биоложди. 1) Ядерная оболочка (кариолемма)
 Скачать 1.23 Mb. Скачать 1.23 Mb.
|

|

| Облигатные элементы | Факультативные элементы |
| Структурные гены Гены тРНК Гены рРНК Обращенные повторы и палиндромы (регуляторные сайты) Сателлитная ДНК (центромерная, теломерная) | Мобильные генетические элементы (транспозоны) Псевдогены Вирусные и бактериальные последовательности Ретротранскрипты (кДНК) |
Облигатные последовательности представлены, в частности, характерными для данного вида структурными генами, число и расположение которых в геноме отличаются постоянством, и являются видоспецифичным. Облигатные элементы обеспечивают:
- формирование и передачу видоспецифических и индивидуальных, структурных, и функциональных признаков в ряду поколений;
- контроль онтогенетического развития.
Облигатные и факультативные элементы генома могут изменяться под влиянием факторов среды. Последствия изменений в факультативных элементах отличаются от таковых в структурных генах, т.к. последние приводят к изменению и фенотипа.
7) В настоящее время в геноме эукариот (в частности в геноме человека) выделяют три основных компонента или три основных типа ДНК-последовательностей.
1. Уникальные или редко повторяющиеся последовательности нуклеотидов в ДНК (нередко их называют - уникальные гены). Эти последовательности занимают большую часть генома, они составляют 60-70% ядерной ДНК. Встречаются такие последовательности один или несколько раз в геноме. Эти последовательности включают кодирующие участки и внутригенные некодирующие области. Кодирующие участки получили название экзоны, некодирующие участки этих последовательностей названы интронами. Экзоны уникальных генов разделены некодирующими участками – интронами (рис. 5).
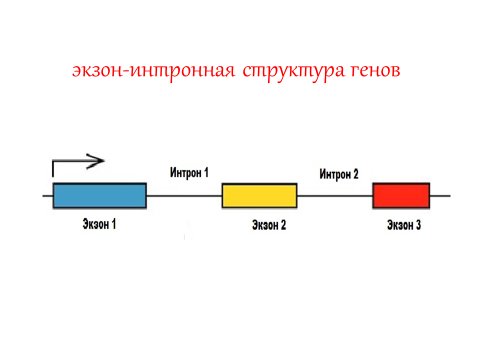
Рис. 5. Экзон-интронная структура гена
Экзоны – это кодирующие участки ДНК, которые будут транслироваться в аминокислотные последовательности белка. Количество экзонов в уникальных генах может быть различным. Встречаются гены, имеющие только один экзон. Максимальное число экзонов - 364, но в среднем в уникальных генах около 8 экзонов. В геноме эукариот на долю экзонов приходится 1,2-1,5% ДНК, на долю интронов около 24% ДНК. Однако лишь сумма последовательностей экзонов составляет последовательность гена, которая будет транслироваться в последовательность аминокислот в белке. Многие уникальные последовательности (уникальные гены) уже хорошо изучены. Они образуют основную массу структурных и регуляторных генов. Первым уникальным геном был открыт ген белка глобина, который входит в состав гемоглобина.
2. Умеренно повторяющиеся последовательности генома. Они составляют 10-15% ядерной ДНК. Такие последовательности повторяются сотни и тысячи раз в геноме. Эти последовательности включают гены рибосомных РНК и белков гистонов. Повторяющиеся единицы в этих последовательностях содержат определенный ген (или даже группу генов) и участки некодирующей ДНК, разделяющие гены - спейсеры. Так для белков гистонов в каждой повторяющейся единице 5 генов, которые соответствуют 5 видам гистонов:
| H1 | спейсер | H2A | спейсер | H2B | спейсер | H3 | спейсер | H4 |
3. Высокоповторяющиеся последовательности генома. Они составляет 25-28% ядерной ДНК. Эти последовательности повторяются сотни тысяч и даже миллионы раз в геноме, но каждая копия этих последовательностей относительно короткая (от нескольких пар до нескольких сотен пар нуклеотидов). Это некодирующие последовательности, которые включают ДНК структурного гетерохроматина (центромерные и теломерные районы хромосом).
В целом гены у эукариот занимают 30% всей ДНК, кодирующие области в геноме составляют порядка 3% всей ДНК. На долю кодирующих нуклеотидных последовательностей для белков в геноме приходится около 1,5% ДНК. Так организован ядерный геном эукариот (в том числе человека).
8) Избыточность генома эукариот Минусы "избыточной" ДНК: - увеличение времени синтеза ДНК; - cложнее организовывать удвоение ДНК; - высокая энергоемкость - на 1 нуклеотид для включения в цепь ДНК нужно затратить 60 молекул АТФ. Неопределенное следствие: -благодаря зависимости размера ядра от количества ДНК происходит увеличение размеров клетки. Плюсы "избыточной" ДНК: - возникает возможность создания сложного регуляторного аппарата, позволяющего поднять организм на более высокий эволюционный уровень. Причины избыточности: 1.Большой размер генов (за счет наличия интронов). 2. Присутствие повторенных последовательностей. Повторяются и гены, и некодирующие участки. 3. Наличие большого числа некодирующих последовательностей
9)

10)

11) Нуклеосомный. Нуклеосома — это структурная часть хромосомы, образованная совместной упаковкой нити ДНК с гистоновыми белками , , и . Они образуют напоминающие по форме шайбу белковые тела — коры, состоящие из восьми молекул (по две молекулы каждого вида гистонов) – гистонный октамер. Хроматин на этой стадии имеет вид «бусин», нанизанных на «нить» (ДНК), формирует нуклеофиламент (нуклеосомную нить). Участок ДНК между нуклеосомами называется линкерной ДНК, линкерные ДНК играют важную роль в дальнейшей упаковке хроматина, так как содержат нуклеотидные последовательности, специфически узнаваемые различными негистоновыми белками. Молекула ДНК комплектируется с белковыми корами, спирально накручиваясь на них. Благодаря нуклеосомной организации в основе структуры хроматина лежит нить, представляющая собой цепочку повторяющихся единиц — нуклеосом. В результате нуклеосомной организации хроматина диаметр двойной спирали ДНК в 2 нм увеличивается до 10 нм.Нуклеомерный. Дальнейшая компактизация нуклеосомной нити обеспечивается гистоном , который, соединяясь с линкерной ДНК и двумя соседними белковыми телами, сближает их друг с другом и сворачивает нуклеосомную фибриллу в спираль наподобие соленоида, формируя толстую нить сверхбусинок, толщина нити=30нм. Нуклеомер – глобула, образованная нуклеосомами. Хромомерный. Наиболее сложный. Спираль складывается, образуя петли различной длины, соединенные негистоновыми белками. Петли образуют розетковидные структуры. Диаметр такой структуры составляет 300 нм. Наиболее типична для интерфазной хромосомы. Хромонемный. 300 нм-фибриллы дополнительно сворачиваются, происходит сближение петель с образованием петлевых доменов. Диаметр такой структуры – 700 нм. Хроматида. Происходит спиральная укладка хромонемных нитей. Диаметр – 1400 нм.
12) Хроматин — это вещество хромосом — комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоидау прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК. Основную массу хроматина составляют белки гистоны. Гистоны являются компонентом нуклеосом, —надмолекулярных структур, участвующих в упаковке хромосом.
Классификация:
1.Эухроматин – локализующийся ближе к центру ядра, более светлый, более деспирилизованный, менее компактный, более активен в функциональном отношении. Эухроматин - неконденсированный хроматин, с которого происходит синтез белка.
2.Гетерохроматин - конденсированный хроматин, с которого белок не синтезируется. Гетерохроматин - плотно спирализованная часть хроматина, соответствует конденсированным, плотно скрученным сегментам хромосом, что делает их недоступными для транскрипции. Он интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид тёмных пятен, гранул.
Метафазные хромосомы состоят из двух продольных копий, которые называются сестринскими хроматидами и которые образуются при репликации. На стадии метафазы сестринские хроматиды соединены в районе первичной перетяжки, называемой центромерой. Центромера отвечает за расхождение сестринских хроматид в дочерние клетки при делении. На центромере происходит сборка кинетохора — сложной белковой структуры, определяющей прикрепление хромосомы к микротрубочкам веретена деления — движителям хромосомы в митозе. Центромера делит хромосомы на две части, называемые плечами. У большинства видов короткое плечо хромосомы обозначается буквой p, длинное плечо — буквой q. Длина хромосомы и положение центромеры являются основными морфологическими признаками метафазных хромосом.
В зависимости от расположения центромеры различают три типа строения хромосом:
1. Акроцентрические хромосомы, у которых центромера находится практически на конце, и второе плечо настолько мало, что его может быть не видно на цитологических препаратах;
2. Субметацентрические хромосомы с плечами неравной длины;
3. Метацентрические хромосомы, у которых центромерарасположена посередине или почти посередине.
13)
Метафазная хромосома состоит из двух хроматид. Любая хромосома имеет первичную перетяжку (центромеру) (5), которая делит хромосому на плечи.
Центромера (первичная перетяжка) - участок хромосомы, характеризующийся специфическими последовательностью нуклеотидов и структурой. Центромера принимает участие в соединении сестринских хроматид, формировании кинетохора, конъюгации гомологичных хромосом и вовлечена в контроль экспрессии генов.
Именно в области центромеры соединены сестринские хроматиды в профазе и метафазе митоза и гомологичные хромосомы в профазе и метафазе первого деления мейоза. На центромерах же происходит формирование кинетохоров: белки, связывающиеся с центромерой, формируют точку прикрепления для микротрубочек веретена деления в анафазе и телофазе митоза и мейоза.
Отклонения от нормального функционирования центромеры ведут к проблемам во взаимном расположении хромосом в делящемся ядре, и в результате - к нарушениям процесса сегрегации хромосом (распределения их между дочерними клетками). Эти нарушения приводят к анеуплоидии, которая может иметь тяжёлые последствия (например, синдром Дауна у человека, связанный с анеуплоидией (трисомией) по 21-й хромосоме).
Когда говорят о морфологии хромосом, то принимают во внимание следующие признаки: положение центромеры, длину плеч, наличие вторичной перетяжки и спутника.
В зависимости от положения центромеры в кариотипе человека выделяют хромосомы трех типов:
1. Метацентрические, равноплечие хромосомы: первичная перетяжка (центромера) расположена в центре (посередине) хромосомы, плечи хромосомы одинаковые.
2. Субметацентрические, почти равноплечие хромосомы: центромера находится недалеко от середины хромосомы, плечи хромосомы незначительно отличаются по длине.
3. Акроцентрические, очень неравноплечие хромосомы: центромера находится очень далеко от центра (середины) хромосомы, плечи хромосомы существенно различаются по длине.
Короткое плечо обозначают буквой –
Длинное плечо обозначают буквой -
Некоторые хромосомы имеют вторичную перетяжку (6) и спутник (сателлит) (7).
Вторичная перетяжка - участок хромосомы, соединяющий спутник с телом хромосомы. В области вторичной перетяжки расположены гены рибосомных РНК, происходит синтез рРНК и происходит формирование и сборка ядрышка. Такая вторичная перетяжка поэтому называется еще ядрышковым организатором.Вторичные перетяжки могут быть у одних хромосом на длинном плече, у других - на коротком.
От первичной вторичная перетяжка отличается отсутствием заметного угла между сегментами хромосомы.
У человека вторичную перетяжку имеют хромосомы9, 13, 14, 15, 21 и 22.
14) Кариотип человека
Понятие о кариотипе получило распространение на этапе исследования в медицине генетических заболеваний, когда стали активно изучать строение и функции хромосом. Получили открытие синдром Эдвардса, синдром Клайнфельтера. Кариотип, представляющий собой клеточный хромосомный комплекс, является постоянным. У человека нормой является наличие хромосом, количество которых равно 46. Из них 22 пары являются аутосомами и две — это половые хромосомы.
У представительниц женского пола они обозначаются как ХХ, у представителей мужского пола — ХУ. Главная особенность хромосомного набора — это видовая специфичность кариотипа. Функции хромосом заключаются в том, что каждая из них является носительницей генов, которые отвечают на наследственность.
Нормальный мужской кариотип — это кариотип 46, ХУ. Нормальный женский кариотип выглядит как кариотип 46, ХХ. Набор хромосом остается неизменным на протяжении всей жизни. Поэтому сдать кариотип достаточно один раз в жизни.
В характеристике кариотипа вначале указывается общее число хромосом и набор половых хромосом. Затем (при наличии мутаций) указываются геномные, а после – хромосомные мутации. Большое диагностическое значение для идентификации хромосом имеют хромосомные маркеры (chromosome landmarks) – полосы, наиболее четко выделяющиеся при дифференциальной окраске и разделяющие хромосому на районы. Нумеруются районы, а также полосы внутри района по направлению от центромеры к теломере. На некоторых полосах удается локализовать определенные гены генетической карты хромосомы (рис. 1).[2. 86-89]
Нормальный кариотип человека
В кариотипе человека 46хромосом или 23пары. Парные хромосомы называют гомологичными, они имеют одинаковою длину и форму, содержат аллельные гены. В состав хромосом входит 40 % ДНК, 40 % гистоновых белков и 20 % негистоновых белков. Комплекс всех химических веществ, входящих в состав хромосом, называется хроматином. Хромосомы могут находиться в клетках в двух структурных и функциональных состояниях – спирализованном и деспирализованном. В период интерфазы они находятся в деспирализованном состоянии. В спирализованном состоянии они находятся в период митоза. Максимальной спирализации хромосомы достигают в метафазе митоза. Метафазная хромосома состоит из двух хроматид, соединенных в области первичной перетяжки (центромеры). Некоторые хромосомы имеют вторичные перетяжки и спутники. Центромера делит хроматиду на два плеча.
15) 49.Дифференциальное окрашивание хромосом, его сущность и возможности.
Применяют 4 метода дифференциальной окраски: Q, G, R, C. Все они выявляют закономерную линейную неоднородность фрагментов по длине метафазных хромосом. Характер окрашивания специфичен для негомологичной хромосомы, что дает их точную идентификацию. Постоянство локализации окрашиваемых фрагментов позволяет составить «химические» карты хромосом. Сопоставление этих карт с генетическими используется для расшифрования функционально-генетических особенностей различных районов хромосом. 50.Что такое бенд при дифференциальной окраске хромосом. Определите положение представленного участка хромосомы: 1p31.
Бенд- участок хромосом, который отличается от соседнего по интенсивности окраски.
1p31 – первый бенд, локализованный в третьем районе короткого плеча первой хромосомы.
16)
В 1960 г. английский генетик Патау предложил классифицировать хромосомы человека на основании относительной длины и положения центромеры (центромерного индекса). Центромерный индекс - отношение длины короткого плеча к длине всей хромосомы. В соответствии с центромерным индексом различают метацентрические (перетяжка посередине), субметацентрические (одно плечо длиннее второго), и акроцентрические хромосомы (с непропорционально очень коротким одним плечом).
| | | | | | |
метацентрические субметацентрические акроцентрические
В 1960 г на международном генетическом симпозиуме в Денвере (США) была принята Международная (Денверская) классификация хромосом человека. Основные принципы классификации разработал Патау. В классификации учтены длина и форма хромосом. Все пары аутосом нумеруют арабскими цифрами от 1 до 22 в порядке уменьшения их длины. Половые хромосомы обозначают латинскими буквами Х и У и располагают в конце раскладки. У женщин в норме половые хромосомы ХХ, а у мужчин ХУ.
Все пары аутосом распределяются на 7 групп в соответствии с длиной и формой хромосом. Группы обозначают латинскими буквами от А до G. Группы четко отличаются друг от друга
Группа А (1,2,3 пары) самые длинные метацентрические (1,3) и субметацентрическая (2) хромосомы. Хромосома 1 - самая большая метацентрическая хромосома, центромера расположена посередине. Самой большой субметацентрической хромосомой является хромосома 2. Хромосома 3 почти на 20% короче хромосомы 1 и, следовательно, легко идентифицируется. Абсолютная длина от 11 мкм (1 пара) до 8,3 мкм (2 пара).
Группа В (4 и 5 пары) длинные субметацентрические хромосомы. Они не различаются между собой без дифференцированного окрашивания. Абсолютная длина 7,7 мкм.
Группа С (6 – 12 пары). Хромосомы среднего размера, субметацентрические. При стандартном (рутинном) окрашивании Х-хромосому нельзя отличить от других хромосом этой группы. Она по размерам сходна с хромосомами 6 и 7 пары. Абсолютная длина от 7,7 мкм (6 пара) до 5,8 мкм.
Группа D (13 - 15 пары). Эти акроцентрические хромосомы по форме сильно отличаются от всех других хромосом человека. Все три пары на коротком плече содержат спутники. Длина проксимальных участков коротких плеч варьирует, спутники могут отсутствовать, а могут быть очень большими, могут ярко флуоресцировать, а могут и не давать флуоресценции. Абсолютная длина от 4,2 мкм.
Группа Е (16 - 18 пары). Относительно короткие субметацентрические хромосомы. Абсолютная длина 3,6-3,5 мкм.
Группа F - (19 – 20 пары) маленькие метацентрические хромосомы. В препаратах при рутинной окраске они выглядят одинаково, но при дифференциальном окрашивании резко различаются. Абсолютная длина 2,9 мкм.
Группа G (21 – 22 пары) – две пары самых маленьких акроцентрических хромосом. На коротком плече имеют спутник. Изменчивость их коротких плеч так же значительна, как и в хромосомах группы D. Абсолютная длина 2,9 мкм.
Y-хромосома маленькая акроцентрическая хромосома длиной 2,8 мкм. Обычно (но не всегда) больше, чем хромосомы группы G, и хроматиды ее длинного плеча, как правило, лежат параллельно одна другой. Этим она отличается от хромосом группы G, у которых хроматиды длинных плеч образуют широкий угол.
Х хромосома сходна при рутинном окрашивании с хромосомами гриппы С, отличается при использовании дифференцированного окрашивания. Х - хромосома субметацентрическая, длиной 6,8 мкм.