Физиология сенсорных систем. Анализаторы. 12. физиология сенсорных систем (анализаторы)
 Скачать 2.54 Mb. Скачать 2.54 Mb.
|

|
Проводниковый отдел зрительного анализатора начинается зрительным нервом, который направляется из глазницы в полость черепа. В полости черепа зрительные нервы образуют частичный перекрёст, причём, нервные волокна, идущие от наружных (височных) половинок сетчатки, не перекрещиваются, оставаясь на своей стороне, а волокна, идущие от внутренних (носовых) половин её, перекрещиваясь, переходят на другую сторону (рис. 12.2). 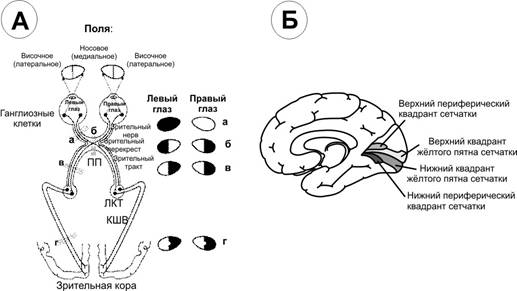 Рис. 12.2. Зрительные пути (А) и корковые центры (Б). А. Области перерезки зрительных путей обозначены строчными буквами, а возникающие после перерезки дефекты зрения показаны справа. ПП — перекрест зрительного нерва, ЛКТ — латеральное коленчатое тело, КШВ — коленчато–шпорные волокна. Б. Медиальная поверхность правого полушария с проекцией сетчатки в области шпорной борозды. После перекрёста зрительные нервы называются зрительными трактами. Они направляются к среднему мозгу (к верхним буграм четверохолмия) и промежуточному мозгу (латеральные коленчатые тела). Отростки клеток этих отделов мозга в составе центрального зрительного пути направляются в затылочную область коры головного мозга, где расположен центральный отдел зрительного анализатора. В связи с неполным перекрёстом волокон к правому полушарию приходят импульсы от правых половин сетчаток обоих глаз, а к левому – от левых половин сетчаток. Строение сетчатки.Самый наружный слой сетчатки образован пигментным эпителием. Пигмент этого слоя поглощает свет, вследствие чего зрительное восприятие становится более чётким, уменьшается отражение и рассеивание света. К пигментному слою прилежат фоторецепторные клетки. Из-за своей характерной формы они получили название палочек и колбочек. Фоторецепторные клетки на сетчатке расположены неравномерно. Глаз человека содержит 6-7 млн. колбочек и 110-125 млн. палочек. На сетчатке имеется участок размером 1,5 мм, который называют слепым пятном. Он совсем не содержит светочувствительных элементов и является местом выхода зрительного нерва. На 3-4 мм кнаружи от него находится желтое пятно, в центре которого расположено небольшое углубление – центральная ямка. В ней находятся только колбочки, а к периферии от неё число колбочек уменьшается и возрастает число палочек. На периферии сетчатки находятся только палочки. За фоторецепторным слоем расположен слой биполярных клеток (рис. 12.3), а за ним – слой ганглиозных клеток, которые контактируют с биполярными. Отростки ганглиозных клеток образуют зрительный нерв, содержащий около 1 млн. волокон. Один биполярный нейрон контактирует со многими фоторецепторами, а одна ганглиозная клетка – со многими биполярными. Рис. 12.3. Схема соединения рецепторных элементов сетчатки с сенсорными нейронами. 1 – фоторецепторные клетки; 2 – биполярные клетки; 3 – ганглиозная клетка. Отсюда, понятно, что импульсы от многих фоторецепторов сходятся к одной ганглиозной клетке, ибо число палочек и колбочек превышает 130 млн. Лишь в области центральной ямки каждая рецепторная клетка соединена с одной биполярной, а каждая биполярная – с одной ганглиозной, что создаёт наилучшее условия видения при попадании на неё световых лучей. Различие функций палочек и колбочек и механизм фоторецепции.Целый ряд факторов свидетельствует о то, что палочки являются аппаратом сумеречного зрения, т. е. функционируют в сумерках, а колбочки – аппаратом дневного зрения. Колбочки воспринимают лучи в условиях яркой освещённости. С их деятельностью связано восприятие цвета. О различиях в функциях палочек и колбочек свидетельствует структура сетчатки разных животных. Так, сетчатка дневных животных – голубей, ящериц и др. – содержит преимущественно колбочки, а ночных (например, летучих мышей) – палочки. Наиболее отчётливо воспринимается цвет при действии лучей на область центральной ямки, если же они попадают на периферию сетчатки, то возникает бесцветное изображение. При действии лучей света на наружном сегменте палочек зрительный пигмент родопсин разлагается на ретиналь – производное витамина А и белок опсин. На свету после отделения опсина происходит превращение ретиналя напосредственно в витамин А, который из наружных сегментов перемещается в клетки пигментного слоя. Считают, что витамин А увеличивает проницаемость клеточных мембран. В темноте происходит восстановление родопсина, для чего необходим витамин А. При его недостатке возникает нарушение видения в темноте, что называют куриной слепотой. В колбочках имеется светочувствительное вещество, сходное с родопсином, его называют йодопсином. Оно тоже состоит из ретиналя и белка опсина, но структура последнего неодинакова с белком родопсина. Вследствие целого ряда химических реакций, которые протекают в фоторецепторах, в отростках ганглиозных клеток сетчатки возникает распространяющееся возбуждение, направляющееся в зрительные центры головного мозга. Оптическая система глаза. На пути к светочувствительной оболочке глаза – сетчатке – лучи света проходят через несколько прозрачных поверхностей – переднюю и заднюю поверхности роговицы, хрусталика и стекловидного тела. Разная кривизна и показатели преломления этих поверхностей определяют преломление световых лучей внутри глаза (рис. 12.4). 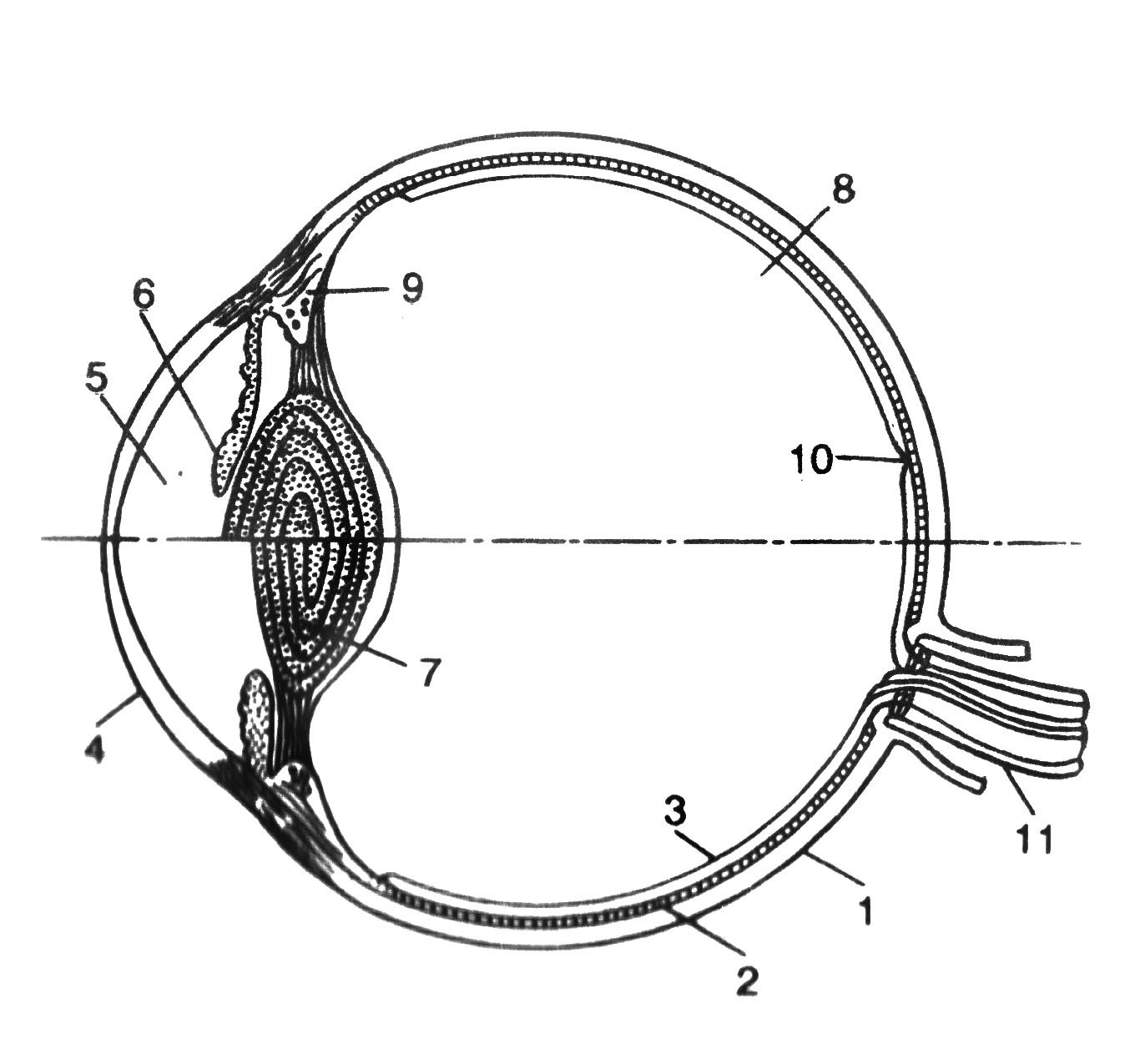 Рис. 12.4. Механизм аккомодации (по Гельмгольцу). 1 — склера; 2 — сосудистая оболочка; 3 — сетчатка; 4 — роговица; 5 — передняя камера; 6 — радужная оболочка; 7 — хрусталик; 8 — стекловидное тело; 9 — ресничная мышца, ресничные отростки и ресничный поясок (цинновы связки); 10 — центральная ямка; 11 — зрительный нерв. Преломляющую силу любой оптической системы выражают в диоптриях (D). Одна диоптрия равна преломляющей силе линзы с фокусным расстоянием 100 см. Преломляющая сила глаза человека составляет 59 D при рассматривании далёких и 70,5 D при рассматривании близких предметов. На сетчатке получается изображение, резко уменьшенное, перевёрнутое вверх ногами и справа налево (рис. 12.5). 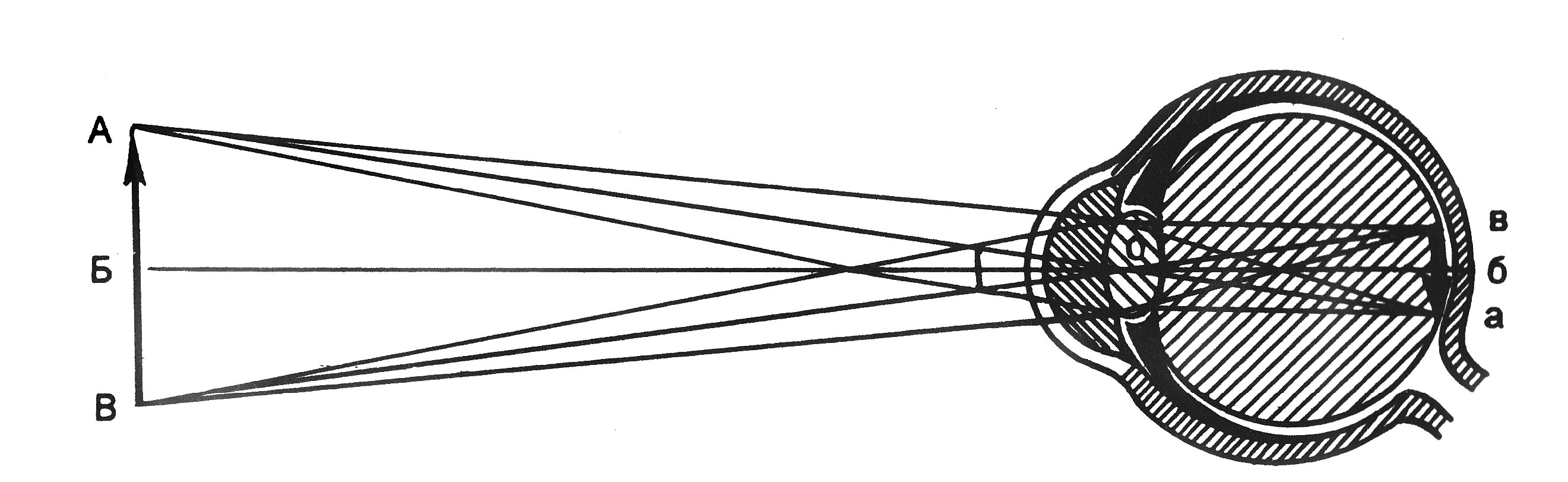 Рис. 12.5. Ход лучей от объекта и построение изображения на сетчатой оболочке глаза. АВ — предмет; ав — его избражение; 0 — узловая точка; Б — б — главная оптическая ось. Аккомодация. Аккомодацией называют приспособление глаза к ясному видению предметов, расположенных на разном расстоянии от человека. Для ясного видения объекта необходимо, чтобы он был сфокусирован на сетчатке, т. е. чтобы лучи от всех точек его поверхности проецировалась на поверхность сетчатки (рис. 12.6). 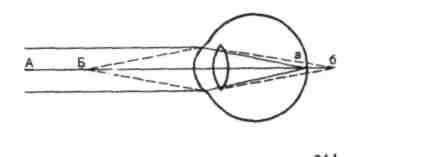 Рис. 12.6. Ход лучей от близкой и далекой точек. Объяснение в тексте Когда мы посмотрим на далёкие предметы (А), их изображение (а) сфокусировано на сетчатке и они видны ясно. Зато изображение (б) близких предметов (Б) при этом расплывчато, так как лучи от них собираются за сетчаткой. Главную роль в аккомодации играет хрусталик, изменяющий свою кривизну и, следовательно, преломляющую способность. При рассматривании близких предметов хрусталик делается более выпуклым (рис 12.4), благодаря чему лучи, расходящиеся от какой-либо точки объекта, сходятся на сетчатке. Аккомодация происходит благодаря сокращению ресничных мышц, которые изменяют выпуклость хрусталика. Хрусталик заключён в тонкую прозрачную капсулу, которую всегда растягивают, т. е. уплощают, волокна ресничного пояска (циннова связка). Сокращение гладких мышечных клеток ресничного тела уменьшает тягу цинновых связок, что увеличивает выпуклость хрусталика в силу его эластичности. Ресничные мышцы иннервируются парасимпатическими волокнами глазодвигательного нерва. Введение в глаз атропина вызывает нарушение передачи возбуждения к этой мышце, ограничивает аккомодацию глаза при рассматривании близких предметов. Наоборот, парасимпатомиметические вещества – пилокарпин и эзерин – вызывают сокращение этой мышцы. Наименьшее расстояние от предмета до глаза, на котором этот предмет ещё ясно видим, определяет положение ближней точки ясного видения, а наибольшее расстояние – дальней точки ясного видения. При расположении предмета в ближней точке аккомодация максимальна, в дальней – аккомодация отсутствует. Ближайшая точка ясного видения находится на расстоянии 10 см. Старческая дальнозоркость. Хрусталик с возрастом теряет эластичность, и при изменении натяжения цинновых связок его кривизна меняется мало. Поэтому ближайшая точка ясного видения находится теперь не на расстоянии 10 см от глаза, а отодвигается от него. Близкие предметы при этом видны плохо. Это состояние называется старческой дальнозоркостью. Пожилые люди вынуждены пользоваться очками с двояковыпуклыми линзами. Аномалии рефракции глаза. Преломляющие свойства нормального глаза называют рефракцией. Глаз без всяких нарушений рефракции соединяет параллельные лучи в фокусе на сетчатке. Если параллельно идущие лучи сходятся за сетчаткой, то тогда развивается дальнозоркость. В этом случае человек плохо видит близко расположенные предметы, а далеко расположенные – хорошо. Если же лучи сходятся перед сетчаткой, то тогда развивается близорукость, или миопия. При таком нарушении рефракции человек плохо видит далеко расположенные предметы, а близко расположенные – хорошо (рис. 12.7). 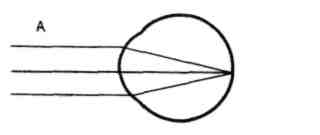 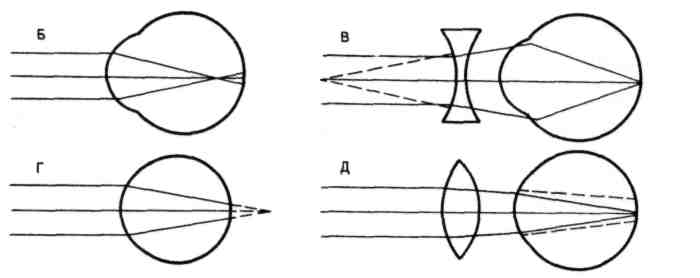 Рис. 12.7. Рефракция в нормальном (А), близоруким (Б) и дальнозорком (Г) глазу и оптическая коррекция близорукости (В) и дальнозоркости (Д) схема Причина близорукости и дальнозоркости заключена в нестандартной величине глазного яблока (при близорукости оно вытянутое, а при дальнозоркости оно приплюснутое короткое) и в необычной преломляющей силе. При близорукости необходимы очки с вогнутыми стёклами, которые рассеивают лучи; при дальнозоркости – с двояковыпуклыми, которые собирают лучи. К аномалиям рефракции относится также астигматизм, т. е. неодинаковое преломление лучей в разных направлениях (например, по горизонтальному и вертикальному меридиану). Этот недостаток в очень слабой степени присущ всякому глазу. Если посмотреть на рисунок 12.8, где одинаковые по толщине линии расположены горизонтально и вертикально, то одни из них кажутся более тонкими, другие – более толстыми. Рис. 12.8. Чертеж для выявления астигматизма Астигматизм обусловлен не строго сферической поверхностью роговой оболочки. При астигматизме сильных степеней эта поверхность может приближаться к цилиндрической, что исправляется цилиндрическими линзами, компенсирующими недостатки роговицы. Зрачок и зрачковый рефлекс. Зрачком называют отверстие в центре радужной оболочки, через которое лучи света проходят внутрь глаза. Зрачок способствует чёткости изображения на сетчатке, пропуская только центральные лучи и устраняя так называемую сферическую аберрацию. Сферическая аберрация состоит в том, что лучи, попавшие на периферические части хрусталика, преломляются сильнее центральных лучей. Поэтому, если не устранить периферических лучей, на сетчатке должны получиться круги светорассеяния. Мускулатура радужной оболочки способна изменять величину зрачка и тем самым регулировать поток света, поступающего в глаз. Изменение диаметра зрачка изменяет световой поток в 17 раз. Реакция зрачка на изменение освещённости носит адаптивный характер, так как несколько стабилизирует уровень освещённости сетчатки. Если прикрыть глаз от света, а затем открыть его, то расширившийся при затмении зрачок быстро суживается. Это сужение происходит рефлекторно («зрачковый рефлекс»). В радужной оболочке имеется два вида мышечных волокон, окружающих зрачок: кольцевые, иннервируемые парасимпатическими волокнами глазодвигательного нерва, другие – радиальные, иннервируемые симпатическими нервами. Сокращение первых вызывает сужение, сокращение вторых – расширение зрачка. Соответственно этому, ацетилхолин и эзерин вызывают сужение, а адреналин – расширение зрачка. Зрачки расширяются во время боли, при гипоксии, а также при эмоциях, усиливающих возбуждение симпатической системы (страх, ярость). Расширение зрачков – важный симптом ряда патологических состояний, например болевого шока, гипоксии. Поэтому расширение зрачков при глубоком наркозе указывает на наступающую гипоксию и является признаком опасного для жизни состояния. У здоровых людей размеры зрачков обоих глаз одинаковые. При освещении одного глаза зрачок другого тоже суживается; такая реакция называется содружественной. В некоторых патологических случаях размеры зрачков обоих глаз различны (анизокория). Это может происходить вследствие поражения симпатического нерва с одной стороны. Зрительная адаптация. При переходе от темноты к свету наступает временное ослепление, а затем чувствительность глаза постепенно снижается. Это приспособление зрительной сенсорной системы к условиям яркой освещённости называется световой адаптацией. Обратное явление (темновая адаптация) наблюдается при переходе из светлого помещения в почти неосвещённое. В первое время человек почти ничего не видит из-за пониженной возбудимости фоторецепторов и зрительных нейронов. Постепенно начинают выявляться контуры предметов, а затем различаются и их детали, так как чувствительность фоторецепторов и зрительных нейронов в темноте постепенно повышается. Повышение световой чувствительности во время пребывания в темноте происходит неравномерно: в первые 10 минут она увеличивается в десятки раз, а затем в течение часа – в десятки тысяч раз. Важную роль в этом процессе играет восстановление зрительных пигментов. Пигменты колбочек в темноте восстанавливаются быстрее родопсина палочек, поэтому в первые минуты пребывания в темноте адаптация обусловлена процессами в колбочках. Этот первый период адаптации не приводит к большим изменениям чувствительности глаза, так как абсолютная чувствительность колбочкового аппарата невелика. Следующий период адаптации обусловлен восстановлением родопсина палочек. Этот период завершается только к концу первого часа пребывания в темноте. Восстановление родопсина сопровождается резким (в 100000 – 200000 раз) повышением чувствительности палочек к свету. В связи с максимальной чувствительностью в темноте только палочек, слабо освещённый предмет виден лишь периферическим зрением. Теории цветоощущения. Существует ряд теорий цветоощущения; наибольшим признанием пользуется трёхкомпонентная теория. Она утверждает существование в сетчатке трёх разных типов цветовоспринимающих фоторецепторов – колбочек. О существовании трёхкомпонентного механизма восприятия цветов говорил ещё В.М. Ломоносов. В дальнейшем эта теория была сформулирована в 1801 г. Т. Юнгом, а затем развита Г. Гельмгольцем. Согласно этой теории, в колбочках находятся различные светочувствительные вещества. Одни колбочки содержат вещество, чувствительное к красному цвету, другие – к зелёному, третьи – к фиолетовому. Всякий цвет оказывает действие на все три цветоощущающих элемента, но в разной степени. Эта теория прямо подтверждена в опытах, где микроспектрофотометром измеряли поглощение излучений с разной длиной волны у одиночных колбочек сетчатки человека. Согласно другой теории, предложенной Э. Герингом, в колбочках есть вещества, чувствительные к бело-черному, красно-зелёному и желто-синему излучениям. В опытах, где микроэлектродом отводили импульсы ганглиозных клеток сетчатки животных при освещении монохроматическим светом, обнаружили, что разряды большинства нейронов (доминаторов) возникают при действии любого цвета. В других ганглиозных клетках (модуляторах) импульсы возникают при освещении только одним цветом. Выявлено 7 типов модуляторов, оптимально реагирующих на свет с разной длиной волны (от 400 до 600 нм). В сетчатке и зрительных центрах найдено много так называемых цветооппонентных нейронов. Действие на глаз излучений в какой-то части спектра их возбуждает, а в других частях спектра – тормозит. Считают, что такие нейроны наиболее эффективно кодируют информацию о цвете. Цветовая слепота. Частичная цветовая слепота была описана в конце XVIII в. Д. Дальтоном, который сам ею страдал (поэтому аномалию цветовосприятия назвали дальтонизмом). Дальтонизм встречается у 8% мужчин и намного реже у женщин: возникновение его связывают с отсутствием определённых генов в половой непарной у мужчин Х-хромосоме. Для диагностики дальтонизма, важной при профессиональном отборе, используют полихроматические таблицы. Люди, страдающие этим заболеванием, не могут быть полноценными водителями транспорта, так как они не могут различать цвет огней светофоров и дорожных знаков. Существует три разновидности частичной цветовой слепоты: протанопия, дейтеранопия и тританопия. Каждая из них характеризуется отсутствием восприятия одного из трех основных цветов. Люди, страдающие протанопией («краснослепые») не воспринимают красного цвета, сине-голубые лучи кажутся им бесцветными. Люди, страдающие дейтеранопией («зеленослепые») не отличают зелёные цвета от темно-красных и голубых. При тританопии – редко встречающейся аномалии цветового зрения, не воспринимаются лучи синего и фиолетового цвета. Все перечисленные виды частичной световой слепоты хорошо объясняются трехкомпонентной теорией цветоощущения. Каждый вид этой слепоты – результат отсутствия одного из трёх колбочковых цветовоспринимающих веществ. Встречается и полная цветовая слепота – ахромазия, при которой в результате поражения колбочкового аппарата сетчатки человек видит все предметы лишь в разных оттенках серого. Роль движения глаз для зрения. При рассматривании любых предметов глаза двигаются. Глазные движения осуществляют 6 мышц, прикреплённых к глазному яблоку. Движения двух глаз совершаются одновременно и содружественно. Рассматривая близкие предметы, необходимо сводить, а рассматривая далёкие предметы – разводить зрительные оси двух глаз. Важная роль движений глаз для зрения определяется также тем, что для непрерывного получения мозгом зрительной информации необходимо движение изображения на сетчатке. Импульсы в зрительном нерве возникают в момент включения и выключения светового изображения. При длящемся действии света на одни и те же фоторецепторы импульсация в волокнах зрительного нерва быстро прекращается и зрительное ощущение при неподвижных глазах и объектах исчезает через 1-2 с. Чтобы этого не случилось, глаз при рассматривании любого предмета производит не ощущаемые человеком непрерывные скачки. Вследствие каждого скачка изображение на сетчатке смещается с одних фоторецепторов на новые, вновь вызывая импульсацию ганглиозных клеток. Продолжительность каждого скачка равна сотым долям секунды, а амплитуда его не превышает 20º. Чем сложнее рассматриваемый объект, тем сложнее траектория движения глаз. Они как бы прослеживают контуры изображения, задерживаясь на наиболее информативных его участках (например, в лице – это глаза). Кроме того, глаз непрерывно мелко дрожит и дрейфует (медленно смещается с точки фиксации взора) – саккады. Эти движения также играют роль в дезадаптации зрительных нейронов. Типы движений глаз. Имеется 4 типа движений глаз.
Бинокулярное зрение. При взгляде на какой-либо предмет у человека с нормальным зрением не возникает ощущения двух предметов, хотя и имеется два изображения на двух сетчатках. Изображения всех предметов попадают на так называемые корреспондирующие, или соответственные, участки двух сетчаток и в восприятии человека эти два изображения сливаются в одно. Надавите слегка на один глаз сбоку: немедленно начнёт двоиться в глазах, потому что нарушилось соответствие сетчаток. Если же смотреть на близкий предмет, конвергируя глаза, то изображение какой-либо более отдалённой точки попадает на неидентичные (диспаратные) точки двух сетчаток (рис. 12.9). Диспарация играет большую роль в оценке расстояния, и, следовательно, в видении глубины рельефа. Человек способен заметить изменение глубины, создающее сдвиг изображения на сетчатках на несколько угловых секунд. Бинокулярное слитие или объединение сигналов от двух сетчаток в единый зрительный образ происходит в первичной зрительной коре. Зрение двумя глазами значительно облегчает восприятие пространства и глубины расположения предмета, способствует определению его формы и объёма. 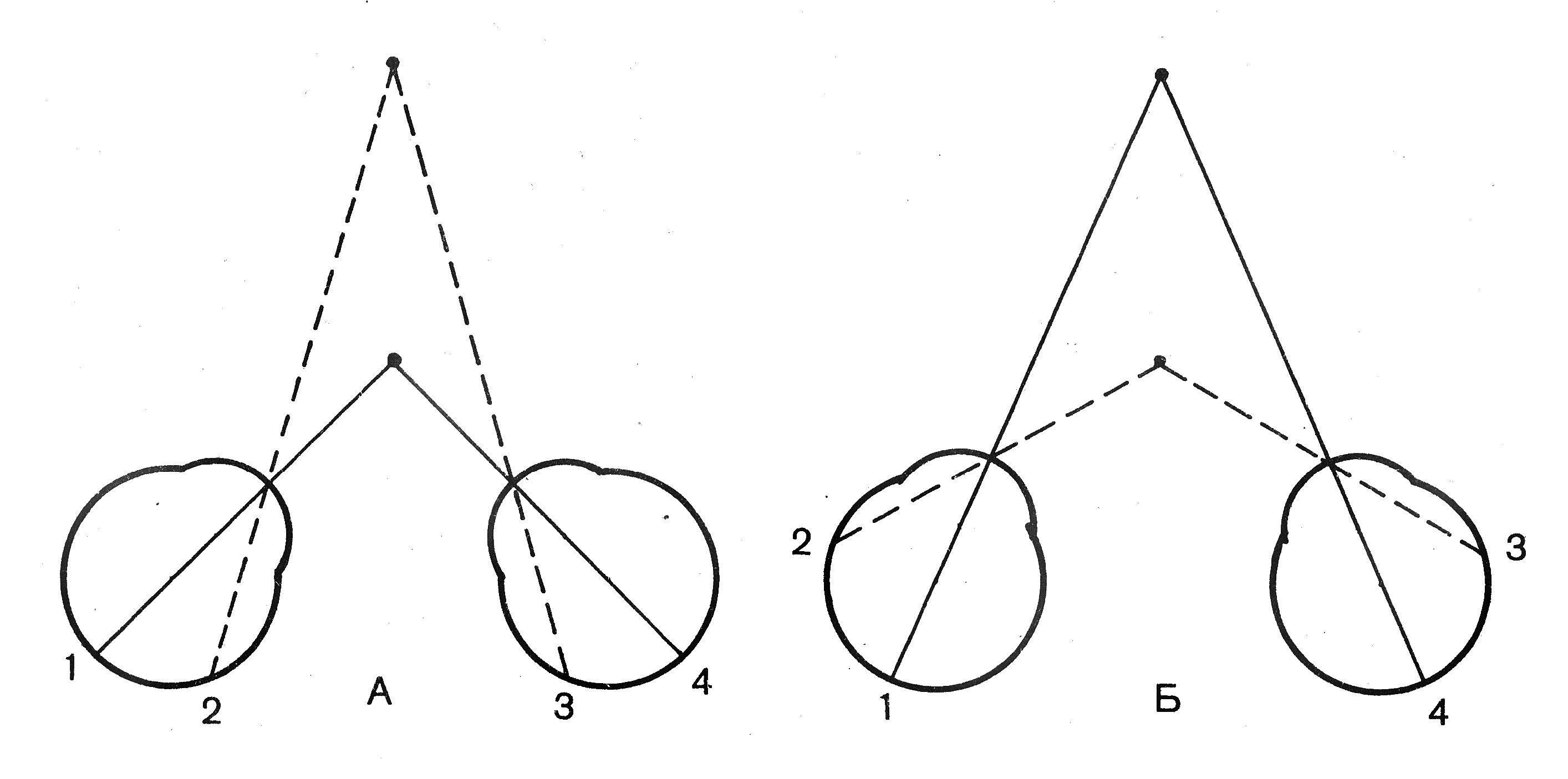 Рис. 12.9. Ход лучей при бинокулярном зрении. А – фиксирование взором ближайшего предмета; Б – фиксирование взором дальнего предмета; 1, 4 – идентичные точки сетчатки; 2, 3 – неидентичные (диспаратные) точки. Слуховой анализатор Слуховой анализатор (слуховая сенсорная система) – второй по значению дистантный анализатор человека. Слух играет важнейшую роль именно у человека в связи с возникновением членораздельной речи. Акустические (звуковые) сигналы представляют собой колебания воздуха с разной частотой и силой. Они возбуждают слуховые рецепторы, находящиеся в улитке внутреннего уха. Рецепторы активируют первые слуховые нейроны, после чего, сенсорная информация передаётся в слуховую область коры большого мозга (височный отдел) через ряд последовательных структур. Орган слуха (ухо) – это периферический отдел слухового анализатора, в котором расположены слуховые рецепторы. Строение и функции уха представлены в табл. 12.2, рис. 12.10. Таблица 12.2. Строение и функции уха
|