1. Введение Жизнь на Земле представляет собой форму существования материи
 Скачать 28.03 Mb. Скачать 28.03 Mb.
|

15.2.1. Учение об ареалахАреал - это область географического распространения систематической единицы (вида, рода, семейства и т. д.). География растений имеет дело, прежде всего, с ареалами различных видов. Зная ареалы видов какого-либо рода, можно составить представление об ареале всего рода в целом. Так же обстоит дело и с ареалом семейства, который складывается из ареалов отдельных родов.Местонахождение - это конкретный географический пункт, где найдено или наблюдалось отдельное растение. Документом, подтверждающим нахождение в конкретном пункте определенного растения, служит, прежде всего, гербарный образец с указанием места его сбора. Пункты местонахождений отдельных растений, относящихся к изучаемому виду, могут быть нанесены на карту в виде точек. Карты с нанесенными на них точками - местонахождениями получили название точечных карт ареалов (рис. 15.14 ). Если соединить контурной линией все периферийные точки - местонахождения, то такую карту ареала называют точечно-контурной (рис. 15.15). Если распространение вида в пределах ареала не имеет существенного значения в окончательном варианте карты, территорию ареала обводят контурной линией и заштриховывают. Такая карта ареала называется контурно-штриховой ( рис. 15.16). 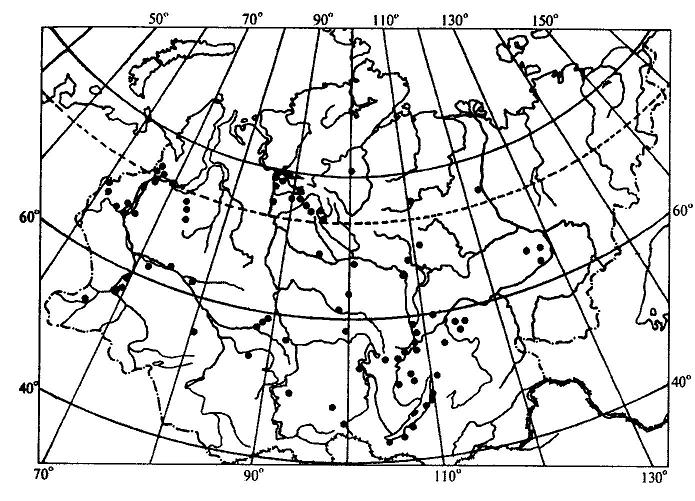 Рис. 15.14. Пример точечной карты ареала (ареал зубровки арктической (Hierochloe arctica)). Ни один вид не занимает всю территорию своего ареала сплошь. Это связано с тем, что даже в небольшом, относительно однородном географическом районе не наблюдается полной выравненности экологических условий. Как правило, ареалы растений охватывают значительные части суши со сложной топографией и множеством разнообразных местообитаний. Каждый вид нуждается по своей экологической природе в строго определенном местообитании, таким образом, неоднородность физико-географических условий на территории ареала неизбежно порождает прерывистое распределение особей и популяций вида, следовательно, можно говорить о топографической прерывистости ареала. Однако топографическая прерывистость ареала не нарушает его географической целостности. Ареал считается сплошным при непрерывном географическом распространении вида, хотя и при наличии прерывистости топографической. Растения водоемов, солончаков, болот, песчаных почв, приуроченные к специфическим местообитаниям, нередко обнаруживают особенно значительную прерывистость своего топографического распределения. Промежуточные пространства с иными местообитаниями заполняются популяциями других видов в соответствии с их экологической приспособленностью. В результате этого один и тот же географический район может входить в ареалы многих видов, т. е. ареалы разных растений обычно налагаются друг на друга в тех или иных частях. Например, в Западной Сибири произрастают такие древесные породы, как сибирский кедр, пихта, ель, сосна. Эти виды находятся здесь в пределах своего ареала, но в то же время каждый занимает свое местообитание: сосна на песчаных террасах рек, ель в поймах, на богатых наносных почвах, кедр и пихта на суглинках ближе к водоразделам. Ареалы этих видов налагаются только в определенной части равнины, тогда как размеры и общие очертания ареалов значительно расходятся. 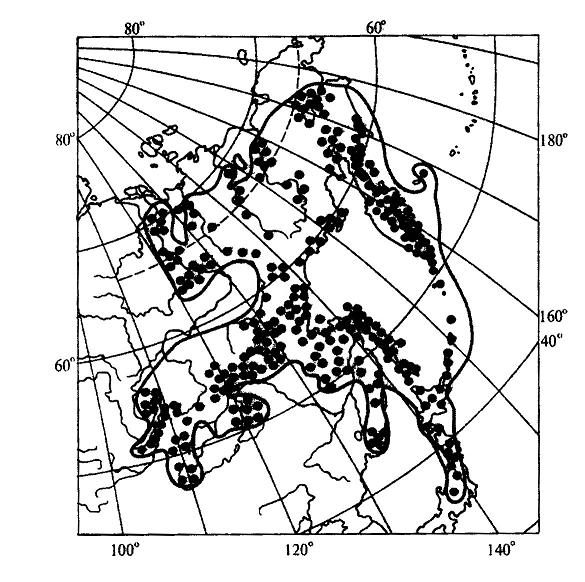 Рис. 15.15. Пример точечно-контурной карты ареала ( ареал кедрового стланика (Pinuspumila)). Рассматривая географическое распространение растений, мы можем иногда встретиться с таким фактом, когда промежуток между отдельными местонахождениями вида охватывает значительную часть суши, которая не может быть включена в единый ареал данного вида, так как здесь имеет место географическая прерывистость. В этом случае говорят о разрыве ареала, или его дизъюнкции. Дизъюнкция представляет собой довольно распространенное явление, особенно в родах и семействах. Таковы ареалы родов орех (Juglans), к которому относятся о. грецкий (J. regia), о. маньчжурский (J. mandshurica) и ряд американских видов; саговник (Cycas) и многих других. Разорванные (дизъюнктивные) ареалы обычно являются наследием геологического прошлого, т. е. представляют остатки некогда сплошных ареалов. Подтверждением этому являются ископаемые остатки растений, находимые на территориях современных разрывов ареалов. Например, в третичных отложениях северной Евразии, в том числе и Сибири, широко известны остатки орехов, родственных грецкому ореху, что свидетельствует о былом обширном ареале этого рода, имеющем в настоящее время дизъюнктивный характер. 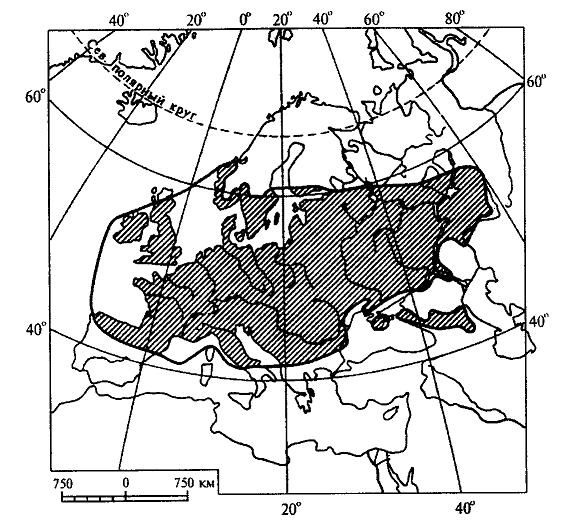 Рис. 15.16. Пример контурно-штриховой карты ареала (ареал дуба обыкновенного (Quercusrobur)). Ареал образуется в результате расселения вида и последовательного завоевания им определенного географического пространства и является результатом взаимодействия биологических особенностей вида с физико-географическими условиями в их историческом развитии. Следовательно, к основным факторам формирования ареалов можно отнести следующие: 1. Биологические особенности растений: а) естественные способы распространения; б) экологическая амплитуда. 2. Современные физико-географические условия, в которых происходит расселение вида. 3. Возраст вида. 4. Изменения физико-географических условий материков в геоисторическом масштабе. 5. Деятельность человека. В зависимости от тех причин, которые обусловливают границы ареалов, в географии растений различают следующие типы границ: климатические, непереходимые, почвенно-грунтовые, биотические, исторические. 1. Климатические границы возникают как следствие приспособленности растений к режиму тепла и влаги той природной зоны, где каждый данный вид существует в оптимальных условиях. У большинства растений Северного полушария, особенно древесных, полярная граница является климатической (ель, лиственница и др). Зависимость границ ареалов от климата проявляется в том, что во многих случаях эти границы идут по определенным параллелям или повторяют изолинии каких-либо климатических показателей. Наглядным примером являются северная и южная границы ареала семейства пальм, которые проходят по северному и южному тропикам. 2. Непереходимые границы возникают за счет физических преград, встающих на пути распространения растений. Сюда относятся обширные водные пространства - океаны, моря, высокие горные хребты, особенно в случае их широтного простирания. Эту же роль играют большие площади открытых ландшафтов пустынь, степей, тундр - для лесных растений и значительные лесные пространства - для растений степей и пустынь. 3. Почвенно-грунтовые, или эдафические, границы образуются при исчезновении необходимых местообитаний в связи с резким изменением химизма или физических свойств почв. Так, сильная засоленность северной половины Крымского полуострова является препятствием для расселения сюда многих степных видов, распространенных на юге Украины. Географически сплошная вечная мерзлота на северо-востоке Сибири является причиной образования здесь границ ареалов ряда древесных пород, обычных в остальной Сибири, таких как ель, сосна, сибирский кедр. 4. Биотические границы иногда называют еще конкурентными, так как связывают их с невозможностью внедрения вида в какую-либо местность потому, что необходимые для него местообитания уже раньше успели занять популяции другого вида со сходными экологическими требованиями. Именно это обстоятельство нередко ограничивает роль естественных факторов географического распространения растений. Виды, обосновавшиеся ранее в данной местности, “не пускают” пришельцев, не дают им размножиться и продвинуться за какую-то достигнутую ими границу. Биотическая граница может возникнуть также в том случае, если вид в процессе своего расселения достигает района, где по какой-либо причине отсутствует необходимое для него насекомое - опылитель. Не имея возможности обеспечить себя потомством, вид не перейдет через границу ареала этого насекомого. 5. Исторические границы, в отличие от предыдущих типов, обусловлены не современными причинами, а какими-нибудь событиями геологического прошлого, такими, как фазы горообразования, изменения конфигурации и связей материков, изменения климата. К историческим границам можно отнести также те случаи, когда вид в современную эпоху активно расселяется и не достиг еще потенциально возможных для него пределов. К такому типу можно отнести западную границу ареала сибирского кедра (рис. 15.17 ), который в своем расселении из Западной Сибири перевалил через Урал и Приуралье и еще не успел проникнуть дальше на запад. 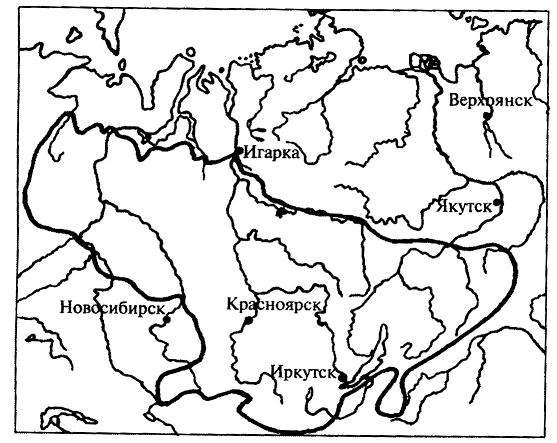 Рис. 15.17. Ареал кедра сибирского (Pinussibirica). Таким образом, у одного и того же вида разные границы его ареала могут иметь разное происхождение и относиться к различным типам. Так, северная, южная и отчасти восточная границы ареала сибирского кедра являются климатическими, северо-восточная - эдафической, а западная - исторической. Ареал в своем развитии проходит определенные стадии. Первоначально, в момент своего возникновения, вид часто имеет незначительный по площади ареал (первичный ареал). Затем, если условия среды благоприятны, происходит расселение вида, и ареал постепенно расширяется. В конце концов, вид достигает своих пределов распространения, ареал перестает существенно увеличиваться по площади, и его границы более или менее стабилизируются. Когда вид расселяется, его ареал находится в стадии прогресса. Однако ареалы могут претерпевать и обратные, регрессивные изменения, сокращаясь по площади. Это происходит вследствие изменения окружающей среды в неблагоприятном для вида направлении (например, похолодание или иссушение климата). В конечном счете, от некогда обширного ареала может остаться лишь очень небольшая его часть. Такие ареалы называют реликтовыми. Примерами растений, имеющих реликтовые ареалы, могут быть гинкго двулопастный (Ginkgobiloba), мамонтово дерево, или секвойядендрон гигантский (Sequoiadendrongiganteum), болотный кипарис (Taxodiumdistichum) и др. В зависимости от формы ареалы делят на две основные группы: 1) сплошные и 2) несплошные. Наиболее распространенными формами среди сплошных ареалов являются: 1. Опоясывающие - вытянутые вдоль всей суши земного шара по широте, это ареалы целых семейств или родов (семейства березовых, сосновых, пальм и др.). 2. Циркумполярные - охватывающие полярную окраину северной суши полукольцом или почти кольцом. 3. Овальные - вытянутые в меридиональном направлении или чаще по широте на ограниченном отрезке того или иного климатического пояса, наиболее распространенная форма ареалов видов и родов, но встречается и у семейств. 4. Лучистые и бахромчатые - обычно неправильной формы с неравномерными, часто многочисленными выступами в разных направлениях, это ареалы активно расселяющихся видов. К несплошным ареалам относятся: 1. Дизъюнктивные, или разорванные, - регрессивные ареалы, распадающиеся на два или несколько изолированных фрагмента (семейство саговниковых, роды орех, каштан и др.); дизъюнктивный ареал может возникнуть и вследствие случайного заноса семян, например перелетными птицами, в пригодные для произрастания вида места, находящиеся на значительном расстоянии от границ его основного ареала. 2. Продырявленные - встречаются у вымирающих папоротников и некоторых видов цветковых растений. 3. Точечные ареалы - слагаются из многочисленных, сильно разъединенных участков, характерны для водных, сорных и рудеральных (мусорных) растений. 4. Ленточные ареалы - представляют собой фрагменты, оторванные от основного, сплошного, ареала и разъединенные большими пространствами, непригодными для обитания данного вида; чаще всего это ареалы древесных видов и сопутствующих им трав и кустарников, проникающих в тундры, пустыни, саванны вдоль рек, около которых они и протягиваются узкими лентами. Размеры ареалов могут колебаться в широком диапазоне. Особенно значительные площади занимают опоясывающие ареалы некоторых семейств, некоторые роды (береза, сосна) также имеют очень обширные сплошные ареалы. Но ареал вида обычно ограничен какой-либо климатической зоной, охватывая лишь отдельную ее часть. Виды, встречающиеся на всех континентах, называются космополитами. К ним относятся некоторые водные растения, например элодея канадская (Elodeacanadensis), ряска малая (Lemnaminor), тростник обыкновенный (Phragmitesaustralis) и др. Обитатели водной среды никогда не испытывают недостатка во влаге и мало зависят от климата. Кроме того, эти растения способны быстро расселяться по территории, так как их семена, плоды, вегетативные органы легко разносятся водой или птицами. Обширные ареалы имеют некоторые сорные растения, произрастающие на нарушенных местообитаниях, где ослаблена конкуренция между растениями, и сопутствующие человеку на всех материках, например пастушья сумка (Capsellabursa-pastoris), крапива двудомная (Urticadioica) и ряд других. Отдельные местонахождения этих растений, как правило, настолько удалены друг от друга, что соединять их в один сплошной ареал было бы неправильно. Подобные ареалы обычно показывают на картах рассеянными точками, и ареал называют точечным. Если растение распространено в пределах примерно половины суши, то его относят к полукосмополитам. Последние наиболее часто встречаются среди споровых растений, например папоротник орляк (Pteridium aquilinum), некоторые мхи. Широко распространенные виды называют эврихорами, а виды, ограниченные в распространении, - стенохорами. Все космополиты - эврихоры, но не все эврихоры являются космополитами, так как некоторые из них встречаются повсеместно лишь на одном из континентов. В природе количественно преобладают мезохоры и стенохоры. К этим двум группам относится около 70% всех видов растений. Среди стенохоров имеются виды, ареал которых ограничен только каким-либо определенным регионом, и вне его они нигде больше не встречаются. Подобные растения называют эндемичными, или эндемиками (эндемами). Можно говорить, например, об эндемиках Кавказа, Крыма, Средней Азии, Алтая и т.д. Эндемичными могут быть не только виды, но и другие таксоны (роды, семейства и т.д.). Одни районы земного шара более богаты эндемиками, другие - менее, в третьих - эндемики отсутствуют. Особенно велика доля эндемичных растений во флоре древних морских островов. Так, во флоре Новой Зеландии насчитывается около 75% эндемичных видов. Древние острова особенно богаты эндемиками потому, что их растительный мир на протяжении очень длительного времени был полностью изолирован, и никакого обмена растениями с другими территориями не происходило. В этих условиях на островах в ходе эволюции возникли новые виды. Наряду с этим здесь могли сохраниться и некоторые растения, вымершие на остальной территории земного шара. Сравнительно велик процент эндемичных растений также в высоких поясах древних горных систем (субальпийском и альпийском). Причина этого явления - отсутствие обмена растениями с окружающими равнинными территориями. Высокогорья подобны островам в океане, флора их изолирована. В горных системах более молодого возраста эндемиков значительно меньше. Различают два типа эндемизма - палеоэндемизм (реликтовый эндемизм) и неоэндемизм. В первом случае речь идет о вымирающих таксонах, которые прежде были широко распространены, но сейчас встречаются лишь на очень ограниченных территориях. Например, гинкго двулопастный (Ginkgobiloba) - единственный современный представитель целого класса голосеменных растений, который в мезозойскую эру был широко распространен по всей Евразии и Северной Америке, - растет в диком состоянии на очень небольшой территории в горах Восточного Китая. Мамонтово дерево (Sequoiadendrongiganteum) встречается только в горах северной Калифорнии, тогда как в третичном периоде было распространено по всему Северному полушарию. Такие виды можно назвать реликтовыми эндемиками. В случае неоэндемизма речь идет, напротив, о молодых видах. Их ареал ограничен, главным образом, по той причине, что они еще не успели достаточно широко распространиться по земной поверхности. Примером неоэндемиков могут быть некоторые “молодые” виды первоцвета (Primula), колокольчика (Campanula), крупки (Draba), встречающиеся на Кавказе. Часто встречается явление викаризма. Викарирующими, или викарными (замещающими), называют виды и другие таксоны, которые мало различаются по морфологическим признакам и близко родственны между собой, но территориально разобщены. Эти виды как бы замещают друг друга на разной территории. В качестве примера можно назвать два вида ели, произрастающих в России, - ель европейскую (Piceaabies) и ель сибирскую (P. obovata). По морфологическим признакам они довольно похожи, но географическое распространение их различно. Ареал первого вида расположен в европейской части России, второго – включает северо-восток европейской части, Сибирь и Дальний Восток. Хорошим примером викаризма служат также виды лиственницы (Larix), которые в разных частях Северного полушария образовали почти непрерывный ряд форм, замещающих одна другую.
|