобина, т.е. к анемии. Данный конкретный случай называется талассимией. Если нарушен синтез ζ и ε цепей гибнет эмбрион. Если не работает ген для α-подобных цепей, то наблюдается патология α0 – талассимия. Развитие эмбриона идет нормально, развитие плода затруднено, что приводит к мертворождению, либо к выкидышу. Возможен вариант α+ - талассимии, когда α – цепи синтезируются, но не в достаточном количестве. В этом случае может наблюдаться анемия у плода, которая будет продолжаться после рождения. Если не работает ген для -цепей, β0 – талассимия. Развитие эмбриона и плода нормально, но вскоре после рождения смерть. Β + -талассимия. Анемия при развитии плода, которая будет продолжатся после рождения. Известен случай β0 – талассимии, которая связана с 2 нарушениями регуляции действия гена, но при котором не будет наблюдаться анемии. При этом ген для -цепей не репрессируется. Наблюдается большое сродство к кислороду. Фетальный гемоглобин – в условиях высокогорья таким людям лучше. β-цепи не активируются, γ- не репрессируются 29. Репарация ДНК. Реактивационная, эксцизионная, пострепликативная и индуцируемая Репарация - система исправления ошибок, возникающих при репликации. Системы репарации: 1) Апуринизация - утрата пуринового основания (аденина или гуанина) , которая блокирует работу ДНК – полимераз 2) Окислительное дезаминирование и алкилирование азотистых оснований, приводящие к мутациям типа замены оснований 3) Возникновение тиминовых димеров при УФ облучении, сопровождающееся возникновением ковалентных связей между соседними тиминами и блокировкой работы ДНК - полимераз 4) Ковалентная сшивка двух цепей ДНК вследствие разрыва фосфодиэфирных связей при ионизирующем облучении Типы репарации: конститутивная, при которой ферменты репарации уже синтезированы в клетке -реактивационная (одноэтапное восстановление повреждений, происходящее в темноте или на свету. Темновая – апуриновые сайты восстанавливаются с помощью ДНК-инсертаз, присоединяя пуриновое азот основание на место удаленного. Алкилированные нуклеотиды узнаются ферментом алкилтрансферазой, присоединяющему метильную группу к собственному цистеину, после чего фермент инактивируется Световая – удаление тиминовых димеров, возникающих при действии УФ. Неферментативная или прямая фотореактивация – УФ лучи длиной 240 нм расщипляют ковалентные связи между тиминовыми основаниями. Ферментативная – фермент фотолиаза взаимодействует с тиминовыми димерами, разрывает связи, и структура ДНК восстанавливается. Дефект фотолиазы – пигментная ксеродерма, при которой наблюдается атрофия кожи, гиперпигментация и раковые опухоли) -эксцизионная (вырезание поврежденного участка ферментом эндонуклеазой. Дефект – синдром ЛуиБар, при котором происходит нарушение функций мозжечка и имунной системы) -репликативная (Дефект – синдром Блума, при котором наблюдается низкий рост, поражение кожи и возникновение опухолей лимфатических органов и жкт) -рекомбинационная -транскрипционная (При помощи белков транскрипционного комплекса. Дефект – синдром Коккейна, при котором наблюдается задержка роста, аномалии скелета и сетчатки, микроцефалия, разрушение миелиновой оболочки нервных волокон, умственная отсталость) индуцибельная, при которой повреждения индуцируют работу генов и происходит синтез ферментов репарации (при множественных повреждениях) |